
Pandy Vijayapandi
Department of Pharmacology, Faculty of Medicine, University of Malaya, 50603 Kuala Lumpur, Malaysia
Subhash Harisankar
Department of Pharmacology, KMCH College of Pharmacy, Coimbatore-641 035, Tamil Nadu, India
Jose Nancy
Department of Pharmacology, KMCH College of Pharmacy, Coimbatore-641 035, Tamil Nadu, India
Journal of Pharmacology and Toxicology
Year: 2012 | Volume: 7 | Issue: 2 | Page No.: 87-95







ABSTRACT
The effect of acute treatment of telmisartan, an angiotensin AT1 receptor blocker at various doses (0.01, 0.05, 0.1, 1, 5, 10 and 20 mg kg-1, p.o.) on immobility in mice forced swim test has been studied. In addition, telmisartan in combination with fluoxetine hydrochloride, reserpine, haloperidol and reboxetine has also been studied on immobility. In the present study, telmisartan per se elicits significant bell shaped dose dependent depression- like effect in the forced swim test. In the drug-interaction studies, telmisartan (0.1, 1 and 5 mg kg-1, p.o.) significantly reversed the fluoxetine hydrochloride and reboxetine-induced immobility. Telmisartan, at same doses potentiated the reserpine and haloperidol-induced immobility in the forced swim test. The depression-like effect produced by telmisartan in the forced swim test in mice might be due to the unopposed AT2 receptor stimulation because of the complete blockade of AT1 receptor even at the low doses of telmisartan. The obtained results in the drug-interaction studies revealed the neuromodulatory effect of telmisartan on brain monoaminergic system.
PDF Abstract XML References Citation
Received: November 02, 2011;
Accepted: February 15, 2012;
Published: March 14, 2012
How to cite this article
Pandy Vijayapandi, Subhash Harisankar and Jose Nancy, 2012. Depression-like Effect of Telmisartan in Mice Forced Swim Test: Involvement of Brain Monoaminergic System. Journal of Pharmacology and Toxicology, 7: 87-95.
DOI: 10.3923/jpt.2012.87.95
URL: https://scialert.net/abstract/?doi=jpt.2012.87.95
DOI: 10.3923/jpt.2012.87.95
URL: https://scialert.net/abstract/?doi=jpt.2012.87.95
INTRODUCTION
Telmisartan is a potent, long-lasting, non-peptide antagonist of the angiotensin II type-1 (AT1) receptors that is indicated for the treatment of essential hypertension. It also provides a novel approach in addressing the multifactorial components of metabolic syndrome represents cluster metabolic and cardiovascular risk factors that predispose the sufferers to heart attack, stroke, heart failure and sudden cardiac death (Huq, 2007). AT1 receptor blockers (ARBs) are better alternative agents for heart failure in patients who are unable to tolerate ACE inhibitors (Balakumar and Singh, 2007). ARBs can also suppress the growth of melanoma tumors (Akhavan et al., 2011). Telmisartan is selectively and insurmountably inhibits stimulation of the AT1 receptor by angiotensin II (ang II). It has a high affinity to AT1 receptor with a Ki value of 3.7 nM which is higher than losartan and its active metabolite EXP 3174, whose values are 23.7 and 10.4 nM, respectively (Wienen et al., 2000). It has been reported telmisartan can penetrate the blood-brain-barrier in a dose and time-dependent manner to inhibit centrally mediated effects of ang II (Gohlke et al., 2001). The presence of endogenous brain ang II system, capable of producing its own ang II to stimulate ang II receptors inside the blood brain barrier and not accessible to circulating ang II was well established (Saad et al., 2006a, b, 2005; Phillips and Sumners, 1998; Saavedra, 1992; Ganten et al., 1984). Recently, brain renin angiotensin system has been implicated in additional functions including regulation of cerebral blood flow and cerebroprotection, stress, depression, seizure, nociception, alcohol consumption, memory consolidation, with possible roles in the aetiology of Alzheimer's Disease (AD), Parkinson's disease and diabetes (Abdel-Salam et al., 2007; Wright and Harding, 2011). Numerous anatomical, physiological and pharmacological studies provided evidence for interaction between brain ang II and central monoaminergic systems (Stancheva et al., 2009). The role of ang II as a modulator of brain neurotransmitter systems suggests the therapeutic value of ang II-receptor antagonists not only as antihypertensive drugs but also as drugs useful in neuropsychiatric disorders associated with altered neurotransmitter activity (Stancheva et al., 2009). Ang II is known to stimulate catecholamine release mediated by an AT1 subtype (Pandi et al., 2007; Zimmerman, 1981) is located on presynaptic nerve terminals. Critchley and his colleagues (Critchley et al., 2004) have found that AT1 receptor antagonist candesartan and the ACE inhibitor ramipril, increased basal catecholamine release from the anaesthetized dog's adrenal gland along with decreases in blood pressure. However, it has also been shown that AT2 receptors stimulation induces catecholamine secretion in cultured porcine chromaffin cells (Takekoshi et al., 2001). Thus, there seems to be some controversy about the effect of AT1 receptor blockade on the catecholamine secretion. Lim et al. (2010) demonstrated that olmesartan at low concentrations inhibits the catecholamine secretion evoked by cholinergic stimulation as well as by direct membrane depolarization from the rat adrenal medulla but at high concentrations it rather potentiates the acetylcholine-evoked catecholamine secretion.
It has been reported that the peptide and non-peptide ang II receptor antagonists altered the level of the biogenic monoamines such as dopamine, noradrenaline and serotonin in the frontal cortex, striatum, hypothalamus and hippocampus of rats. However, there are only limited data concerning the effects of ang II receptor antagonists on the activity of the noradrenergic neurotransmitter system (Stancheva et al., 2002). In case of AT1 receptor blockade, this is supposed to reduce catecholamine release leading to endogenous depression, since deficiency of aminergic transmission in the CNS might be causative of endogenous depression (Hardman et al., 2001; Okpako, 2002). But on converse, losartan an AT1 receptor blocker elicits biphasic responses in the forced swim test i.e., decrease immobility at lower doses (0.1, 1.0 and 5 mg kg-1, i.p.) and increase at higher dose (20 and 100 mg kg-1, i.p.) (Vijayapandi and Nagappa, 2005). Telmisartan has the strongest affinity to AT1 receptors among various angiotensin receptor blockers (Kakuta et al., 2005) that has not been hitherto reported for its effect on immobility in mice forced swim test. So the present study is designed to evaluate the effect of telmisartan on immobility using forced swim test in mice.
MATERIALS AND METHODS
Drugs and chemicals: Telmisartan (Glenmark Pharma, India), fluoxetine hydrochloride (Sun Pharma, India), haloperidol (RPG Life Sciences Ltd., India), reserpine (Sisco Research Laboratories Pvt. Ltd., India), reboxetine (Wockhardt Ltd., India) were used. Telmisartan and reboxetine were suspended in carboxy methyl cellulose solution (CMC, 1% w/v) and administered orally by using oral feeding needle. Haloperidol and fluoxetine hydrochloride were dissolved in normal saline and administered intraperitoneally. Reserpine was dissolved in a few drops of glacial acetic acid and the volume was made up with normal saline and given intraperitoneally. All the drug solutions were prepared afresh at beginning of each experiment. All the drugs administered intraperitoneally were given in a constant volume of 1 mL/100 g body weight.
Animals: Male Swiss albino mice (20-25 g) were used. Animals were allowed food and water ad libitum up to the time of experimentation. Prior to use, the mice were housed in polypropylene cages in groups of 6 animals under natural light-dark cycle. Institutional Animal Ethics Committee (IAEC) approved the experimental protocol and care of the animals was taken as per guidelines of CPCSEA, India.
Forced swim test: The behavioral despair test has been used as a test of depressive-like behavior. The animals were forced to swim individually in glass cylinder (30 cm height, 22.5 cm diameter) containing 15 cm water at room temperature. The animals were individually trained in 15 min sessions, using the apparatus described above one day prior to the experimentation. During experimentation each animal was placed into the cylinder one at a time and left there for 6 min. The duration of immobility of each mouse was recorded. A mouse was judged to be immobile when it ceased struggling and remaining floating motionless in the water and making minimal movements necessary to keep its head above water (Pandy et al., 2009). Observation was done by an experienced behavioral psychopharmacologist (H.S.) who was blind to the treatment. All experiments were conducted between 8 a.m to 2 p.m.
Statistical analysis: Results are presented as Mean±SEM. Experimental data in, were analysed by one way Analysis of Variance (ANOVA) followed by post hoc comparisons between drug treated at various time intervals and vehicles treated control using Dunnett’s test. The experimental data were also analysed by one way analysis of variance followed by student’s t- test. Statistical significance was set at p<0.05.
RESULTS AND DISCUSSION
Effect of telmisartan per se and in combination with fluoxetine hydrochloride, reserpine, haloperidol and reboxetine on forced swimming induced immobility in mice is shown in Table 1 and 2. Telmisartan per se perorally at different doses 0.01, 0.05, 0.1, 1, 5, 10, 20 mg kg-1 showed bell-like shaped dose response (i.e.,) increased immobility at 0.05, 0.1, 1, 5, 10 and no change at 0.01, 20 mg kg-1 in the mice forced swim test (Table 1). As shown in Table 2, telmisartan (0.1, 1 and 5 mg kg-1) reversed fluoxetine, reboxetine- induced immobility and potentiated reserpine, haloperidol-induced immobility in mice forced swim test. Animals treated with telmisartan (0.1 mg kg-1) with reserpine (1 mg kg-1) were died due to acute toxicity.
Angiotensin II, either acting directly, or by modulating the activity of other neurotransmitters, has been implicated in a range of cognitive and behavioural processes in animals and humans. The pharmacological interruption of central angiotensin II activity has been shown to have both antidepressant- like and anxiolytic-like effects. Evidence exists to implicate angiotensin II in the aetiology and treatment of depressive illness (Gard, 2002). In the light of earlier case reports, angiotensin-converting enzyme inhibitors have shown antidepressant- like effects. In hypertensive patients who also suffered from depression, it was noted that, unlike other antihypertensive therapies, the angiotensin-converting enzyme inhibitor captopril elicited a significant improvement in mental state (Zubenko and Nixon, 1984; Deicken, 1986; Germain and Chouinard, 1988). Similar results were reported in learned helplessness, an animal model of antidepressant activity. In mice, a single intraperitoneal dose of captopril 10 or 30 mg/kg reduced immobility to the same extent as the reduction produced by the proven antidepressants imipramine (30 mg kg-1) and mianserin (3 mg kg-1) intraperitoneal administration (Giardina and Ebert, 1989).
| Table 1: | Effect of telmisartan (Tel) per se on immobility in mice forced swim test |
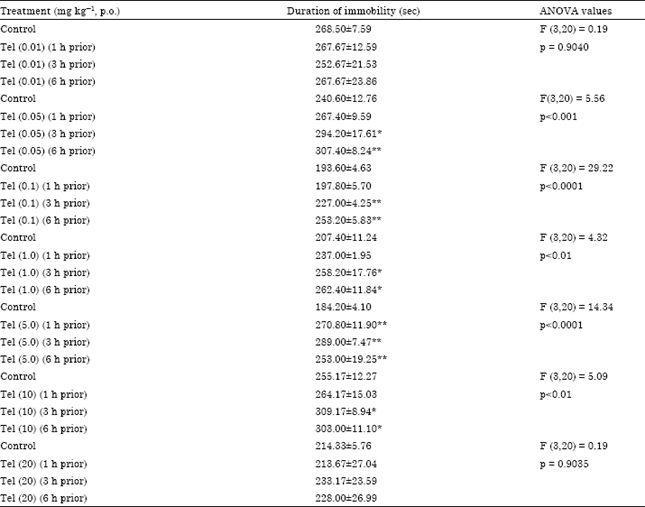 | |
| Six minutes test, Values are Means±SE of 6 animals in each group, *p<0.05, **p< 0.01 (one-way ANOVA / Dunnett’s test: As compared to control group) | |
It was further reported losartan elicited antidepressant activity in the forced swim test in mice (Gard et al., 1999). But on converse, losartan potassium produced biphasic effects on immobility in mice despair swim test (i.e.,) reduced immobility at lower dose (0.1, 1 and 5 mg kg-1) enhanced immobility at higher dose (20 mg kg-1, i.p.). It was reported that the obtained biphasic effects of losartan potassium on immobility in mice might be due to inhibitory effect on AT1 receptor at lower dose and pronounced effect on AT2 receptor at higher dose (Vijayapandi and Nagappa, 2005).
Telmisartan has the strongest affinity to AT1 angiotensin receptors and it can easily penetrate the blood brain barrier (Gohlke et al., 2001). Telmisartan is the most potent sartan regarding the inhibition of sympathetic neurotransmission (Balt et al., 2001). The results of the present study showed the dose-dependent depression-like activity of telmisartan on immobility in mice. (i.e.) it enhanced immobility time at doses 0.05-10 mg kg-1. The depression- like activity was further confirmed by drug interaction studies with reserpine, fluoxetine, haloperidol and reboxetine-induced immobility in mice.
The brain has at least two major angiotensin II receptor subtypes, type I (AT1) and type II (AT2) (Saad et al., 2007a, b; Wright and Harding, 1995).
| Table 2: | Interaction of telmisartan (Tel, 3 h prior) (p.o.) upon fluoxetine (Fluox) (i.p.), reserpine (Res) (i.p.), haloperidol (Hal) (i.p.) and reboxetine (Rebox) (p.o.)-induced immobility in mice forced swim test |
 | |
| Six minutes test. Values are Means±SE of 6 animals in each group. p-values *p<0.05, **p<0.01, ***p<0.001 as compared between treatments. aControl group, bFluoxetine treated group, cReserpine treated group, dHaloperidol treated group, eReboxetine treated group, #: Deceased due to toxicity | |
Angiotensin II has been documented to act on CNS and regulate the feeding and drinking habit in rats (Eghoghosoa et al., 2011). In mouse central nervous system, the AT1 receptors were found in areas involved in the regulation of fluid homeostasis and hormone release such as subfornical organ and the paraventricular nucleus. AT2 receptors were detected in sensory and motor related areas such as the superior colliculus and the inferior olive (Hauser et al., 1998). Biochemical and electrophysiological evidence suggest that AT1 and AT2 receptors have antagonistic effects. Activation of AT1 and AT2 receptors elicited stimulatory and inhibitory effects, respectively, on mitogen-activated protein kinase in neurons (Sumners and Gelband, 1998; Nahamias and Strosberg, 1995; Zhang and Pratt, 1996). In catecholaminergic neurons, ang II elicited an AT1 receptor mediated decrease in the delayed rectifier K+ current (IK) followed by an AT2 receptor mediated increase in IK (Sumners and Gelband, 1998). These studies collectively suggest that functional interaction between the two receptor subtypes have a key role in ang II induced neuromodulatory actions in neurons. The AT2 receptor can act as AT1 receptor antagonist (AbdAlla et al., 2001). The AT2 receptor is the first identified example of a G-protein coupled receptor which acts as a receptor-specific antagonist. The inhibitory effects of AT2 receptor as been attributed to the AT2 mediated activation of protein phosphatases. AT2, in the brain plays important role in some types of stress-induced hyperthermia in mice (Watanabe et al., 1999). AT2 is expressed in locus coeruleus which supplies a noradrenergic projection to many brain areas and is thought to regulate arousal, attention, anxiety, affective and pain responses (Ashton, 1987). This may imply a functional interaction between AT2 sites and norepinephrine in the brain. In tests of anxiolytic behavior, there may be an initial, short lived AT2 receptor mediated increase in anxiety- related behavior is produced (Gard, 2002). AT2 receptors in the adrenal medulla, participate in the inhibition of ang II-induced catecholaminergic release resulting from AT1 blockade (Jezova et al., 2003). Angiotensin II via AT2 receptor stimulates IK, hyperpolarizes the cell which would counter-balance the neuronal activity and turn off of evoked release of norepinephrine (Gelband et al., 1997). Stimulation of AT2 receptors reduces catecholamine biosynthesis via decrease in cGMP levels (Takekoshi et al., 2002).
The depression- like effect produced by telmisartan might be due to the activation of AT2 receptors, since AT1 receptor antagonist specifically block the effect of ang II on the AT1 receptor and redirect the effects of ang II to the unopposed AT2 receptor. Therefore stimulation of AT2 receptor as an indirect result of AT1 receptor antagonism may contribute to the depression-like effect of telmisartan. Depression is an affective disorder, due to decrease in monoamine levels in the brain. The unopposed AT2 receptor stimulation might decrease the release of monoamines. It has been previously reported that unopposed stimulation of AT2 receptor might cause long term adverse effects (Jackson and Garrison, 1996). So it might be a probable mechanism by which telmisartan produce depression- like effect in despair swim test.
Reversal of antidepressant effect of fluoxetine (selective serotonin reuptake inhibitor), reboxetine (selective noradrenaline reuptake inhibitor) by telmisartan demonstrated functional interaction of telmisartan with adrenergic and serotonergic system in the brain. In addition, telmisartan potentiated haloperidol (dopamine D2 receptor antagonist), reserpine (a monoamine depletor) -induced immobility confirms its interaction with dopaminergic system. The present study revealed the depression- like activity of telmisartan in mice forced swim test mediated through neuromodulatory effect on brain monoaminergic system.
Telmisartan dissociates very slowly from the receptors once it is bound and unlike candesartan it does not re-associate with receptor after dissociation. This slow dissociation may account for the insurmountable profile of the drug (Maillard et al., 2002). In the present study, telmisartan showed long lasting effect on immobility even at 6 h after treatment which might be due insurmountable binding to AT1 receptor. Angiotensin II, time-dependently dissociate telmisartan, olmesartan, candesartan, valsartan, losartan and active metabolite of losartan, EXP 3174 from membrane components containing human AT1 receptor: the dissociation constants of each ARB was 0.003248, 0.004171, 0.005203, 0.009946, 0.01027 and 0.008561 min-1 with corresponding half-life (t 1/2) of 213, 166, 133, 70, 67 and 81 min, respectively (Kakuta et al., 2005). It was also demonstrated that telmisartan could have the strongest affinity to AT1 receptors among various ARBs examined. The rank order of affinity was shown as telmisartan>olmesartan > candesartan > EXP 3174 = valsartan = losartan. The interaction of telmisartan with ang II type I (AT1) or type II (AT2) receptors was determined in radio ligand experiments (Wienen et al., 2000). Inhibition of 125I-labeled ang II binding by telmisartan revealed a high affinity of this compound for AT1 receptors in rat lung preparations with Ki value of 3.7 nM. In contrast, the AT1 selective antagonist losartan and its active metabolite, EXP 3174 are less potent with Ki values of 23.7 nM and 10.4 nM, respectively (Entzeroth and Hadamovsky, 1991). Losartan potassium significantly enhanced immobility time in mice only at higher doses (20 and 100 mg kg-1, i.p.) (Vijayapandi and Nagappa, 2005). But the enhancement in immobility time was noted even at lower doses (0.05 mg kg-1) of telmisartan which might be due to high affinity of telmisartan for AT1 receptors.
The present study results demonstrated the telmisartan showed depression- like effect on immobility in mice forced swim test. The interaction of telmisartan with the monoaminergic modulating drugs such as fluoxetine, reboxetine, reserpine and haloperidol on mice forced swim test further confirms the interaction of brain RAS with monoaminergic system. However, the present study results on mice contradict in current clinical settings. Since telmisartan is well tolerated by patients and no major adverse effects were noticed till date. So these discrepancies might be due to species difference. Normally, telmisartan is administered to the patients in chronic fashion. Chronic treatment of telmisartan might attenuate depression-like effect in patients. So further chronic behavioural animal studies are called for to confirm its CNS effectiveness.
CONCLUSION
The present study elicits depression-like activity in mice forced swim test which might be mediated through neuromodulatory mechanism on brain monoaminergic system. The therapeutic potential of telmisartan as a new psychopharmacological agent to modulate brain monoaminergic neurotransmitter system may warrant further exploration.
ACKNOWLEDGMENTS
The authors are grateful to The Principal, KMCH College of Pharmacy and Dr. Thavamani D. Palaniswami, Secretary, Kovai Medical Centre Research and Education Trust, Coimbatore for providing encouragement and necessary lab facilities to carry out this project and Prof. Datin Dr. Zahurin Mohamed, The Head, Department of Pharmacology, University of Malaya, Kuala Lumpur, Malaysia for providing facility to write this manuscript.
REFERENCES
- AbdAlla, S., H. Lother, A.M. Abdel-tawab and U. Quitterer, 2001. The angiotensin II AT2 receptor is an AT1receptor antagonist. J. Biol. Chem., 276: 39721-39726.
CrossRef - Akhavan, M.M., M. Karimi, M. Ghodrati and H. Falahtpishe, 2011. AT1 receptors activation enhances the expression of MMP-2, MMP-13 and VEGF but not MMP-9 in B16F10 melanoma cells. Pak. J. Biol. Sci., 4: 821-830.
CrossRef - Balakumar, P. and M. Singh, 2007. Recent advances in pharmacotherapy for heart failure: Future directions. Trends Med. Res., 2: 61-71.
CrossRefDirect Link - Balt, J.C., M.J. Mathy, M. Pfaffendrof and P.A. van Zwieten, 2001. Inhibition of angiotensin II induced facilitation of sympathetic neurotransmission in the pithed rat: A comparison between losartan, irbesartan, telmisartan and captopril. J. Hypertens., 19: 465-473.
PubMed - Eghoghosoa, A.R., O.L. Aderemi, S.A. Olufemi and O.I. Peter, 2011. Effect of angiotensin I-converting enzyme inhibitor, captopril, on body weight and food and water consumption in oral contraceptive-treated rats. Am. J. Biochem. Mol. Biol., 1: 95-100.
CrossRefDirect Link - Gelband, C.H., M. Zhu, D. Lu, L.P. Reagan and S.J. Fluharty et al., 1997. Functional interactions between neuronal AT1 and AT2 receptors. Endocrinology, 138: 2195-2198.
PubMedDirect Link - Gohlke, P., S. Weiss, A. Jansen, W. Wienen and J. Stangier et al., 2001. AT1 receptor antagonist telmisartan administered peripherally inhibits central responses to angiotensin II in conscious rats. J. Pharmacol. Exp. Ther., 298: 62-70.
PubMed - Huq, F., 2007. Molecular modeling analysis of the metabolism of irbesartan. J. Pharmacol. Toxicol., 2: 238-247.
CrossRef - Kakuta, H., K. Sudoh, M. Sasamata and S. Yamagishi, 2005. Telmisartan has the strongest binding affinity to angiotensin type 1 receptor. Comparison with other angiotensin II type 1 receptor blockers. Int. J. Clin, Pharmacol. Res., 25: 41-46.
PubMed - Abdel-Salam, O.M.E., S. El-Shenawy and S.M. Nofal, 2007. Effect of ramipril, valsartan and candesartan on thermal and visceral pain in mice. J. Pharmacol. Toxicol., 2: 533-541.
CrossRefDirect Link - Pandy, V., N. Jose and H. Subhash, 2009. CNS activity of methanol and acetone extracts of Acorus calamus leaves in mice. J. Pharmacol. Toxicol., 4: 79-86.
CrossRefDirect Link - Pandi, V., A.N. Nagappa and P.A. Thakurdesai, 2007. Effects of losartan potassium on central dopaminergic system in mice. J. Pharmacol. Toxicol., 2: 551-558.
CrossRefDirect Link - Phillips, M.I. and C. Sumners, 1998. Angiotensin II in central nervous system physiology. Regul. Pept., 78: 1-11.
PubMed - Saad, W.A., L.A. de Arruda Camargo, J. Antunes-Rodrigues, W.A. Saad, I.F.M.S. Guarda and R.S. Guarda, 2005. Angiotensin and arginine vasopressin receptor subtypes of the lateral preoptic area effect on the sodium balance. J. Biological Sci., 5: 562-567.
CrossRefDirect Link - Saad, W.A., I.F.M.S. Guarda, L.A. de Arruda Camargo, W.A. Saad, R.S. Guarda, T.A.F.B. Santos and S. Simoes, 2006. Endothelial and neuronal nitric oxide synthase inhibitors influences angiotensin II pressor effect in central nervous system. Int. J. Pharmacol., 2: 341-347.
CrossRefDirect Link - Saad, W.A., I.F.M.S. Guarda, L.A.D.A. Camargo, R.S. Guarda, W.A. Saad and T.A.F.B.D. Santos, 2006. Influence of arginine vasopressin receptor and nitric oxide on the water, sodium intake and arterial blood pressure induce by angiotensin injected into third ventricle of the brain. J. Biological Sci., 6: 182-186.
CrossRefDirect Link - Saad, W.A., I.F.M.S. Guarda, L.A. de Arruda Camargo, T.A.F.B. dos Santos and W.A. Saad, 2007. Effects of nitric oxide and arginine vasopressin on water intake induced by central angiotensin II. Part 1. J. Medical Sci., 7: 845-849.
CrossRefDirect Link - Saad, W.A., I.F.M.S. Guarda, L.A. de Arruda Camargo, T.A.F.B. dos Santos and W.A. Saad, 2007. Effects of nitric oxide and arginine vasopressin on sodium intake induced by central angiotensin II. Part 2. J. Boil. Sci., 7: 806-810.
CrossRefDirect Link - Stancheva, S., L. Alova and M. Stefanova, 2009. Effect of peptide and nonpeptide antagonists of angiotensin II receptors on noradrenaline release in hypothalamus of rats with angiotensin II-induced increase of water intake. Pharmacol. Rep., 61: 1206-1210.
PubMed - Stancheva, S., L. Alova, M. Velkova and V. Georgiev, 2002. The effects of peptide and nonpeptide antagonists of angiotensin II receptors on the level of brain biogenic monoamines in rats with angiotensin II-induced increase of water intake. Methods Find Exp. Clin. Pharmacol., 24: 287-290.
CrossRefPubMed - Takekoshi, K., K. Ishii, S. Shibuya, Y. Kawakami, K. Isobe and T. Nakai, 2002. Angiotensin II type 2 receptor counter-regulates type 1 receptor in catecholamine synthesis in cultured porcine adrenal medullary chromaffin cells. Hypertension, 39: 142-148.
PubMed - Takekoshi, K., K. Ishii, Y. Kawakami, K. Isobe and T. Nakai, 2001. Activation of angiotensin II subtype 2 receptor induces catecholamine release in an extracellular Ca2-dependent manner through a decrease of cyclic guanosine 3`,5`-monophosphate production in cultured porcine adrenal medullary chromaffin cells. Endocrinology, 142: 3075-3086.
CrossRefPubMed - Wienen, W., M. Entzeroth, C. Jacobus, A. VanMeel and J. Stangier et al., 2000. A review on Telmisartan: A novel, long-acting angiotensin II receptor antagonist. Cardiovasc. Drug Rev., 18: 127-154.
CrossRef - Wright, J.W. and J.W. Harding, 1995. Brain angiotensin receptor subtypes AT1, AT2 and AT 4 and their functions. Regul. Pept., 59: 269-295.
CrossRefDirect Link - Zimmerman, B.G., 1981. Adrenergic facilitation by angiotensin: Does it serve a physiological function?. Clin. Sci., 60: 343-348.
PubMed - Zubenko, G.S. and R.A. Nixon, 1984. Mood-elevating effect of captopril in depressed patients. Am. J. Psychiatry, 141: 110-111.
PubMed