
Tania R. Stoyanoff
SEGCyT-CONICET, Faculty of Medicine, National Northeast University
Juan S. Todaro
SEGCyT, Faculty of Medicine, National Northeast University
Maria V. Aguirre
Department of Biochemistry, Faculty of Medicine, National Northeast University. Moreno 1240 (3400), Corrientes, Argentina
Nora C. Brandan
Department of Biochemistry, Faculty of Medicine, National Northeast University. Member of Research Career, CONICET- Argentina
Journal of Pharmacology and Toxicology
Year: 2012 | Volume: 7 | Issue: 8 | Page No.: 344-358







ABSTRACT
Adriamycin (ADR)-induced nephropathy in rodents is an experimental model commonly used for studies of chronic human renal diseases. The molecular associations involved in renal apoptosis linked to hypoxia and cell stress response in this model are not completely known. The aim of this study was to determine the associations among the expression patterns of the Inducible Nitric Oxide Synthase (iNOS), the heat shock protein 60 (Hsp60) and the Hypoxia Inducible Factor-1 Alpha (HIF-1α) linked to apoptosis in renal cortex in the nephrotic syndrome progression induced by ADR administration. Male BALB/c mice were treated with a single dose of ADR (11 mg kg-1; i.v.). Tubulointerstitial nephrosis was monitored by histopathological assessment and by biochemical determinations on 7, 15 and 30 days following drug exposure. These results were evaluated in conjunction with renal expression of iNOS, Hsp60 and HIF-1α. Cortical apoptosis was examined by TUNEL assay. The increment of renal apoptotic cells in tubulointerstitial areas was accompanied by the decrease in Bcl-xL/Bax ratio and the enhancement of the active caspase-3 and Hsp60 expressions from day 7 onwards. iNOS and HIF-1α increased concomitant with the renal apoptosis and the tubule interstitial injury. Taking the previous information into account, data indicate that the over expression of renal HIF-1α, iNOS and Hsp60 are concurrent with the apoptotic events triggered by ADR. These results contribute to additional knowledge of the molecular complex events involved in the context of ADR-induced nephropathy progression.
PDF Abstract XML References Citation
Received: October 26, 2012;
Accepted: February 12, 2013;
Published: February 23, 2013
How to cite this article
Tania R. Stoyanoff, Juan S. Todaro, Maria V. Aguirre and Nora C. Brandan, 2012. Cell Stress, Hypoxic Response and Apoptosis in Murine Adriamycin-induced Nephropathy. Journal of Pharmacology and Toxicology, 7: 344-358.
DOI: 10.3923/jpt.2012.344.358
URL: https://scialert.net/abstract/?doi=jpt.2012.344.358
DOI: 10.3923/jpt.2012.344.358
URL: https://scialert.net/abstract/?doi=jpt.2012.344.358
INTRODUCTION
The fact that Adriamycin (ADR) is a well-known inducer of renal injury in rodents, mirrors what is seen in the human chronic kidney disease. Animals treated with ADR develop a nephrotic and tubular syndrome, a model able to characterize mechanisms of progressive renal disease (Bertani et al., 1982; Okuda et al., 1986; Wang et al., 2000). The induction of apoptosis is an important cytotoxic mechanism of ADR (Muller et al., 1998). Renal tubular apoptotic cells have been observed in ADR-treated rats (Zhang et al., 1996) and programmed cell death of renal tubules is a characteristic of tubular atrophy (Schelling et al., 1998; Khan et al., 1999).
In addition, low oxygen tension plays a key role in the establishment of renal disease (Nangaku, 2004) and it also induces apoptosis in murine ADR-nephrosis (Kairaitis et al., 2005). It is well known that the Hypoxia Inducible Factor-1α (HIF-1α) is a crucial mediator during cell response post hypoxic challenge in experimental kidney diseases (Tanaka and Nangaku, 2009).
Several mechanisms seem to account for the toxic effects of ADR (Lee and Harris, 2011) which suggests that the cause of this anthracycline induced toxicity is a multifactorial and complex process. One of the mechanisms of ADR-induced nephrotoxicity is ascribed to oxidative injury resulting either from the generation of free radicals (ROS) and/or Nitric Oxide (NO) which, in turn, cause diverse oxidative damage on critical cell components, such as plasma membranes and mitochondria (Barbey et al., 1989; Oktem et al., 2010). Furthermore, the NO produced by Inducible Nitric Oxide Synthase (iNOS) has been implicated in the pathogenesis of many glomerulopathies in animal models, as a signalling molecule regulating apoptosis and inflammation (Wang et al., 1997; Ozen et al., 2001).
It is well known that the exposure to stressors can alter the physiological tissular proteome with the expression induction of specific proteins (i.e., heat shock proteins) and the translation suppression of usually occurring ones (Bukau and Horwich, 1998; Beck et al., 2000). Moreover, it has been described that if the level of stress is too high to be attenuated by the cell stress response, cell death can be ensued by necrotic and/or apoptotic pathways.
The intracellular chaperone heat shock protein 60 (Hsp60), induced by monocytes/macrophages under cellular stress, has been proposed to be a danger signal leading to cell demise (Zheng et al., 2004; Lang et al., 2005).
The downstream regulation of HIF-1α on the iNOS gene (Hierholzer et al., 2001) and the role of NO in the HIF-1α system (Brune and Zhou, 2003) are well known. However, uncertainty remains with regard to the apoptotic events in the context of HIF-1α, Hsp60 and iNOS expressions in ADR-induced nephropathy.
Because cells often use similar molecular defences against xenobiotics, more elucidation related to the underlying molecular mechanisms of this model may provide insights into the biological pathways relevant to ADR-nephrotoxicity.
In the present time course study of ADR nephropathy, HIF-1α, iNOS and Hsp60 were chosen as selective markers for the assessment of tissular hypoxia, NO generation and cellular response to stressful stimuli, respectively.
Therefore, this in vivo study is the first report that establishes the association among iNOS, Hsp60 and HIF-1α expressions in tubular apoptosis in the nephrotic syndrome progression induced by ADR administration.
MATERIALS AND METHODS
Animal treatment and experimental design: Experimental protocol involved the use of an in vivo murine model of ADR-induced nephropathy in a 30 days study period. Assays were performed using inbred male BALB/c mice (22-25 g, age: 6-8 weeks) from the animal facility of Northeast National University, Argentina. All animals were kept at a constant temperature (22±2°C) with 12 h light/dark cycles and had free access to standard diet and distilled water. Local ethical animal committee approval was obtained before the study was started. The animals were randomly divided into two treatment groups which received a single intravenous injection of each drug into the tail vein. Group 1 was injected with 11 mg kg-1 of Adriamycin (ADR) (Doxorubicin hydrochloride, Pharmacia and Upjohn, Argentina). Group 2 was treated with sterile saline and acts as a control group (Day 0). At 0, 7, 15 and 30 days post treatment, mice (n = 6-9/time point) were anesthetized with pentobarbital (60 mg kg-1) intraperitoneally and were bled by heart puncture. After being sacrificed by cervical dislocation, the kidneys were quickly excised and washed with cold saline. Renal samples were used in routine histology, immunoblottings, iNOS immunohistochemistry, TUNEL and caspase-3 activity assays while serums were used in biochemical determinations.
All experimental procedures were conducted following the Guide for the Care and Use of Laboratory Animals (National Institute of Health, Bethesda, MD, USA).
Renal function assessment: Serum Creatinine (sCr), albumin and Blood Urea Nitrogen (BUN) concentrations were determined with a Synchron CX7 autoanalyzer (Beckman, CA). Proteinuria was assessed with spot urine dipsticks (Multistix 10SG, Bayer) as negative or trace, 1+(0.30 g L-1), 2+(1 g L-1), 3+(3 g L-1) or 4+(20 g L-1).
Histopathological examination: Renal tissue samples were fixed in 4% paraformaldehyde and embedded in paraffin. Sections (4 μm thick) were processed according to Periodic Acid-Schiff (PAS) staining. Histopathological evaluation was confirmed by two independent histopathologists unaware of the experimental design. Semiquantitative assessment of glomerular and tubulointerstitial injury was performed using scores ranging from 0 to 3, as follows: 0, normal kidney; 1, mild changes; 2, moderate changes; 3, severe changes. The scores were determined in each section selected at random and 20 fields were examined under 40 x magnification. Images were taken using an Olympus Coolpix-microdigital camera fitted to a CX-35 microscope (Olympus, Japan).
In situ cell death detection (TUNEL assay): Terminal deoxynucleotidyl transferase-mediated deoxyuridin Triphosphate Nick End Labeling (TUNEL) assay was performed using an in situ Cell Death Detection Kit (Roche, Indianapolis, IN, USA) according to the manufacturer’s instructions. Brown labeled TUNEL positive cells were counted underx400 magnification. The apoptotic index was calculated as the percentage of TUNEL-positive cells/total number of renal cells.
Western blot analysis: Expressions of Bax, Bcl-xL, Hsp60, iNOS and cleaved caspase-3 were determined by immunoblotting from cytosolic renal extracts as previously described (Aquino-Esperanza et al., 2008). Briefly, whole kidneys were homogenized and lysed into ice-cold buffer [10 mM HEPES pH 7.4, 10 mM KCl, 1.5 mM MgCl2, 0.5 mM dithiothreitol, 0.1% IGEPAL (Sigma Co, MO, USA)], supplemented with a protease inhibitor cocktail. Cell lysates were centrifuged at 14,000 g and the supernatant was used as cytosolic fraction.
The nuclear pellets were gently resuspended with ice-cold-wash buffer (20 mM HEPES pH 7.4, 1.5 mM MgCl2, 420 mM ClNa, 25% glycerol, 0.2 mM EDTA [ethylene glycol-bis(beta-aminoethyl ether)-N.N.N’.N’-tetraacetic acid], 0.5 mM DDT), 0.2 mM PMSF with protease inhibitors and incubated for 1 h at 4°C. Samples were centrifuged at 16,000 g at 4°C for 30 min and the supernatants were collected as nuclear extracts for HIF-1α immunoblotting.
Cytosolic or nuclear proteins (40 μg) were resolved into 8 or 12% SDS-PAGE. They were blotted onto nitrocellulose membranes (Bio-Rad, CA, USA) and probed with 1:500 dilutions of primary antibodies: Anti-Bax, anti-Bcl-xL, anti-Hsp60 (Santa Cruz Biotechnology, USA), anti-cleaved caspase-3 (Cell Signaling Technology, USA), anti-iNOS (Cayman Chemicals, MI, USA), anti-HIF-1α (Novus Biologicals, Littleton, CO) and anti-β actin (Sigma-Aldrich). Membranes were then incubated with horseradish peroxidase-conjugated secondary antibodies (Jackson Immunoresearch Inc, USA).
Immunocomplexes were detected by an Opti4CN kit (Bio-Rad, CA, USA). Band optical density (OD) was analyzed using Scion Image 3.0 Software. Results are expressed as the ratio: (protein of interest OD/β-actin OD).
Immunohistochemistry: Fixed renal sections (4 μm thick) were deparafinized, rehydrated through a graded ethanol series using routine protocols. Endogenous peroxidase activity was blocked with 3% H2O2 for 15 min and the sections were pre-treated in citrate buffer (pH 6.0) in a microwave for antigen unmasking. They were incubated with a rabbit polyclonal iNOS antibody (Cayman Chemical, USA, dilution 1:50) for 18 h at 4°C. iNOS was detected using anti-rabbit biotinylated secondary antibody and a streptavidin-biotin horseradish peroxidase complex (DAKO kit) with diaminobenzidine as the substrate. Negative control samples were processed as described above except for the primary antibody which was omitted and replaced with buffer phosphate saline. Slides were counterstained with hematoxylin and the immunostaining was evaluated with light microscopy.
Caspase-3 activity: Caspase-3 activity was measured in the supernatants of renal samples with a commercially available caspase-3 assay kit (Sigma, St. Louis, MO, USA) as previously described (Aispuru et al., 2008). Briefly, proteolytic reactions were carried out in extraction buffer containing 20 μg of cytosolic protein extract and 40 μM Ac-DEVD-pNA. The reaction mixtures were incubated at room temperature for 2 h and the formation of pNA was measured at 405 nm. Experiments were performed in triplicate. Caspase-3 activity was calculated as fold increase of untreated ADR samples (Day 0) at each time point of the study.
Statistical analysis: Differences between group means were estimated using a one-way Analysis of Variance (ANOVA) followed by Dunnett’s t-test. Analyses were carried out using the software package Instat and Prism 4.0 (GraphPad software Inc., San Diego, USA). A p-value <0.05 was considered statistically significant.
RESULTS
Functional and pathological effects in ADR-induced nephropathy: All experimental animals developed progressive renal disease characterized by proteinuria, hypoalbuminemia and impaired renal function.
| Table 1: | Time course of renal functional changes in ADR-treated and control mice |
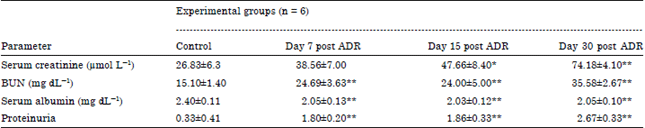 | |
| ADR: Adriamycin, BUN: Blood urea nitrogen, SEM: Standard error of the mean, n: No. of mice, Data are presented as Mean±SEM, *p<0.05, **p<0.01 compared with control group. One-way ANOVA statistical technique was used to analyze the differences between groups at 0.05 as level of significance. Post-hoc Dunnet test was performed | |
Blood urea nitrogen [BUN] (15.10±1.40 vs 24.69±3.63 mg dL-1, p<0.01) and proteinuria increased from day 7 until the end of the experimental schedule (0.33±0.41 vs 1.80±0.20 mean scores, p<0.01). Serum creatinine enhanced significantly from day 15 onwards (26.83±6.3 vs 47.66±8.40 μmol L-1, p<0.05) reaching the maximal value on day 30 (74.18±4.10 μmol L-1, p<0.01) supporting the impairment of renal function (Table 1).
Histopathological examination of renal sections revealed a gradual glomerular, tubular and interstitial injury, all of which were more severe on day 30 post ADR injection. Progressive tubular atrophy involved vacuolization, many intratubular casts and a stepwise reduction in tubular cell height. Glomeruli were reduced in size. The expansion of the mesangium was observed through the increase in PAS-positive material. Moreover, a mild interstitial infiltration of mononuclear cells was noticed at the end of the experimental period (Fig. 1a-d).
Histophatological changes were assessed through semiquantitative scores for glomerulosclerosis and tubular damage (Fig. 1e). Scores for glomerulosclerosis increased progressively from day 15 onwards, reaching the maximal on day 30 (3.66±0.22 vs 0.30±0.17, p<0.001). Scores for tubular damage enhanced significantly from day 7 (p<0.05), reaching the maximum at the end of the study (3.66±0.22 vs 0.76±0.25, p<0.001).
Tubular cell apoptosis in ADR-induced nephropathy: For localization and quantification of apoptotic cells within the kidney, TUNEL assay was performed on renal tissue sections. Figure 2a, shows there were very few apoptotic cells in the kidneys of the control group. In ADR-treated group, apoptotic cells were mainly located in tubular and interstitial areas. Moreover, apoptosis was scarcely noted in the glomerular and vascular areas of the kidney (Fig. 2b). Compared with the control group, semi-quantitative analysis revealed that apoptotic cells in tubulointerstitial areas (per 400x field) were gradually increased by ADR (Fig. 2c).
Expression of Bcl-xL/Bax: The functional balance of pro-apoptotic versus anti-apoptotic members of the Bcl-2 protein family determines whether a cell will live or die. Hence, Bax and Bcl-xL expressions were evaluated in kidney samples following the experimental schedule. The analysis of the blots revealed that the Bax expression increased progressively during the study (p<0.01) (Fig. 3a). On the other hand, the immunoreactivity of Bcl-xL decreased from day 15 onwards (Fig. 3b).
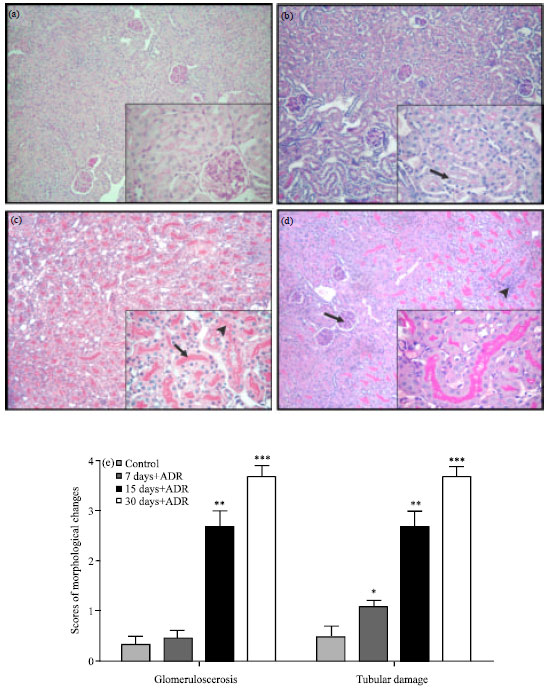 | |
| Fig. 1(a-e): | Histopathology of renal cortices in Adriamycin (ADR)-induced nephropathy, (a) Control mice: Saline treated shows normal organization of glomeruli and tubules, (b) ADR treated mice: Focal increase of tubular vacuolization on day 7, (c) ADR treated mice: ADR Tubular atrophy reflected by intraluminal casts (arrow) on day 15, vacuolization and a stepwise reduction in tubular cell height (arrowhead), (d) ADR treated, arrow: Increment in periodic acid Schiff (PAS)-positive material, indicative of glomerulosclerosis, was detected in the glomeruli on day 30, arrowhead Mild interstitial infiltration was found and (e) Semiquantitave scores of renal injury are increased after ADR injection compared with the control group, Values are Mean±SEM, *p<0.05, **p<0.01, ***p< 0.001 vs. control group |
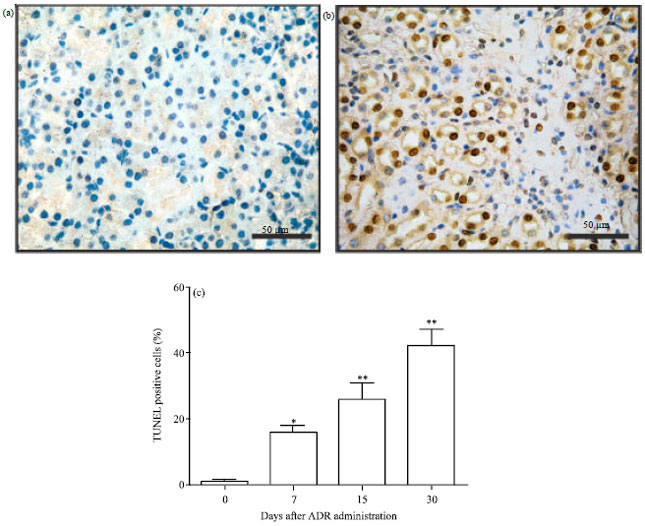 | |
| Fig. 2(a-c): | Renal apoptosis in Adriamycin induced nephropathy (a) Control mice, (b) Day 30 post ADR treatment and (c) Apoptotic percentages in renal sections post ADR treatment, Values are mean±SEM of three animals, *p<0.05, **p<0.01 vs. control group |
Consequently, the ADR-treated group revealed a shift of the Bcl-xL/Bax ratio to a pro-apoptotic effect in parallel with the development of nephropathy (Fig. 3c).
Caspase-3 active expression and activity: To determine the involvement of the active forms of caspase-3 in ADR nephropathy, caspase-3 immunoblotting and the enzymatic activity assay were performed. Experimental data indicate that the cleaved active forms of caspase-3 (20 and 17 kDa) were over expressed from day 7 to day 30, coincident with the maximal apoptosis (Fig. 4a). In addition, the caspase-3 activity assay in renal cell lysates (Fig. 4b) showed an increment of 2-3 folds over control in accordance with the blotting profiles.
iNOS expression: Free radical production and/or NO release induced by ADR are thought to be responsible for the injuring effects of this anthracycline. Therefore, the expression of iNOS was studied by immunoblotting in order to elucidate the link between NO generation and the pathophysiological renal progression post ADR administration. Figure 5a shows that iNOS was markedly upregulated between days 15 and 30 (p<0.01).
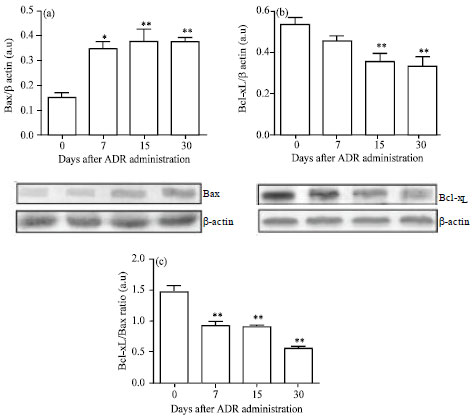 | |
| Fig. 3(a-c): | Bcl-xL and Bax expressions in Adriamycin (ADR)-induced nephropathy. Representative immunoblots of Bax (a) and Bcl-xL (b) are shown. Data are normalized to β-actin as loading control for both proteins. Bars (c) represent the Bcl-xL/Bax ratio. Values are Mean±SEM of six mice per group, *p<0.05, **p<0.01 vs. Control group |
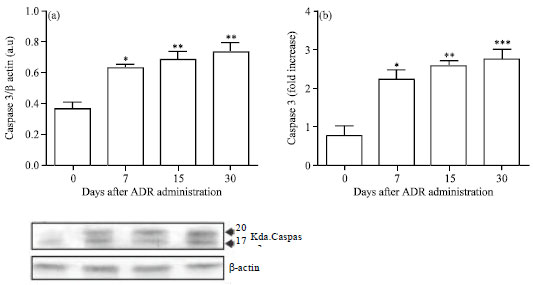 | |
| Fig. 4(a-b): | Caspase-3 expression and caspase-3 activity assay in renal cells following Adriamycin treatment (a) Immunoblots of cleaved caspase-3 expression in ADR-induced nephropathy and (b) Caspase-3 activity assay. Caspase-3 activity was measured with a colorimetric assay kit that relies on caspase-mediated cleavage of p-nitroanilide (pNA) from a synthetic caspase substrate peptide with the amino acid sequence: Asp-Glu-Val-Asp (DEVD), Values are Mean±SEM of six mice per group, *p<0.05, **p<0.01, ***p< 0.001 vs. control group |
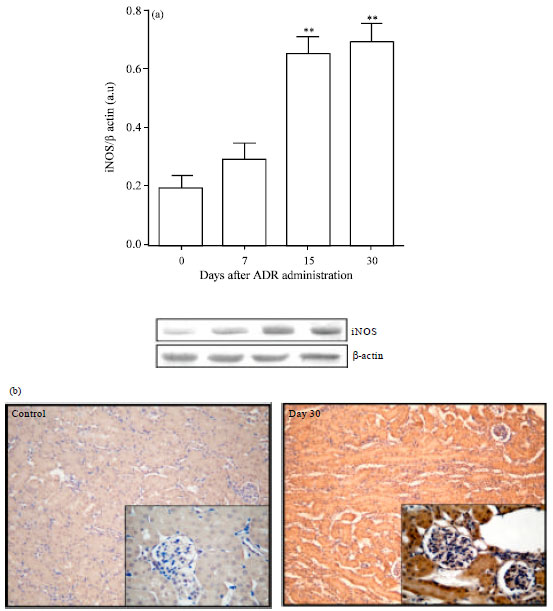 | |
| Fig. 5(a-b): | iNOS expressions in renal cells post Adriamycin (ADR) treatment. (a) Immunoblottings of iNOS. Data are normalized to β-actin as loading control. Values are Mean±SEM of six mice per group. ** p<0.01 vs. control group. (b) Photomicrographs of iNOS immunoreactivity corresponding to control mice and day 30 post ADR. iNOS over expression was mainly noticed in renal tubular cells. Magnification 100 x and 400 x, respectively |
Besides this, immunohistochemical examination of iNOS revealed that ADR administration caused significant increase of the iNOS immunoreactivity in the cytoplasm of renal tubular cells and in some glomerular cells compared with the control group (Fig. 5b). These findings showed that iNOS over-expression was coincident with tubule-interstitial injury and renal apoptosis.
Hsp60 expression: To investigate whether Hsp60 induction occurred in this experimental model and its relationship with apoptosis, the intracellular Hsp60 expression was studied by Western blotting. As illustrated in Fig. 6, renal Hsp60 was over expressed from day 7 to day 30 (p<0.01). Data reveals that Hsp60 can act as a danger signal and it contributes to the programmed death cell in ADR induced nephropathy, since its increment was related to the maximal apoptotic indexes.
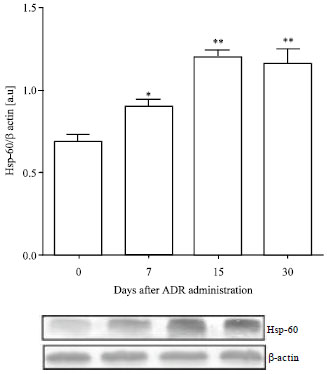 | |
| Fig. 6: | Hsp60 expressions in renal cells post Adriamycin treatment. Data are normalized to β-actin as loading control. Values are Mean±SEM of six mice per group. *p<0.05, ** p<0.01 vs. control group |
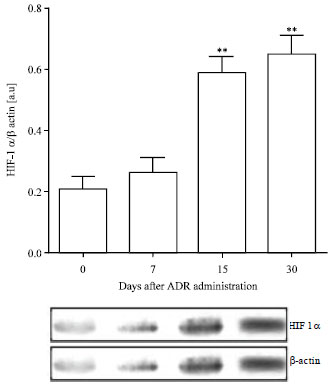 | |
| Fig. 7: | Hypoxia inducible factor (HIF-1α) expression in renal cells post Adriamycin treatment. Data are normalized to β-actin as loading control, Values are Mean±SEM of six mice per group, ** p<0.01 vs. control group |
HIF-1α expression: HIF-1α was evaluated by Western blotting analysis to observe changes in its expression in renal tissue during the hypoxic response induced by ADR (Fig. 7).
HIF-1α expression was over expressed from day 15 until the end of the experimental study (p<0.01). The enhancement of HIF-1α (15 to 30 days) was coincident with the renal functional impairment, the tubule-interstitial injury and the maximal apoptotic indexes at the end of this experimental schedule.
DISCUSSION
Apoptosis plays a central role not only in the physiological processes of kidney growth and remodeling but also contributes to the tubular cell demise in a variety of settings; including ischemic, toxic and septic acute tubular necrosis, as well as in chronic tubular atrophy; which is a characteristic of chronic renal diseases (Ortiz et al., 2002).
This time course study attempts to describe associations between renal apoptosis in the context of the hypoxic and the stressed renal cortex after ADR administration.It is conceivable that in the stressed kidneys the functional disturbance caused by ADR would be reflected in molecular changes. As it was aforementioned, key molecules were chosen to evaluate tissular hypoxia (HIF-1 alpha), NO generation (iNOS) and a classical molecule involved in stress response (Hsp60) in parallel with ADR-induced apoptosis.
Deleterious effects of ADR on glomerular endothelial cells have been linked to the development of proteinuria (Jeansson et al., 2009) that consequently leads to a progressive tubulointerstitial scarring (Turnberg et al., 2006).
The findings of this study show an overt proteinuria that appeared within the first week following ADR administration and remain significantly increased with progressive renal pathological and functional changes throughout the experimental period. The impaired renal function is associated with the classical described histopathological damages (i.e., tubular casts, mild glomerulosclerosis and tubular epithelial cell vacuolization) and with differential profiles in the expression of several molecules related to apoptosis, hypoxia and oxidative stress.
The apoptosis has been reported in renal proximal tubular and kidney epithelial rat cell lines under ADR effect (Chen et al., 2006) as well as in renal tubular cells in ADR-treated rats (Zhang et al., 1996). Renal tubular epithelial cell apoptosis causes tubular atrophy, a feature of renal disease progression. Moreover, the mitochondria have also been identified as one of the targets in the ADR-induced subcellular damage (Oktem et al., 2010).
The balance between the pro-apoptotic (Bax, Bak) and the anti-apoptotic (Bcl-2, Bcl-xL) members of the Bcl-2 family of proteins determines the ability of cells to either survive or undergo the activation of the executioner procaspase-3, conducing to the subsequent programmed cell death (Wei et al., 2001; Youle and Strasser, 2008; Brenner and Mak, 2009; Aquino-Esperanza et al., 2008). An increment in Bax/Bcl-2 or Bax/Bcl-xL ratios may activate caspase-3 and modulates the apoptotic pathway in a variety of experimental settings (Yang et al., 2002; Chien et al., 2007; Aispuru et al., 2008; Klemm et al., 2008).
Previous studies have revealed that the apoptosis of glomerular cells after ADR administration was accompanied by an increase of Bax and a reduction of Bcl-2 expressions (Liu et al., 2007). Furthermore, increased cleaved caspase-3 levels and decreased Bcl-xL expression have been observed in cultured podocytes ADR-induced apoptosis (Zhou et al., 2011). In line with these reports, the results of this study indicate that the progressive increment of renal cell apoptosis is concomitant with the decrease in the Bcl-xL/Bax ratio, the over expression of the active caspase-3 forms (17 and 20 Kda) and the maximal caspase-3 activity. Moreover, the results obtained show that TUNEL-positive cells are to a larger extent localized in the tubule-interstitial areas and are scarcely noted in the glomeruli, in agreement with findings (Ozen et al., 2001).
In addition to the severe glomerular and tubulointerstitial damages induced by ADR, the peritubular capillary loss and the reduced blood flow oxygen supply lead to chronic interstitial and tubular cell hypoxia. Therefore, HIF-1α is stabilized in the progressive renal disease and it might be related to interstitial fibrosis. This fact is in accordance to previous reports which emphasize that chronic renal hypoxia is a final common pathway to the end-stage kidney injury (Kairaitis et al., 2005; Nangaku and Eckardt, 2007). However, it must be considered that several factors relevant to renal injury, including NO and TNF-α, also induce nonhypoxic HIF-1α stabilization (Sandau et al., 2001).
It has been communicated that hypoxia induces apoptosis of renal tubular cells with tubular atrophy in a murine model (Khan et al., 1999). According to this report, the present study shows that the nuclear HIF-1α over expression was coincident with the renal functional impairment, the tubulointerstitial damage and the apoptotic enhancement at the end of the experimental setting.
The link between NO production and apoptosis has been assigned to several processes. In addition to the direct cytotoxicity of NO (Dalkara and Moskowitz, 1996), it has been reported that iNOS contributes to DNA fragmentation and programmed cell death (Wang et al., 1997). The experimental data from Western blotting and immunohistochemistry of iNOS reinforce the association between iNOS over expression and the apoptotic enhancement for the most part in renal tubular cells from day 15 to 30 following ADR treatment.
In this time course study, it is worth mentioning that both iNOS and HIF-alpha expressions enhanced progressively with the tubule interstitial injury and the renal apoptosis. On the other hand, heat shock proteins are essential for cell function under physiological situations and contribute decisively to the survival of cells subjected to adverse environmental conditions. They have long been suspected to act as danger signals in stressed kidneys. The Hsp60-Hsp10-chaperonins system is coupled to the binding and hydrolysis of ATP. In eukaryotic cells, Hsp60 is localized primarily in the matrix space of mitochondria, where in cooperation with mitochondrial Hsp70, it assists in the folding and assembly of imported proteins (Beck et al., 2000).
It has been reported that ADR induced the over-expression of Hsp70 (Strauss and Porras, 2007). However, unlike Hsp 27,-70 and-90 with widely described pro-survival functions, Hsp60 is somewhat controversial with both pro-survival and pro-apoptotic functions depending on the injured context (Chandra et al., 2007). The present time course study of Hsp60 expression provides new in vivo evidence that this chaperone plays a pro-apoptotic function accompanying caspase-3 activation, Bax enhancement and Bcl-xL decrease during ADR-induced nephropathy. Moreover, this study is in agreement with another experimental setting which demonstrated that hypoxia caused the disassociation of the Hsp60-Bax complex with translocation of cytosolic Hsp60 to the plasma membrane and Bax to the mitochondria with the consequent triggering of the apoptotic pathway (Gupta and Knowlton, 2002).
CONCLUSION
The data presented herein suggest that changes in the ratio of Bax to Bcl-xL may contribute to the caspase-3 activation and that renal apoptosis is associated with iNOS, Hsp60 and HIF-1α over expression in the complex scenario of ADR induced nephropathy.
In summary, these observations provide additional insights into the cellular and molecular apoptotic events involved in the context of renal hypoxia and cell stress response in the progressive renal disease induced by this anthracycline.
ACKNOWLEDGMENTS
This study was supported by grants from SEGCyT UNNE (PI 17/0060) and by funds from CONICET Argentina (PIP 1302).The authors are grateful to Drs. P. Teibler and Dr. V. Cerdera for the histopathological assessments and to Prof. M. Delfino for assistance with English revision.
REFERENCES
- Aispuru, G.R., M.V. Aguirre, J.A. Aquino-Esperanza, C.N. Lettieri, J.A. Juaristi and N.C. Brandan, 2008. Erythroid expansion and survival in response to acute anemia stress: The role of EPO receptor, GATA-1, Bcl-xL and caspase-3. Cell Biol. Int., 32: 966-978.
PubMed - Aquino-Esperanza, J.A., M.V. Aguirre, G.R. Aispuru, C.N. Lettieri, J.A. Juaristi, M.A. Alvarez and N.C. Brandan, 2008. In vivo 5-flourouracil-induced apoptosis on murine thymocytes: Involvement of FAS, Bax and Caspase3. Cell Biol. Toxicol., 24: 411-422.
CrossRef - Barbey, M.M., L.M. Fels, M. Soose, K. Poelstra, W. Gwinner, W. Bakker and H. Stolte, 1989. Adriamycin affects glomerular renal function: Evidence for the involvement of oxygen radicals. Free Radic. Res. Commun., 7: 195-203.
PubMed - Beck, F., W. Neuhofer and E. Muller, 2000. Molecular chaperones in the kidney: Distribution, putative roles and regulation. Am. J. Renal. Physiol., 279: F203-F215.
Direct Link - Bertani, T., A. Poggi, R. Pozzoni, F. Delaini and G. Sacchi et al., 1982. Adriamycin-induced nephrotic syndrome in rats: Sequence of pathologic events. Lab. Invest., 46: 16-23.
PubMed - Brune, B. and J. Zhou, 2003. The role of nitric oxide (NO) in stability regulation of hypoxia inducible factor-1 α (HIF-1 α). Curr. Med. Chem., 10: 845-855.
PubMed - Bukau, B. and A.L. Horwich, 1998. The Hsp70 and Hsp60 chaperone machines. Cell, 92: 351-366.
CrossRefPubMedDirect Link - Chandra, D., G. Choy and D.G. Tang, 2007. Cytosolic accumulation of HSP60 during apoptosis with or without apparent mitochondrial release. J. Biol. Chem., 282: 31289-31301.
PubMed - Chien, C.T., S.K. Shyue and M.K. Lai, 2007. Bcl-xL augmentation potentially reduces ischemia/reperfusion induced proximal and distal tubular apoptosis and autophagy. Transplantation, 84: 1183-1190.
PubMed - Gupta, S. and A.A. Knowlton, 2002. Cytosolic heat shock protein 60, hypoxia and apoptosis. Circulation, 106: 2727-2733.
PubMed - Hierholzer, C., B.G. Harbrecht, T.R. Billiar and D.J. Tweardy, 2001. Hypoxia-inducible factor-1 activation and cyclo-oxygenase-2 induction are early reperfusion-independent inflammatory events in hemorrhagic shock. Arch. Orthop. Trauma Surg., 121: 219-222.
PubMed - Jeansson, M., K. Bjorck, O. Tenstad and B. Haraldson, 2009. Adriamycin alters glomerular endothelium to induce proteinuria. J. Am. Soc. Nephrol., 20: 114-122.
CrossRef - Kairaitis, L.K., Y. Wang, M. Gassmann, Y.C. Tay and D.C. Harris, 2005. HIF-1alpha expression follows microvascular loss in advanced murine adriamycin nephrosis. Am. J. Physiol. Renal. Physiol., 288: 198-206.
PubMed - Khan, S., R.P. Cleveland, C.J. Koch and J.R. Schelling, 1999. Hypoxia induces renal tubular epithelial cell apoptosis in chronic renal disease. Lab. Invest., 79: 1089-1099.
PubMed - Klemm, K., C. Eipel, D. Cantre, K. Abshagen, M.D. Menger and B. Vollmar, 2008. Multiple doses of erythropoietin impair liver regeneration by increasing TNF-α, the Bax to Bcl-xL ratio and apoptotic cell death. PLoS One, Vol. 3.
Direct Link - Lang, A., D. Benke, F. Eitner, D. Engel and S. Ehrlich et al., 2005. Heat shock protein 60 is released in immune-mediated glomerulonephritis and aggravates disease: In vivo evidence for an immunologic danger signal. J. Am. Soc. Nephrol., 16: 383-391.
PubMed - Muller, I., D. Niethammer and G. Bruchelt, 1998. Anthracycline-derived chemotherapeutics in apoptosis and free radical cytotoxicity. Int. J. Mol. Med., 1: 491-494.
PubMed - Nangaku, M., 2004. Hypoxia and tubulointerstitial injury: A final common pathway to end-stage renal failure. Nephron. Exp. Nephrol., 98: e8-e12.
CrossRefDirect Link - Oktem, G., A. Uysal, O. Oral, E.D. Sezer and M. Olukman et al., 2010. Resveratrol attenuates doxorubicin-induced cellular damage by modulating nitric oxide and apoptosis. Exp. Toxicol. Pathol., 64: 471-479.
CrossRefPubMedDirect Link - Okuda, S., Y. Oh, H. Tsuruda, K. Onoyama, S. Fujimi and M. Fujishima, 1986. Adriamycin-induced nephropathy as a model of chronic progressive glomerular disease. Kidney Int., 29: 502-510.
CrossRef - Ortiz, A., P. Justo, M.P. Catalan, A.B. Sanz, C. Lorz and J. Egido, 2002. Apoptotic cell death in renal injury: The rationale for intervention. Curr. Drug Targets Immune. Endocr. Metabol. Disord., 2: 181-192.
Direct Link - Ozen, S., Y. Usta, I. Sahin-Erdemli, D. Orhan and B. Gumusel et al., 2001. Association of nitric oxide production and apoptosis in a model of experimental nephropathy. Nephrol. Dial. Transplant, 16: 32-38.
Direct Link - Schelling, J.R., N. Nkemere, J.B. Kopp and R.P. Cleveland, 1998. Fas dependent fratricidal apoptosis is a mechanism of tubular epithelial cell deletion in chronic renal failure. Lab. Invest., 78: 813-824.
PubMed - Strauss, M. and N. Porras, 2007. Differential expression of HSP70 and ultraestructure of heart and liver tissues of rats treated with adriamycin: Protective role of L-carnitine. Invest. Clin., 48: 33-43.
PubMedDirect Link - Tanaka, T. and M. Nangaku, 2009. Drug discovery for overcoming Chronic Kidney Disease (CKD): Prolyl-hydroxylase inhibitors to activate Hypoxia-Inducible Factor (HIF) as a novel therapeutic approach in CKD. J. Pharmacol. Sci., 109: 24-31.
PubMedDirect Link - Turnberg, D., M. Lewis, J. Moss, Y. Xu, M. Botto and H.T. Cook, 2006. Complement activation contributes to both glomerular and tubulointerstitial damage in adriamycin nephropathy in mice. J. Immunol., 177: 4094-4102.
PubMedDirect Link - Wang, Y., Y.P. Wang, Y.C. Tay and D.C.H. Harris, 2000. Progressive adriamycin nephropathy in mice: Sequence of histologic and immunohistochemical events. Kidney Int., 58: 1797-1804.
CrossRef - Wei, M.C., W.X. Zong, E.H. Cheng, T. Lindsten and V. Panoutsakopoulou et al., 2001. Pro-apoptotic BAX and BAK: A requisite gateway to mitochondrial dysfunction and death. Science, 292: 727-730.
PubMedDirect Link - Yang, B., T.S. Johnson, G.L. Thomas, P.F. Watson, B. Wagner, P.N. Furness and A.M. El Nahas, 2002. A shift in the Bax/Bcl-2 balance may activate caspase-3 and modulate apoptosis in experimental glomerulonephritis. Kidney Int., 62: 1301-1313.
CrossRefPubMedDirect Link - Youle, R.J. and A. Strasser, 2008. The BCL-2 protein family: Opposing activities that mediate cell death. Nat. Rev. Mol. Cell Biol., 9: 47-59.
CrossRefPubMedDirect Link - Zheng, L., M. He, M. Long, R. Blomgran and O. Stendahl, 2004. Pathogen-induced apoptotic neutrophils express heat shock proteins and elicit activation of human macrophages. J. Immunol., 173: 6319-6326.
PubMed - Zhou, Y., X. Kong, P. Zhao, H. Yang and L. Chen et al., 2011. Peroxisome proliferator-activated receptor-α is renoprotective in doxorubicin-induced glomerular injury. Kidney Int., 79: 1302-1311.
PubMed - Sandau, K.B., J. Zhou, T. Kietzmann and B. Brune, 2001. Regulation of the hypoxia-inducible factor 1α by the inflammatory mediators nitric oxide and tumor necrosis factor-α in contrast to desferroxamine and phenylarsine oxide. J. Biol. Chem., 276: 39805-39811.
CrossRef