
Chinaka O. Nwaehujor
Department of Biochemistry, Faculty of Basic Medical Sciences, University of Calabar, Calabar, Nigeria
Okwoche J. Ode
Department of Veterinary Pharmacology and Toxicology, Faculty of Veterinary Medicine, University of Abuja, Nigeria
Florence C. Nwinyi
Department of Veterinary Pharmacology and Toxicology, Faculty of Veterinary Medicine, University of Abuja, Nigeria
Nkeiruka E. Udeh
Department of Veterinary Physiology and Pharmacology, Faculty of Veterinary Medicine, Michael Okpara University of Agriculture, Umudike, Abia State, Nigeria
Journal of Pharmacology and Toxicology
Year: 2012 | Volume: 7 | Issue: 4 | Page No.: 181-191







ABSTRACT
The hypoglycemic and anti-oxidant effects of the methanolic extract of Buchholzia coriacea fruits in streptozotocin-induced diabetic rats were investigated. It was tolerated up to dose of 2000 mg kg-1 orally in rats. Treatment with 150, 300 and 600 mg kg-1 of the extract induced significant (p<0.05) dose dependent decreases in the respective fasting blood glucose values of 6.61+2.14, 5.12+1.01 and 3.84+0.81 mmol L-1 from the initial mean value of 14.15±5.12 mmol L-1 when compared with the negative control. Glibenclamide (2 mg kg-1) however, lowered the fasting blood glucose to 3.51±0.75 mmol L-1. The serum concentrations of catalase and reduced glutathione were significantly (p<0.05) higher in rats treated with glibenclamide (2 mg kg-1) and various doses (150, 300 and 600 mg kg-1) of the extract in contrast to the values in negative control rats. The extract also decreased serum triglycerides and total serum cholesterol. The fruit extract dose dependently reduced lipid peroxidation in diabetic rats. The ability of scavenging free radicals was measured by DPPH reduction and Ferric Reducing/Antioxidant Power (FRAP) spectrophotometric assays. In the DPPH assay, the highest concentration (400 μg mL-1) of the extract produced 66% antioxidant activity compared to 80% with ascorbic acid at the same concentration. The FRAP value increased concentration dependently from 1.1 to 1.7 μM (100-400 μg mL-1) while ascorbic acid has a FRAP value of 2 μM at 1000 μg mL-1. The findings suggest that the extract could be a potential source of a novel anti-diabetic and antioxidant agent.
PDF Abstract XML References Citation
Received: February 04, 2012;
Accepted: March 21, 2012;
Published: June 11, 2012
How to cite this article
Chinaka O. Nwaehujor, Okwoche J. Ode, Florence C. Nwinyi and Nkeiruka E. Udeh, 2012. Effects of Methanol Extract of Buchholzia coriacea Fruit in Streptozotocin-induced Diabetic Rats. Journal of Pharmacology and Toxicology, 7: 181-191.
DOI: 10.3923/jpt.2012.181.191
URL: https://scialert.net/abstract/?doi=jpt.2012.181.191
DOI: 10.3923/jpt.2012.181.191
URL: https://scialert.net/abstract/?doi=jpt.2012.181.191
INTRODUCTION
Diabetes mellitus is a metabolic disorder that is rapidly reaching epidemic proportions and the World Health Organization has predicted that by 2025, 300 million people will be affected worldwide (Kopelman and Hitman, 1998; Amos et al., 1997). Diabetes eventually leads to diseases of the coronary arteries and the cerebrovascular system, renal failure, blindness, neurological complications and premature death (Harris et al., 1987; Jayatilake et al., 1993). Two forms of diabetes mellitus, Type 1 and Type 2 have been identified. Type 1 diabetes mellitus is primarily due to the autoimmune-mediated destruction of pancreatic β-cells of the islets, resulting in absolute insulin deficiency. People with type 1 diabetes mellitus must rely on exogenous insulin to prevent the development of ketoacidosis for survival. It has been noted that the number of people with Type 1 diabetes is dramatically increasing worldwide (Pavana et al., 2008).
The incidence of Type 1 diabetes mellitus is low relative to Type 2 which accounts for more than 90% of the diabetic cases globally. Type 2 diabetes mellitus is characterized by a failure of normal insulin levels to stimulate glucose uptake by tissue cells. People with Type 2 diabetes mellitus are not dependent on exogenous insulin. Generally speaking, excessive production and reduced utilization of glucose by tissues are the basic cause of diabetes (Chattopadhyay, 1993). Available treatment regimens include insulin, oral anti-hyperglycemic agents and inhibitors of a-glycosidase enzyme although alternative therapies are also used including plant products (Sabo et al., 1999). Nearly, 80% of rural African communities use ethnotherapy to control or treat diabetes mellitus. Several plants have been studied and were found to possess anti-diabetic and anti-oxidant properties (Al-Hader et al., 1993; Al-Enazi, 2007). The use of these plants is based on the belief that they have low toxicity and cost less than the semi-synthetics or synthetics drugs.
Buchholzia coriacea belongs to the Capparaceae family (Keay, 1989). The seeds gave the plant its common name of “wonderful kola” because of its popular usage in traditional medicine. The seeds are covered in a purple aril which is chewed in Ivory Coast and is known to have a sharp pungent taste. The plant is documented to possess diverse medicinal potentials. Okoli et al. (2010) reported the anti-plasmodial properties of the plant, the ground seeds were therefore routinely mixed with palm oil and taken orally as treatment for malaria (Adjanohoun et al., 1996). The Cameroonians use the seed as remedy to relieve chest pain (Thomas et al., 1989). It was also reported to have analgesic effects (Ezeja et al., 2011) and anthelminthic potentials (Nweze and Asuzu, 2006). The aim of the study was to evaluate the possible anti-diabetic and anti-oxidant activities of extracts of B. coriacea fruit using in vivo and in vitro experimental models.
MATERIALS AND METHODS
Plant collection and extraction: The fresh fruits of B. coriacea were plucked from its tree in Nsukka Local Government Area of Enugu State, Nigeria and identified by a qualified botanist (taxonomist) from the Bioresources Development and Conservation Programme (BDCP) Nsukka, Enugu state. The fruits were cut into small pieces, dried and pulverised into coarse powder. Extraction was done by cold maceration. One kilogram of the plant material was extracted in 80% methanol for 72 h with intermittent shaking at 2 h interval. The extract was then filtered using Whatman No. 1 filter paper. The filtrate was concentrated using rotary evaporator (Rotavapor R 210, Büchi, Switzerland) at 40°C. The yield was determined and the extract stored in a refrigerator.
Acute toxicity study: Five groups of Wistar rats of both sexes with each group containing five rats were used. Four groups were treated orally with varying doses of the Buchholzia coriacea fruit extract at 250, 500, 1000 and 2000 mg kg-1, respectively. Group 5 was given an equivalent volume of distilled water to serve as control. The animals were observed for toxic signs like excitability, dullness, diarrhoea, in-appetence and death over 72 h.
Experimental animals: Male Wistar rats with average weight of 122-137 g were used as test animals. They were obtained from the Laboratory Animal Unit of the Faculty of Veterinary Medicine, University of Nigeria, Nsukka. The animals were housed in cages at room temperature and under a light period of 16-18 h daily for a period of 2 weeks prior to the commencement of the experiment. Standard commercial rat pellets (Grand Cereals Ltd. Jos, Nigeria) and water were provided ad-libitum. The laboratory animals were used in accordance with laboratory practice regulation and the principle of humane laboratory animal care as documented by Zimmermann (1983).
Screening of Buchholzia coriacea for anti-diabetic activity: All rats were fasted overnight before the administration of streptozotocin (STZ) obtained from Sigma Aldrich, USA. Freshly prepared STZ dissolved in 0.1 M citrate buffer and pH 4.5 was given by a single intraperitoneal injection at a dose of 50 mg kg-1. Control rats were injected with citrate buffer only. After 3 days post challenge with STZ, blood glucose levels were measured. Rats with blood glucose level above 9 mmol L-1 were considered diabetic and were selected. They were randomly grouped into five groups of five animals each. Group I, II and III were treated daily with the extract of B. coriacea dissolved in distilled water at doses of 150, 300 and 600 mg kg-1, respectively. Group IV was treated daily with a standard drug, Glibenclamide (2 mg kg-1) and group V received distilled water only.
At Day 7 and 14 of treatment, blood samples were collected from the rat tail vein. The fasting blood sugar level was measured using the Accu-chek Advantage II glucometer. Percentage reductions in fasting blood glucose level in experimental animals were calculated using the formula given below:
Blood was collected from the rats on day 14 of treatment for the antioxidant and biochemical assays.
In vivo antioxidant activities of B. coriacea and serum lipid biochemical assays
Assay of catalase (CAT) activity: Catalase activity was measured according to the method of Aebi (1984). A given volume (0.1 mL) of the serum was pipetted into cuvette containing 1.9 mL of 50 mM phosphate buffer of pH 7.0. Reaction was started by the addition of 1.0 mL of freshly prepared 30% (v/v) hydrogen peroxide (H2O2). The rate of decomposition of H2O2 was measured spectrophotometrically from changes in absorbance at 240 nm. The enzyme activity was expressed as units mL-1 protein.
Assay of reduced glutathione (GSH) concentration: Reduced glutathione was determined by the method of Ellman (1959). A volume (1.0 mL) of serum was treated with 0.5 mL of Ellman’s reagent (19.8 mg of 5, 5- dithiobis-2-nitrobenzoic acid (DTNB) in 100 mL of 0.1% sodium nitrate) and 3.0 mL of phosphate buffer (0.2 M, pH 8.0). Then 0.4 mL of distilled water was added. The mixture was thoroughly mixed; absorbance was read at 412 nm and expressed as units mL-1.
Estimation of lipid peroxidation (malondialdehyde): Lipid peroxidation in the plasma was estimated colorimetrically as thiobarbituric acid reactive substances (TBARS) using the method of Buege and Aust (1978). A principle component of TBARS is malondialdehyde (MDA), a product of lipid peroxidation. In brief, 0.1 mL of tissue homogenate (tris-HCl buffer, pH 7.5) was treated with 2 mL (1:1:1 ratio) of TBA-TCA-HCl reagent (thiobarbituric acid 0.37%, 0.25 N HCl and 15% TCA). The mixture was placed in water bath for 15 min; it was then allowed to cool. The absorbance of clear supernatant was measured against reference blank at 535 nm. Concentration was expressed as nmol mL-1. This method is based on the principle that acetic acid detaches the lipid and protein of a tissue, thiobarbituric acid reacting with lipid peroxide, hydrogen peroxide and oxygen-labile double bond to form the colour adducts with maximal absorbance at 530 nm.
Serum lipid biochemical analyses
Serum triglycerides (TG) assay: Determination of serum triglycerides was carried out with Cayman’s Triglyceride Assay kit (Cayman’s Chemical Company, USA). The samples and reagents were freshly prepared as specified and serial dilutions of triglyceride standard diluent were made accordingly. Serum sample from each animal in the groups was prepared on day 14 of treatment. Triglyceride standard diluent was used as the blank. Ten microliter of triglyceride standard was added with pipette into designated wells on a plate and this was followed by the addition of equal volume (10 μL) of serum sample from individual rats into separate wells. The set up was done in duplicates. The reaction was initiated by adding 150 μL of diluted enzyme buffer solution to each well. The micro titre plate was carefully shaken for 5 sec to achieve proper mixing before it was covered with the plate cover. The plate was incubated at room temperature for 15 min before absorbance was read at 540 nm using a plate reader. The concentration of triglycerides in the samples was determined from the linear regression of the standard curve by substituting the corrected absorbance value for each sample into the equation:
Determination of total serum cholesterol: Total serum cholesterol was assayed with total cholesterol kit obtained from Crest Biosystems, India. Cholesterol esterase hydrolyses esterified cholesterols to free cholesterol. The free cholesterol is oxidized to form hydrogen peroxide which further reacts with phenol and 4-aminoantipyrine by the catalytic action of peroxidase to form a red coloured quinone-imine dye complex. Intensity of the colour formed is directly proportional to the amount of cholesterol present in the sample. The absorbance of the Standard (Abs. S) and Test samples (Abs. T) against the Reagent blank was read at 505 nm. Distilled water was used as zero setting in the spectrophotometer. The concentration of cholesterol in the samples was determined from the equation:
In vitro antioxidant activities of B. coriacea
Antioxidant capacity of B. coriacea fruit extract with 1, 1- diphenyl-2-picrylhydrazyl radical (DPPH) spectrophotometric assay: The method of Mensor et al. (2001) was adopted. Two milliliter of test extract at concentrations ranging from 10 μg mL-1 400 μg mL-1 was each mixed with 1 mL of 0.5 mM DPPH (in methanol). Absorbance at 517 nm was taken after 30 min incubation in the dark at room temperature. The concentrations were prepared in triplicates. The percentage antioxidant activity was calculated as follows:
One mL of methanol plus 2 mL of the extract was used as blank while 1 mL of 0.5 mM DPPH solution plus 2 mL of methanol was used as control. Ascorbic acid was used as reference standard.
Ferric reducing/antioxidant power (FRAP) assay: The total antioxidant potential of sample was determined using a ferric reducing ability of plasma (FRAP) assay of Benzie and Strain (1999) as a measure of “antioxidant power”. FRAP assay measures the change in absorbance at 593 nm owing to the formation of a blue colored Fe11-tripyridyltriazine compound from colorless oxidized Fe111 form by the action of electron donating antioxidants. Standard curve was prepared using different concentrations (100-1000 μmol L-1) of FeSO4.7H2O. All solutions were used on the day of preparation. In the FRAP assay the antioxidant efficiency of the extract under the test was calculated with reference to the reaction signal given by an Fe2+ solution of known concentration, this representing a one-electron exchange reaction. The results were corrected for dilution and expressed in μmol Fe11/L. Vitamin C was measured within 1 h after preparation. The sample to be analyzed was first adequately diluted to fit within the linearity range. All determinations were performed in triplicate:
Statistical analysis: All data were expressed as Mean±SEM. Data were analyzed using one way analysis of variance (ANOVA) at 5% level of significance. Dunnett’s test was used to detect the difference among the treatment groups.
RESULTS
Extraction of the plant material: Buchholzia coriacea fruit extract was brownish in colour with an oily consistency. The total solids recovered from extracts were 18% (w/w).
Acute toxicity: The fruit extract of B. coriacea did not cause any mortality up to a dose of 2000 mg kg-1 in the rats treated orally with varying doses (250, 500, 1000 and 2000 mg kg-1) of the crude extract. The extract was well tolerated by the rats with mild signs of toxicity (dizziness) at 2000 mg kg-1.
Anti-diabetic activity of Buchholzia coriacea fruit extract: The methanolic extract of B. coriacea fruits exerted significant (p<0.05) reduction in fasting blood glucose level of diabetic rats at day 7 of treatment. The varying doses (150, 300 and 600 mg kg-1) of the extract produced fasting blood glucose values of 7.52±2.22, 8.02±2.14 and 6.51±1.67 mmol L-1, respectively when compared with 16.74±3.14 mmol L-1 of the distilled water-treated rats (Table 1). Similarly, at day 14 of treatment, 150, 300 and 600 mg kg-1 doses of the extract induced further significant (p<0.05) decreases in the corresponding fasting blood glucose values of 6.61±2.14, 5.12+1.01 and 3.84±0.81 mmol L-1 compared to control which had 14.15±5.12 mmol L-1.
| Table 1: | The effects of methanolic fruit extract of Buchholzia coriacea on fasting blood glucose of streptozotocin-induced diabetic rats |
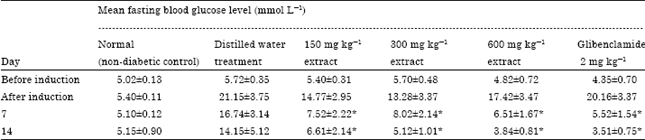 | |
| *Significant decreases at p<0.05 compared to negative control, Values are Mean±SEM, n = 5 in each group | |
| Table 2: | The effects of methanol fruit extract of Buchholzia coriacea on oxidative stress parameters, serum cholesterol and triglycerides of STZ-induced diabetic rats |
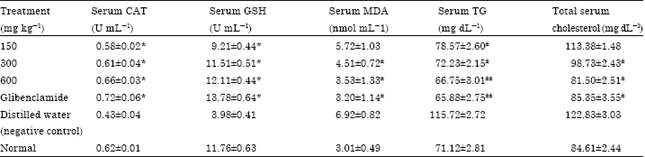 | |
| *Significant (p<0.05) increases compared to negative control, ±,±±Significant decreases compared to negative control at p<0.05 and p<0.01, respectively, values are Mean±SEM. n = 5 in each group | |
Glibenclamide also produced significant (p<0.05) reduction in the mean fasting blood glucose levels of 5.52±1.54 and 3.51±0.75 mmol L-1 at 7 and 14 days of treatment, respectively.
In vivo antioxidant activities of B. coriacea fruit and serum lipid biochemical assays: The serum concentration of catalase and GSH were significantly (p<0.05) higher in rats treated with glibenclamide and various doses (150, 300 and 600 mg kg-1) of the extract in contrast to the values in distilled water treated rats. There was no significant difference (p>0.05) in CAT and GSH levels of rats treated with glibenclamide and the different doses of extract. However, B. coriacea fruit extract induced significant (p<0.05) reduction in MDA values at 300 and 600 mg kg-1 when compared with values in the negative control group which were treated with only distilled water. All the test doses (150, 300 and 600 mg kg-1) of the extract and glibenclamide significantly (p<0.05) reduced total serum triglyceride in the studied animals relative to negative control but the extract was only significantly (p<0.05) effective at 300 and 600 mg kg-1 in decreasing the concentration of total serum cholesterol (Table 2). The suppressive effects of the extract on MDA, serum triglycerides and total serum cholesterol in the experimental rats were found to increase with increasing doses of the extract.
DPPH photometric assay: The crude extract of B. coriacea fruits demonstrated appreciable antioxidant activity with DPPH radical scavenging method. At the lowest concentration (10 μg mL-1), the extract exhibited a mean antioxidant activity of 11% while ascorbic acid produced 76% (Fig. 1). However, at the highest concentration (400 μg mL-1), the extract had 66% antioxidant activity compared to 80% with ascorbic acid at the same concentration.
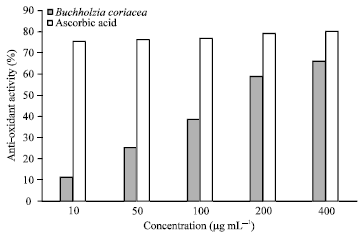 | |
| Fig. 1: | DPPH spectrophotometric assay |
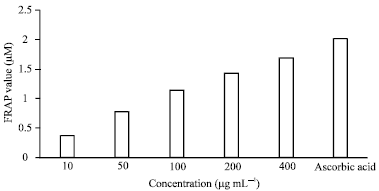 | |
| Fig. 2: | The anti-oxidant activity of B. coriacea methanol fruits extract determined by the FRAP assay |
Ferric reducing/antioxidant power assay (FRAP): The ferric reducing ability of plasma, a measure of the antioxidant ability, showed that the methanol extract of B. coriacea fruits produced a dose dependent antioxidant effect. At 10 μg mL-1, the mean antioxidant power (FRAP value) was 0.37 μM; this increased to 1.1 μM at 100 μg mL-1 and then 1.7 μM at 400 μg mL-1. Ascorbic acid has a FRAP value of 2 μM at 1000 μg mL-1 (Fig. 2).
DISCUSSION
Several plants have been investigated for their anti-diabetic effect and have shown ability to prevent degenerative and metabolic effects in STZ-induced diabetic animal models (Soleimani et al., 2007). Buchholzia coriacea methanol fruit extract was safe up to a dose of 2000 mg kg-1 and did not cause mortality in rats. The crude extract exhibited potent hypoglycemic activities at the test doses (150, 300 and 600 mg kg-1) within the duration of study. The extract exerted its maximal effect at 600 mg kg-1 when mean fasting blood glucose levels in rats were lowered from 6.51±1.67 mmol L-1 on day 7 to 3.84±0.81 mmol L-1 on day 14. This effect was comparable to the action of glibenclamide (2 mg kg-1), a reference anti-diabetic agent which lowered mean fasting blood glucose level of rats from 5.52±1.54 to 3.51±0.75 within the same period.
Streptozotocin (STZ) induces diabetes through β-cell toxicity by inhibition of O-GlcNAC-selective N-acetyl-b-d-glucosaminidase (OglcNAcase), the enzyme that removes O-GlcNAc from protein (Konrad et al., 2001; Palsamy and Malathi, 2007). It has been well reported that partial destruction of beta cells causes a reduction in insulin secretion which is materialized by a chronic hyperglycemia (Ene et al., 2008). Intraperitoneal administration of streptozotocin effectively induces diabetes in normal rats as reflected by glycosuria, hyperglycaemia and body weight loss when compared with normal rats (Nirmala et al., 2008). Glibenclamide, a sulfonylurea is a popular anti-diabetic drug used widely to treat Type 2 diabetes and works by inhibiting adenosine triphosphate (ATP)-sensitive potassium channels in pancreatic beta cells. This inhibition causes cell membrane depolarization and opening of voltage-dependent calcium channels, thus triggering an increase in intracellular calcium into the beta cell which stimulates insulin release. The specific mechanism by which the crude extract of B. coriacea fruit induced its hypoglycemic effect in diabetic rats was not fully understood but it was possible that the extract mediated its action either through the prevention of STZ-induced O-GlcNAcase inhibition or indirect stimulation of insulin release or early regeneration of β-cells of the islets. STZ-induced β-cell toxicity is completely blocked by GlcNAc, which also prevents STZ-induced O-GlcNAcase inhibition (Konrad et al., 2001). The O-linked glycosylation is reversed by OglcNAc β-acetylglucosaminidase (O-GlcNAcase) (Kaneto et al., 2001). In peripheral tissues such as muscle, the hexosamine biosynthetic pathway serves as a glucose sensor linking the rate of glucose flux to the activity of the glucose transport system and transcription machinery (Dong and Hart, 1994; Luo et al., 2007).
B. coriacea fruit extract decreased serum triglycerides in a dose dependent manner and had a similar reducing effect on total serum cholesterol from 300 to 600 mg kg-1 (Table 2). Under normal circumstances insulin activates enzyme lipoprotein lipase and hydrolyses triglycerides thus, increasing uptake of fatty acids into adipose tissue and triglyceride synthesis. It also inhibits lipolysis (Subash-Babu and Ignacimuthu, 2007; Priya et al., 2012). Deficiency of insulin results in failure to activate the enzymes thereby causing hypertriglyceridemia (Mahesh and Brahatheeswaran, 2007). The abnormal high concentration of serum lipids in the diabetic untreated rats may be due, mainly to the increase in the mobilization of free fatty acids from the peripheral fat depots, since insulin inhibits the hormone sensitive lipase (Jasmine and Daisy, 2007).
Several physiological and pathological conditions such as diabetes, ageing, inflammation, viral infections and neurodegenerative diseases may develop through the action of reactive oxygen species (Al-Omar et al., 2004). The crude methanol extract of B. coriacea fruits demonstrated appreciable antioxidant activities as revealed by the in vivo (CAT and GSH) and in vitro (DPPH, FRAP and lipid peroxidation) antioxidant studies. There was dose-dependent preservation of endogenous serum catalase and reduced glutathione by the graded doses (150, 300 and 600 mg kg-1) of the extract. Diabetic rats that were treated with extract and glibenclamide had higher concentration of CAT and GSH compared to untreated ones that received distilled water only (Table 2). GSH also functions as free radical scavenger and in the repair of radical-caused biological damage (Meister, 1984). Lipid peroxidation is one of the characteristic features of chronic diabetes (Palsamy and Malathi, 2007). The extract induced significant (p<0.05) reduction in MDA values (lipid peroxidation) at 300 and 600 mg kg-1 compared to the control. Recent evidence indicates that some saturated and unsaturated aldehydes, including malondialdehyde (MDA), hydroxyaldehydes and other short chain carbonyl compounds contribute to peroxidative cell damage by reacting with sensitive biomolecules (Kim et al., 1999). STZ-diabetic animals exhibit most of the diabetic complications, namely, myocardial, nervous, vas deferens, gastrointestinal, kidney and urinary bladder dysfunction, through oxidative stress (Palsamy and Malathi, 2007). Diabetes generates reactive oxygen species which in turn cause lipid peroxidation and membrane damage (Hunt et al., 1988). ROS produce changes and have been implicated as a cause and consequence of a variety of pathological conditions including diabetes (Brzozowski et al., 1997).
In the DPPH assay, the extract produced a significant (p<0.05) antioxidant activity of 66% against 80% with ascorbic acid at 400 μg mL-1. The principle of FRAP method is based on the reduction of a ferric-tripyridyl triazine complex to its ferrous form in the presence of antioxidants. The FRAP values of the extract at 200 and 400 μg mL-1 were 1.42 and 1.68 μM, respectively while that of ascorbic acid was 2 μM (Fig. 2). This also revealed a high anti-oxidant ability of the crude extract.
The high antioxidant activities of the crude extract relative to the known antioxidant (ascorbic acid) attest to the popular use of preparations from the plant fruit in the local treatment of different health problems by people of diverse background.
In conclusion, this investigation revealed that the crude methanol extract of B. coriacea fruit was tolerated by rats and it possessed significant anti-diabetic, hypolipidemic and antioxidant activities. The observed effects were in support of the folkloric use of preparations from the plant fruit as remedy for broad spectrum of health challenges.
ACKNOWLEDGMENT
The authors sincerely thank Mr. Ozioko of Bioresources Development and Conservation Programme (BDCP) Nsukka, Enugu state, Nigeria for identifying the plant material.
REFERENCES
- Aebi, H., 1984. Catalase in vitro. In: Methods in Enzymology, Packer, L., Academic Press, Cambridge, Massachusetts, United States, ISBN: 9780121820053, pp: 121-126.
CrossRefDirect Link - Al-Enazi, M.M., 2007. Effect of thymoquinone on malformations and oxidative stress-induced diabetic mice. Pak. J. Biol. Sci., 10: 3115-3119.
CrossRefPubMedDirect Link - Al-Hader, A., M. Aqel and Z. Hasan, 1993. Hypoglycemic effects of the volatile oil of Nigella sativa seeds. Int. J. Pharmacol., 31: 96-100.
CrossRefDirect Link - Al-Omar, M.A., C. Beedham and I.A. Al-Sarra, 2004. Pathological roles of reactive oxygen species and their defence mechanisms. Saudi Pharm. J., 12: 1-18.
Direct Link - Amos, A.F., D.J. McCarty and P. Zimmet, 1997. The rising global burden of diabetes and its complications: Estimates and projections to the year 2010. Diabetic Med., 14: S7-S85.
PubMedDirect Link - Benzie, I.F.F. and J.J. Strain, 1999. Ferric reducing/antioxidant power assay: Direct measure of total antioxidant activity of biological fluids and modified version for simultaneous measurement of total antioxidant power and ascorbic acid concentration. Method Enzymol., 299: 15-27.
CrossRefPubMedDirect Link - Brzozowski, T., P.C. Konturek, Z. Sliwowski, D. Drozdowicz, E.G. Hahn and S.J. Konturek, 1997. Importance of nitric oxide and capsaicin-sensitive afferent nerves in healing of stress lesions induced by epidermal growth factor. J. Clin. Gastroenterol., 25: S28-S38.
PubMedDirect Link - Buege, J.A. and S.D. Aust, 1978. Microsomal Lipid, Peroxidation. In: Methods in Enzymology, Vol. 52, Flesicher, S. and L. Packer (Eds.)., Academic Press, New York, pp: 302-310.
CrossRefDirect Link - Chattopadhyay, R.R., 1993. Hypoglycemic effect of Ocimum sanctum leaf extract in normal and streptozotocin diabetic rats. Indian J. Exp. Biol., 31: 891-893.
PubMed - Dong, D.L. and G.W. Hart, 1994. Purification and characterization of an O-GlcNAc selective N-acetyl-beta-D-glucosaminidase from rat spleen cytosol. J. Biol. Chem., 269: 19321-19330.
PubMed - Ellman, G.L., 1959. Tissue sulfhydryl groups. Arch. Biochem. Biophys., 82: 70-77.
CrossRefPubMedDirect Link - Ene, A.C., E.A. Nwankwo and L.M. Samdi, 2008. Alloxan-induced diabetes in rats and the effects of black caraway (Carum carvi L.) oil on their body weights. J. Pharmacol. Toxicol., 3: 141-146.
CrossRefDirect Link - Ezeja, M.I., I.I. Ezeigbo and K.G. Madubuike, 2011. Analgesic activity of the methanolic seed extract of Buchholzia coriacea. Res. J. Pharmaceut. Biol. Chem. Sci., 2: 187-193.
Direct Link - Harris, M.I., W.C. Hadden, W.C. Knowler and P.H. Bennett, 1987. Prevalence of diabetes and impaired glucose tolerance and plasma glucose levels in U.S. population aged 20-74 yr. Diabetes, 36: 523-534.
PubMedDirect Link - Hunt, J.V., R.T. Dean and S.P. Wolff, 1988. Hydroxyl radical production and autoxidative glycosylation. Glucose autoxidation as the cause of protein damage in the experimental glycation model of diabetes mellitus and ageing. Biochem. J., 256: 205-212.
CrossRefPubMedDirect Link - Jasmine, R. and P. Daisy, 2007. Hypoglycemic and hypolipidemic activity of Eugenia jambolana in streptozotocin-diabetic rats. Asian J. Biochem., 2: 269-273.
CrossRefDirect Link - Jayatilake, G.S., H. Jayasuriya, E.S. Lee, N.M. Koonchanok and R.L. Geahlen et al., 1993. Kinase inhibitors from Polygonum cuspidatum. J. Nat. Prod., 56: 1805-1810.
PubMedDirect Link - Kaneto, H., G. Xu, K.H. Song, K. Suzuma, S. Bonner-Weir, A. Sharma and G.C. Weir, 2001. Activation of the hexosamine pathway leads to deterioration of pancreatic β-cell function through the induction of oxidative stress. J. Biol. Chem., 276: 31099-31104.
CrossRefDirect Link - Konrad, R.J., I. Mikolaenko, J.F. Tolar, K. Liu and J.E. Kudlow, 2001. The potential mechanism of the diabetogenic action of streptozotocin: inhibition of pancreatic beta-cell O-GlcNAc-selective N-acetyl-beta-D-glucosaminidase. Biochem. J., 356: 31-41.
PubMedDirect Link - Luo, B., G.J. Parker, R.C. Cooksey, Y. Soesanto, M. Evans, D. Jones and D.A. McClain, 2007. Chronic hexosamine flux stimulates fatty acid oxidation by activating AMP-activated protein kinase in adipocytes. J. Biol. Chem., 282: 7172-7180.
PubMed - Mahesh, N. and D. Brahatheeswaran, 2007. Anti-hyperglycemic activities of aqueous and ethanolic extracts Cynodon dactylon (Linn) streptozotocin-induced diabetic rats. Asian J. Biochem., 2: 66-72.
CrossRefDirect Link - Mensor, L.L., F.S. Menezes, G.G. Leitao, A.S. Reis, T.C. dos Santos, C.S. Coube and S.G. Leitao, 2001. Screening of Brazilian plant extracts for antioxidant activity by the use of DPPH free radical method. Phytother. Res., 15: 127-130.
CrossRefPubMedDirect Link - Priya, E.M., K.M. Gothandam and S. Karthikeyan, 2012. Antidiabetic activity of Feronia limonia and Artocarpus heterophyllus in streptozotocin induced diabetic rats. Am. J. Food Technol., 7: 43-49.
CrossRefDirect Link - Nirmala, A., J. Eliza, M. Rajalakshmi, E. Priya and P. Daisy, 2008. Effect of hexane extract of Cassia fistula barks on blood glucose and lipid profile in streptozotocin diabetic rats. Int. J. Pharmacol., 4: 292-296.
CrossRefDirect Link - Nweze, N.E. and I.U. Asuzu, 2006. Anthelminthic effect of Buchholzia coriacea seeds. Niger. Vet. J., 27: 60-65.
Direct Link - Okoli, B.J., O.S. Okere and S.O. Adeyemo, 2010. The antiplasmodial activity of Buchholzia coriacea. J. Med. Applied Biosci., 2: 21-29.
Direct Link - Palsamy, P. and R. Malathi, 2007. Evaluation of hypoglycemic and hypolipidemic activity of methanolic extract of Cocculus hirsutus (L.) diels leaves in streptozotocin-induced diabetes mellitus rats. Int. J. Biol. Chem., 1: 205-212.
CrossRefDirect Link - Pavana, P., S. Sethupathy and S. Manoharan, 2008. Protective role of Tephrosia purpurea ethanolic seed extract on glycoprotein components in streptozotocin induced diabetic rats. Int. J. Pharmacol., 4: 114-119.
CrossRefDirect Link - Sabo, C.E., S.R. Michael and L.L. Temple, 1999. The use of alternative therapies by diabetes educators. Diabetes Educ., 25: 945-956.
PubMedDirect Link - Soleimani, S., F.F. Azarbaizani and V. Nejati, 2007. The effect of Equisetum arvense L. (Equisetaceae) in histological changes of pancreatic β-cells in streptozotocin-induced diabetic in rats. Pak. J. Biol. Sci., 10: 4236-4240.
CrossRefPubMedDirect Link - Kim, S.S., D.D. Gallaher and A.S. Csallany, 1999. Lipophilic aldehydes and related carbonyl compounds in rat and human urine. Lipids, 34: 489-496.
CrossRefDirect Link - Subash-Babu, P. and S. Ignacimuthu, 2007. Antihyperlipidemic and antioxidant effect of hyponidd in the brain of streptozotocin induced diabetic rat. Int. J. Biol. Chem., 1: 196-204.
CrossRefDirect Link - Zimmermann, M., 1983. Ethical guidelines for investigations of experimental pain in conscious animals. Pain, 16: 109-110.
CrossRefPubMedDirect Link