
J. Patterson
Food Toxicology and Safety Laboratory, Nutritional Biochemistry Laboratory, Department of Food and Animal Sciences, Alabama A and M University, P.O. Box 1628, Normal, AL 35762, USA
J. Boateng
Food Toxicology and Safety Laboratory, Nutritional Biochemistry Laboratory, Department of Food and Animal Sciences, Alabama A and M University, P.O. Box 1628, Normal, AL 35762, USA
L.T. Walker
Department of Food and Animal Sciences, Alabama A and M University, P.O. Box 1628, Normal, AL 35762, USA
M. Verghese
Food Toxicology and Safety Laboratory, Nutritional Biochemistry Laboratory, Department of Food and Animal Sciences, Alabama A and M University, P.O. Box 1628, Normal, AL 35762, USA
Journal of Pharmacology and Toxicology
Year: 2012 | Volume: 7 | Issue: 3 | Page No.: 114-127







ABSTRACT
N-nitrosamines are considered human carcinogens and have been found in cured meats, seafood, vegetables, apples, beer, drinking water, waste water, tobacco products and rubber products. Limited studies exist on the effects of low dose exposure to multiple N-nitrosamines compounds. The objectives of this study were to investigate the cellular mechanisms of action by which N-nitrosamines exhibit toxicity resulting in liver tumors and other effects. Hep2G human liver cells (ATCC HTB-37) was obtained from the American Type Culture Collection (ATCC, Manassas, VA). For assay, 5x104 Hep2G cells/well were seeded in a 24 well culture plate and incubated at 37°C and 7% CO2 until development of a monolayer. Cells were incubated with a combination of selected N-nitrosamines at selected concentrations (0, 4, 8, 16, 32, 64 mM) for 12 and 24 h. Lactate dehydrogenase (LDH) release (% cytotoxicity), histone-related DNA fragmentation and detoxification enzymes were determined. After 12 and 24 h incubation with N-nitrosamines,% cytotoxicity in Hep2G cells displayed a dose-dependent relationship at concentrations of 4, 8 and 16 mM. Cytotoxicity peaked at 16 mM for both time periods and then decreased with increasing concentration (64 mM) to 19.46 (12 h) and 55.73 (24 h). Overall, levels of glutathione-S-transferase (GST), glutathione peroxidase (GPx), Glutathione Reductase (GR) and Superoxide Dismutase (SOD) were higher with control compared to N-nitrosamines-treated cells. Histone-related DNA fragmentation was highest in cells treated with 8 mM (24 h). Possible mechanisms of action may be due to lower detoxification enzymes and/or an increase in H2O2 production, leading to cell death.
PDF Abstract XML References Citation
Received: February 26, 2012;
Accepted: April 14, 2012;
Published: June 04, 2012
How to cite this article
J. Patterson, J. Boateng, L.T. Walker and M. Verghese, 2012. Cytotoxic Effects of Multiple N-nitrosamines in Human Liver Cell Line Hep2G: Possible Mechanisms of Action. Journal of Pharmacology and Toxicology, 7: 114-127.
DOI: 10.3923/jpt.2012.114.127
URL: https://scialert.net/abstract/?doi=jpt.2012.114.127
DOI: 10.3923/jpt.2012.114.127
URL: https://scialert.net/abstract/?doi=jpt.2012.114.127
INTRODUCTION
N-nitrosamines are a class of compounds formed endogenously or during food processing (Magee et al., 1976). They are a part of a larger group of compounds called N-nitroso compounds. N-nitrosamine structure consists of R1R2-N-N-O, where R1 and R2 are alkyl or acyl groups (Lijinsky, 1992; Rostkowska et al., 1998). Food products that have been associated with N-nitrosamine contamination have been divided into five major categories: foods preserved with addition of nitrates or nitrites, foods preserved by smoking (Yurchenko and Molder, 2006; Griesenbeck et al., 2009), foods dried by combustion gas, pickled or salt-preserved food and food stored or grown in high humid conditions (Griesenbeck et al., 2009). Preformed N-nitrosamines have been found in cured meats (Miller et al., 1989; Lijinsky, 1999), seafood (Montesano and Bartsch, 1976; Yamamoto et al., 1984; Song and Hu, 1988), vegetables (Atawodi et al., 1991; Mitacek et al., 1999), apples (Rudell et al., 2005), beer (Goff and Fine, 1979), drinking water (Levallois et al., 2000; Boyd et al., 2011; Ripolles et al., 2011), waste water (Krauss et al., 2009; Boyd et al., 2011), tobacco products (Jansson et al., 2003; Levy et al., 2006) and rubber products (Sen et al., 1989; Proksch, 2001).
More compounds have been tested and approximately 100 compounds in this class are carcinogenic in one or more experimental animals: rats, mice, hamsters, rabbits, minks, dogs, pigs and monkeys. Moreover, many N-nitrosamine compounds are reported to induce hepatocarcinogenesis (Manivasagam et al., 2006). The degree of tumor induction varies with species. Researchers infer that N-nitrosamines are equally carcinogenic in humans since the compounds are metabolized in a similar way in human and animal tissue (Magee et al., 1976).
N-nitrosamines become toxic when they are activated by biotransformation enzyme CYP2E1. Phase I hydroxylation and alkylation reactions yield nitrogen and a carbonium ion. The extremely reactive carbonium ion attacks DNA bases, causing methylation. Human studies using esophageal cells showed the formation of DNA adducts O6-methylguanine, 3-methylguanine and 7-methylguanine (Autrup et al., 1984). The formation of O6 methylguanine leads to a AT-GC transition mutation in the DNA sequence. This specific DNA adduct is most responsible for the mutagenecity of N-nitrosamines (Jagerstad and Skog, 2005). However, 7-methylguanine is used as a biomarker because it is not mutagenic and is present at 10 times the amount of O6-methylguanine and therefore easier to measure. Lack of DNA repair, constitutes the first stage of carcinogenesis called initiation (Sutandyo, 2010).
In addition to alkylation of DNA, evidence exists of CYP2E1 metabolism producing reactive oxygen species and free radicals (Bansal et al., 1996; Ogunlade et al., 2012). These compounds are associated with cancer, heart disease and degenerative diseases. Other cellular effects from N-nitrosamines exposure include apoptosis, enabling DNA repair and cell cycle blockage (Verghese et al., 2006).
Toxic effects of N-nitrosamines may cause cell death by either necrosis or apoptosis. Studies (Montesano and Bartsch, 1976; Autrup et al., 1984; Mitacek et al., 1999) report that N-nitrosamines cause necrosis of liver cells. However, other (Garcia et al., 2009) studies report apoptosis being the mechanism of cell death when exposed to single N-nitrosamines compounds, in various human cell lines including, Hep2G (Garcia et al., 2009), erythrocytes (Bansal et al., 1996), leukemia cells (Garcia et al., 2007), Caco2 (Robichova and Slamenova, 2002). Necrosis is characterized by cell swelling, leakage of cell membrane and blebbing. Apoptosis is characterized by the formation of apoptotic bodies or budding and elimination by phagocytosis (Khan, 2010).
Studies report multiple N-nitrosamine compounds occurring in food at low dose. However, limited studies exist on the effects of low doses exposure of a combination of N-nitrosamine in a cell culture model. Therefore , the aim of this study was to investigate the cellular mechanisms of action in which a combination of N-nitrosamine compounds exhibit toxicity in a cell culture model, including or (a) cytotoxicity, (b) detoxification and antioxidant enzymes, (c) DNA fragmentation and (d) morphological changes.
MATERIALS AND METHODS
General procedures: Hep2G human liver cells (ATCC HTB-37) was obtained from the American Type Culture Collection (ATCC, Manassas, VA). Cells were maintained in Dulbecco’s modified Eagles Media (DMEM) with 10% fetal bovine serum. For assay, 5x104 Hep2G cells/well were seeded in a 24 well culture plate and incubated at 37°C and 7% CO2 until development of a monolayer. After the development of a monolayer, 400 μL of fresh media (DMEM serum free) were added to the wells. N-nitrosamine mixtures (N-nitrosodimethylamine, N-nitrosopyrrolidine, N-nitrosodiethylamine, N-nitrosodipropylamine, N-nitrosodibutylamine, N-nitrosopiperidine, N-nitrosomethylethylamine) were made up to 100 μL with saline at various concentrations (4, 8, 16, 32, 64 mM), added and incubated for 12 and 24 h.
Determination of lactase dehydrogenase (LDH): The cytotoxic effects of N-nitrosamines were measured by the release of lactase dehydrogenase (LDH) from the Hep2G cells into the culture supernatant. The cells were incubated with the extracts for 12 and 24 h. A 10% triton-X solution was used as a positive control and the supernatant was collected. LDH was quantified by colorimetric cytotoxicity detection kit (LDH) (Roche Diagnostics, Indianapolis, IN) according to the manufacturer’s instructions. The absorbance was read at 490/600 nm.
Glutathione-S-transferase activity in cell lysates: Hep2G cells were incubated using N-nitrosamines for 12 and 24 h. The supernatant was collected and centrifuged at 10,000 xg for 10 min in PBS buffer (pH 7) and used for GST assay using a microplate reader (BioTek Synergy HT) and kinetic absorbances were read at 340 nm at 5 min intervals.
Determination of catalase activity: Cell catalase activity was determined using a microplate reader (BioTek Synergy HT) at 240 nm as described by Aebi (1984). The reaction mixture (1 mL) contained 0.02 mL suitably diluted cytosol in phosphate buffer (50 mM, pH 7.0) and 0.1 mL of 30 mM H2O2 in phosphate buffer. The specific activity of catalase was expressed as moles of H2O2 reduced per min per mg protein.
Determination of superoxide dismutase (SOD), glutathione peroxidase (GPx), glutathione reductase (GR) activity: Superoxide dismutase, glutathione, glutathione peroxidase and glutathione reductase enzymes were estimated using assay kits according to manufacturer’s instructions (Cayman Chemicals, Ann Arbor, MI).
Protein determination: Protein was determined according to manufacturer’s instructions using a bicinchoninic acid (BCA) protein assay kit (Thermo Scientific, Rockford, IL).
DNA fragmentation: To determine the effect of N-nitrosamines in induction of apoptosis, 1x104 Hep2G cells were treated with N-nitrosamines for 12 and 24 h. After treatment, cells were washed with PBS and cell lysates were prepared using cell lysis buffer and centrifuged at 200 xg for 10 min. Apoptosis was determined by measuring internucleosomal DNA fragmentation by using a cell death detection ELISA kit (Cell Death Detection ELISA, Roche Molecular Biochemicals, Indianapolis, IN) according to the instructions provided by the manufacturer. Spectrophotometric analysis was carried out using an ELISA plate reader at 405 nm for the histone related DNA fragmentation.
Statistical analysis: Statistical analysis was conducted using SAS 9.1, 2004 (SAS Institute Inc., Cary, NC) using analysis of the variance (One-Way ANOVA) to determine any significant differences among the groups. A separate analysis was used for each time period. Results were presented as Means±Standard Error Mean (SEM). Where significant, means were separated using Tukey’s studentized range test. The level of significance was set at p<0.05.
RESULTS
Effect of N-nitrosamines on cytotoxicity in Hep2G cells: Percent cytotoxicity (LDH release) compared to the control is shown in Fig. 1. After 12 h incubation,% cytotoxicity displayed a dose-dependent relationship at concentrations of 4, 8 and 16 mM. At these concentrations, LDH release (%) ranged from 15.66 to 66.74 (12 h). The% cytotoxicity peaked at 16 mM for both time periods and then decreased at the highest concentration (64 mM) to 19.46 (12 h) and 55.73 (24 h).
Overall, the greatest cytotoxicity (%) was seen at 16 mM-24 h (98.32) which was significantly (p≤0.05) higher compared to all other treatments except 32 mM-24 h (90.20). However, % cytotoxicity at 4 mM-24 h, 8 mM-24 h, 16 mM-12 h (concentration-time) were similar. Furthermore, cells incubated with 8 mM (12 h) and 32 mM (12 h); 32 mM (12 h) and 64 mM (24 h) and 4 mM (12 h) and 64 mM (12 h) had similar cytotoxicity as shown by the release of LDH. In cells treated with N-nitrosoamines, incubation with 4 mM (12 h) and 64 mM (24 h) resulted in the lowest cytotoxicity. There were no significant differences in cytotoxicity (%) at the lower concentrations (4 and 8 mM) in cells incubated for 24 h. This trend was also observed in cells treated with 16 and 32 mM for a 24 h period.
There were significant (p≤0.05) differences between incubation times of 12 and 24 h at each individual concentration which indicated a time-dependent effect. At 4 mM, the% cytotoxicity at 24 h was almost 4-fold higher than that seen at 12 h. The% cytotoxicity at 24 h for 8, 16, 32 and 64 mM was over 1.5-fold higher than the % cytotoxicity at 12 h, at the respective concentrations.
After 12 h incubation, % cytotoxicity at 16 mM was significantly (p≤0.05) higher compared to all other treatments at the same incubation period. However, there were no significant differences in LDH release among 8 mM (45.48%) and 32 mM (48.50%) and 4 mM (15.67%) and 64 mM (19.46%) at the 12 h time period. After 24 h incubation, the highest LDH release (%) was seen at N-nitrosamine concentration of 16 mM which was significantly (p≤0.05) higher compared to 4, 8 and 64 mM (55.73-72.56). There were no significant differences in % cytotoxicity, between 4 and 8 mM at 24 h and 16 mM at 12 h. The lowest % cytotoxicity at 24 h was seen with incubation of N-nitrosamines at 64 mM (55.73%).
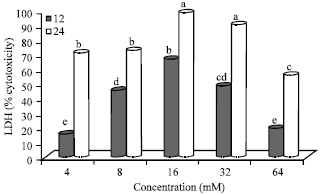 | |
| Fig. 1: | Percent cytotoxicity of N-nitrosamine-treated Hep2G cells, Bars means with same letter are not significantly different by Tukey’s studentized range test (p≤0.05) |
Effect of N-nitrosamines on glutathione-S-transferase activity: Glutathione-S-transferase activity (nmol/mg protein) (GST) in N-nitrosamine-treated Hep2G cells is shown in Fig. 2. Overall, the control (untreated cells) had significantly (p≤0.05) higher GST activity at both time periods (12 and 24 h). GST activity (nmol/mg protein) ranged from to a low of 1.12 (64 mM-12 h) to a high of 8.21 (control-24 h). After 12 h incubation, GST activity decreased in the treated cells (3.81-1.12). Cells treated (12 h) with 4, 8, 16, 32 and 64 mM N-nitrosamines had a 53, 68, 80, 83 and 86% lower GST activity compared to the control (untreated cells). At the 24 h incubation time, there were no differences in GST activity for cells treated with N-nitrosamines (4-64 mM) (1.77-2.09). Cells treated (24 h) with 4, 8, 16, 32 and 64 mM had a 74, 76, 67, 78 and 77% lower GST activity, respectively, compared to the control. The GST activity was higher after 12 h incubation in cells treated with 4 and 8 mM compared to 24 h. However, in cells treated with 16, 32 and 64 mM N-nitrosamines, GST activity was higher at 24 h compared to 12 h incubation. The GST activity was 45 and 23% higher at 12 h compared to 24 h in cells treated with N-nitrosamines at low concentrations (4 and 8 mM). However, at concentrations of 16, 32 and 64 mM, GST activity was 41, 24 and 42% lower at 12 h compared to 24 h incubation.
Effect of N-nitrosamines on glutathione reductase (GR) activity: Glutathione Reductase (GR) activity in cells treated with N-nitrosoamines for 12 and 24 h is shown in Fig. 3. GR activity (nmol/mg protein) ranged from a low of 7.11 (32 mM-24 h) to a high of 16.67 (Control, 24 h).
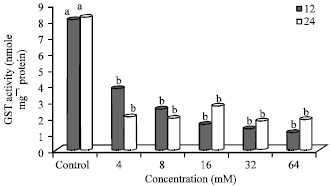 | |
| Fig. 2: | Glutathione-S-transferase activity in N-nitrosamine-treated Hep2G cells, Bars means with same letter are not significantly different by Tukey’s studentized range test (p≤0.05) |
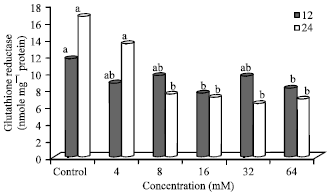 | |
| Fig. 3: | Glutathione reductase activity in N-nitrosamine-treated Hep2G cells, Bars means with same letter are not significantly different by Tukey’s studentized range test (p≤0.05) |
The control cells-24 h (16.67) had a significantly (p≤0.05) higher GR activity compared to 8 mM (24 h), 16 mM (12 and 24 h), 32 mM (24 h) and 64 mM (12 and 24 h).
At 12 h incubation, treatment with N-nitrosamines at 4, 8, 16, 32 and 64 mM resulted in a 24, 17, 35, 17 and 30% lower GR activity compared to the control, with values ranging from 7.63-9.68. In addition, the GR activity of the control (11.66) was significantly (p≤0.05) higher compared to the cells treated with 16 and 64 mM (7.63-8.18) for the same time period (12 h).
After 24 h incubation, GR activity decreased, leveling off at 8 mM. Treatment with 4, 8, 16, 32 and 64 mM of N-nitrosamines resulted in a 19, 55, 57, 62 and 58% lower GR activity compared to the control.
The GR activity in the control (untreated) and 4 mM N-nitrosamine treated cells were higher at 24 h compared to 12 h incubation. However, in cells treated with 8, 16, 32 and 64 mM, GR activity was higher at 12 h compared to 24 h (with 23, 7, 33 and 15% reductions).
Effect of N-nitrosamines on glutathione peroxidase (GPx) activity: Glutathione peroxidase (GPx) activity in cells treated with N-nitrosamines (12 and 24 h) is shown in Fig. 4. GPx (nmol/mg protein) activities (12.23-22.68) were similar to the control except in cells treated with 16 mM N-nitrosamines (24 h). After 12 h incubation, GPx activity was highest in the control (22.17) and lowest at 32 mM (13.77) N-nitrosamine concentration. At 24 h, the control (22.68) had the highest, while cells treated with 16 mM (12.23) N-nitrosamines had the lowest GPx activity.
The GPx activity was lower at 12 h compared to 24 h incubation for untreated (control) cells and cells treated with 4 and 32 mM N-nitrosamines. At higher concentrations (8-64 mM), except 32 mM, GPx activity was higher at 12 h compared to 24 h incubation.
Effect of N-nitrosamines on superoxide dismutase (SOD) activity: Superoxide dismutase (SOD) activity is shown in Fig. 5. SOD activity (U mg-1 protein) was dose-dependent in control, 4, 8 and 16 mM treated cells and ranged from 6.68 to 10.20 for 12 h and 5.23 to 7.92 for 24 h. At 32 mM concentration (12 h), SOD activity was similar to the control. After 12 h, treatment with N-nitrosamines at 4, 8 and 16 mM resulted in 20, 29 and 35% lower SOD activity, respectively compared to the control. After 24 h incubation, values decreased by 11, 25 and 34% for 4, 8 and 16 mM treated cells, respectively, compared to the control (untreated cells). Cells treated with 8 mM (5.91) and 16 mM (5.23) at 24 h had significantly (p≤0.05) lower SOD activity compared to selected control (12 h) and 32 mM (12 h).
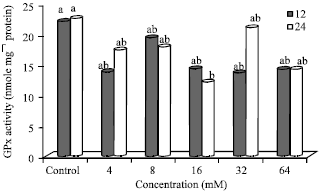 | |
| Fig. 4: | Glutathione peroxidase activity in N-nitrosamine-treated Hep2G cells, Bars means with same letter are not significantly different by Tukey’s studentized range test (p≤0.05) |
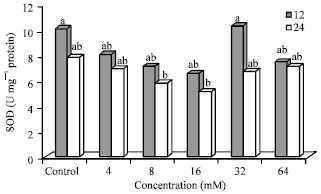 | |
| Fig. 5: | Superoxide dismutase activity in N-nitrosamine-treated Hep2G cells, Bars means with same letter are not significantly different by Tukey’s studentized range test (p≤0.05) |
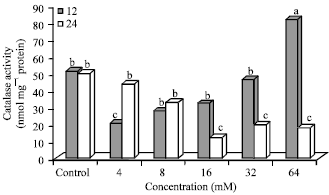 | |
| Fig. 6: | Catalase activity in N-nitrosamine-treated Hep2G cells, Bars means with same letter are not significantly different by Tukey’s studentized range test (p≤0.05) |
SOD activity was lower in all cells (treated and untreated) after 24 h incubation compared to 12 h. Among the treated cells (4-64 mM), those treated with 32 mM at 12 h had the highest SOD activity. SOD activity was 14, 18, 21, 35 and 5% lower at 24 h compared to 12 h in cells treated with N-nitrosamines (4-64 mM).
Effect of N-nitrosamines on catalase (CAT) activity: CAT activity in control and treated cells is shown in Fig. 6. Cells treated with 64 mM (12 h) (81.85) had significantly (p≤0.05) higher CAT activity compared to other treatment groups (12.55-51.36). After 12 h incubation, enzyme activity was significantly (p≤0.05) lower (by 59%) in 4 mM treated cells compared to the control. At higher concentrations (16-64 mM), CAT activity (32.66-81.86) was significantly (p≤0.05) higher at 12 h compared to 24 h incubation. However, CAT activity was not significantly different in cells treated with 8, 16 and 32 mM (12 h). After 24 h incubation, CAT activity for 4, 8, 16, 32 and 64 mM treated cells was 12, 34, 75, 60 and 64% lower, respectively compared to the control. CAT activity in cells treated with 4 and 8 mM N-nitrosamines was higher at 24 h compared to incubation at 12 h. At higher concentrations (16-64 mM), CAT activity was significantly higher at 12 h compared to 24 h incubation.
CAT activity was 62, 57 and 77% lower at 24 h compared to 12 h in cells treated with N-nitrosamines at higher concentrations (16-64 mM). However, at 4 and 8 mM, CAT activity was over 2-fold and 17% higher at 24 h compared to 12 h incubation.
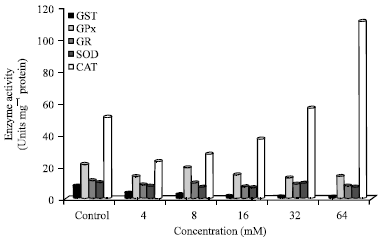 | |
| Fig. 7a: | Trend analysis of enzyme activities after 12 h incubation. GST: Glutathione-S-transferase, GR: Glutathione reductase, GPx: Glutathione peroxidase, SOD: Superoxide dismutase, CAT: Catalase |
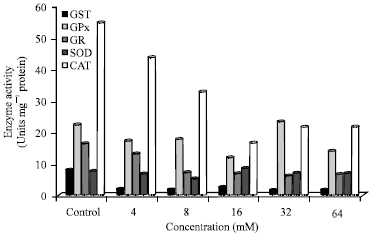 | |
| Fig. 7b: | Trend analysis of enzyme activities after 24 h incubation. GST: Glutathione-S-transferase, GR: Glutathione reductase, GPx: Glutathione peroxidase, SOD: Superoxide dismutase, CAT: Catalase |
Comparison of enzyme activity after 12 h and 24 h incubation: Trend analysis of enzyme activities after 12 h incubation is shown in Fig. 7a. Comparing all the enzyme activities, CAT activity was highest in control and N-nitrosamine treated cells (12 h). There seemed to be a dose response increase in CAT activity with a steady increase seen in 4-64 mM treated cells. GPx activity was higher compared to GR in control and N-nitrosamine treated cells. Furthermore, SOD activity was lower than CAT activity in control and N-nitrosamine treated cells.
Trend analysis of enzyme activities after 24 h incubation is shown in Fig. 7b. CAT activity was again higher compared to other enzyme activities in control and treated cells at all concentrations except 32 mM. There was a dose response decrease seen in cells treated with 4, 8 and 16 mM N-nitrosamines compared to untreated cells. GPx activity was higher than GR activity in control and N-nitrosamine treated cells. The GPx and GR activities were lower at 4, 8, 16 and 64 mM compared to the control (untreated cells).
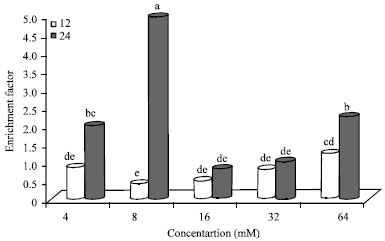 | |
| Fig. 8: | Effect of N-nitrosamines on histone-related DNA fragmentation in Hep2G cells, Bars means with same letter are not significantly different by Tukey’s studentized range test (p≤0.05) |
However, GPx activity was higher at 32 mM while GR had no significant change at the same concentrations. SOD activity was lower than CAT activity in control and N-nitrosamine treated cells.
Histone-related DNA fragmentation in Hep2G cells: Histone-related DNA fragmentation (enrichment factor) with respect to the control is shown in Fig. 8. Overall, cells incubated with 8 mM (24 h) N-nitrosamines had significantly (p≤0.05) higher enrichment factor (4.97) compared to all other N-nitrosamine treated cells (0.41-2.24). The enrichment factor at lower doses (4-8 mM) was also higher than those at selected higher doses (16 and 32 mM). At all concentrations, cells incubated for 24 h had higher enrichment factors compared to those incubated for 12 h. Enrichment factors for N-nitrosamine treated cells at 24 h (4-64 mM) were over 2-, 12-, 1.6-, 1.2- and 1.8-fold, respectively higher compared to the enrichment factors at 12 h.
At 12 h incubation, the enrichment factor showed a steady increase in cells treated with N-nitrosamines (8-64 mM) with values ranging from 0.41 to 1.39. Cells treated with 16 and 32 mM N-nitrosamines had similar enrichment factors at both 12 and 24 h incubation periods.
After 24 h incubation, enrichment factor ranged from 0.82 to 4.97. Cells treated with 64 mM (24 h) and 4 mM (24 h) of N-nitrosamines had similar enrichment factors. The enrichment factor was higher at 24 h incubation compared to 12 h incubation for all cells treated with N-nitrosamines at selected concentrations (4-64 mM). The enrichment factors were 2-, 1.2-, 1.6-, 1.2- and 1.8-fold higher at 24 h compared to 12 h at the selected concentrations (4-64 mM).
DISCUSSION
N-nitrosamines have shown to be potent hepatotoxins mainly due to their activation by CYP 2E1 metabolizing enzymes in the liver. This study evaluated the effects of treatment of N-nitrosamines (N-nitrodimethylamine, N-nitrosodiethylamine, N-nitrosodipropylamine, N-nitrosodibutylamine, N-nitrosopiperidine, N-nitrosopyrrolidine) in a human liver cell line Hep2G. Effects studied include cytotoxicity (LDH release), detoxification and antioxidative enzymes, apoptosis by histone-related DNA fragmentation (apoptosis) and morphological changes.
Percent cytotoxicity was determined by measuring LDH release. LDH is an enzyme present in nearly all cells and is released from the cytosol of damaged cells (Shirazi et al., 2006; Verghese et al., 2006). Results showed the highest LDH release at 16 mM at both 12 and 24 h incubation, followed by a decline. This decrease in release of LDH indicated an increase in the number of dead cells since LDH is only released from cells can be explained by the decline in detoxification and antioxidative enzyme activity. Jeyachandran et al. (2007) reported a 42% increase in alkaline phosphatase release after exposure to nitrosodiethylamine, compared to the control.
Glutathione-S-transferase, glutathione reductase and glutathione peroxidase are classes of glutathione-linked enzymes that play a role in eliminating active molecules, thus decreasing toxicity and toxic effects (Williams et al., 2008). Glutathione-S-transferase is a Phase II detoxification enzyme that conjugates electrophillic molecules with sulfyhydryl groups using glutathione as a substrate (Liska et al., 2006). Glutathione reductase catalyzes the reduction of glutathione disulfide (GSSH) to GSH which is a cellular antioxidant. In turn GSH is used in the reduction of lipid peroxides and H2O2 to water (Comporti, 1987; Bansal et al., 1996). This reduction is catalyzed by glutathione peroxidase.
The two antioxidative enzymes studied were superoxide dismutase (SOD) and catalase (CAT). SOD converts O2-to H2O2 (Gajula et al., 2010) which is then decomposed to H2O by catalase (Fridovich, 1975; Bansal et al., 1996).
Enzyme activity (GST, GR, SOD, CAT) showed a decrease, indicating inhibition activities. Some in vivo and in vitro studies have reported N-nitrosamines inducing detoxification and antioxidative enzymes (Sheweita and Mostafa, 1995; Bansal et al., 1996). These studies test the effect of single N-nitrosamine compounds (i.e., NDMA, NDEA) on the enzyme systems. In our study, Hep2G cells were treated with a combination of seven N-nitrosamine compounds. Thus, a dose-dependent decrease in enzyme activity was probably due to competitive inhibition which occurs when there are many compounds completing for several detoxification enzymes, causing an increase in toxic load (Liska et al., 2006). Our enzyme data correlates with the LDH release data in that the highest % cytotoxicity and lowest detoxification enzyme activities were seen at 16 mM.
CAT activity at 12 h incubation, increased dose-dependently. This trend implies that there was an increase in H2O2 production and CAT was responsible for its decomposition into water (Bansal et al., 1996). Studies have found H2O2 to be a metabolite of N-nitrosamine biotransformation (Kushida et al., 2009; Hebels et al., 2010) which would further explain the dose-dependent increase in CAT activity after 12 h incubation (Fig. 6). GPx converts H2O2 to decrease toxicity; however, the reaction takes place only when GSH is available as a cofactor (Sheweita and Mostafa, 1996). A dose-dependent decrease with other glutathione-linked enzymes implies that GSH may be depleted with increasing concentrations. Similar depletion of glutathione occurred with increasing dose exposure to heavy metals (El-Enany, 2000). Therefore, after 12 h incubation, a decrease in H2O2 in the cell was due to the action of CAT.
CAT and SOD work together to decrease toxic effect of compounds (Fridovich, 1975; Bansal et al., 1996). A slight decrease in SOD activity after 12 h incubation (4-16 mM) is an indication of limited amount of O2-production. However, higher SOD activity at 32 mM indicates an increase in H2O2 which resulted in a higher CAT activity also seen at 64 mM (12 h).
Internucleosomal degradation (ladder) of genomic DNA is a classic characteristic of apoptosis (Arends et al., 1990; Corcoran et al., 1994; Hockenbery, 1995; Darzynkiewicz et al., 1997; Dhanasekaran and Ganapathy, 2011). Our results (Fig. 8) indicate highest DNA fragmentation occurring in cells treated with lower doses (4-8 mM) of N-nitrosamines. In contrast, other studies report no DNA cleavage at lower doses of NDMA (1 mM) (Lin et al., 1999), NPIP, NPYR and NDMA (0.1-4 mM) (Garcia et al., 2007, 2009). A high enrichment factor at low concentrations is probably due to an additive effect of exposure to the combination of N-nitrosamine compounds (Bartsch et al., 1989).
DNA fragmentation at higher doses (16-64 mM) was lower than that at lower doses (4-8 mM). This implies that necrosis is a plausible mechanism at higher concentrations of exposure. Arranz et al. (2007) reported NDMA, NPIP, NDBA and NPYR inducing apoptosis in both Hep2G and HL 60 cell lines when treated singly (Arranz et al., 2007). In our study, at higher concentrations, cell death is probably due to additive or synergistic effects causing increased cell damage, leading to death by necrosis. It is possible that apoptosis is occurring at higher concentration even in the absence of DNA fragmentation. Studies have shown that morphological changes characteristic of apoptosis occur without an indication of internucleosomal DNA fragmentation (Cohen et al., 1992; Collins et al., 1992; Lin et al., 1999).
CONCLUSION
In conclusion, this study has shown that treating Hep2G cells with multiple N-nitrosamines causes cell damage and affects the detoxification mechanisms of the cell. These characteristics may be due to lower detoxification enzymes and an increase in oxidative stress (H2O2 production), leading to cell death by apoptosis (at low doses) or necrosis (at high doses).
ACKNOWLEDGMENT
This project was funded by The Alabama Agricultural Experimental Research Station, Normal AL 35762.
REFERENCES
- Arends, M.J., R.G. Moris and A.H. Wyllie, 1990. Apoptosis: The role of the endonuclease. Am. J. Pathol., 136: 593-608.
PubMed - Arranz, N., A.I. Haza, A. Garcia, J. Rafter and P. Morales, 2007. Protective effect of vitamin C towards N-nitrosamineinduced DNA damage in the single-cell gel electrophoresis (SCGE)/HepG2 assay. Toxicol. In vitro, 21: 1311-1317.
CrossRef - Atawodi, S.E., E.N. Maduagwu, R. Preussmann and B. Spiegelhalder, 1991. Potential of endogenous formation of volatile nitrosamines from nigerian vegetables and spices. Cancer Lett., 57: 219-222.
CrossRef - Autrup, H., C.C. Harris, W. Sao-Ming, B. Lo-Yao and P. Xu-Fang et al., 1984. Activation of chemical carcinogens by cultured human fetal liver, esophagus and stomach. Chem. Biol. Interact., 50: 15-25.
CrossRef - Bansal, A.K., D. Bhatnagar and G.L. Soni, 1996. In vitro effect of n-nitrosodiethylamine on lipid peroxidation and antioxidant system in human erythrocytes. Toxicol. In vitro, 10: 649-653.
CrossRefPubMedDirect Link - Bartsch, H., E. Hietanen and C. Malaveille, 1989. Carcinogenic nitrosamines: Free radical aspects of their action. Free Radical Biol. Med., 7: 637-644.
PubMedDirect Link - Boyd, J.M., S.E. Hrudey and X.F. Li, 2011. Solid-phase extraction and high-performance liquid chromatography mass spectrometry analysis of nitrosamines in treated drinking water and wastewater. TrAC Trends Anal. Chem., 30: 1410-1421.
CrossRef - Cohen, G.M., X.M. Sun, R.T. Snowden, D. Dinsdale and D.N. Skilleter, 1992. Key morphological features of apoptosis may occur in the absence of internucleosomal DNA fragmentation. Biochem. J., 286: 331-334.
PubMed - Collins, R.J., B.V. Harmon, G.C. Gobe and J.F. Kerr, 1992. Internucleosomal DNA cleavage should not be the sole criterion for identifying apoptosis. Int. J. Radiat. Biol., 61: 451-453.
PubMed - Comporti, M., 1987. Glutathione depleting agents and lipid peroxidation. Chem. Phys. Lipids, 45: 143-169.
Direct Link - Corcoran, G.B., L. Fix, D.P. Jones, M.T. Moslen, P. Nicotera, F.A. Oberhammer and R. Buttyan, 1994. Apoptosis: Molecular control point in toxicity. Toxicol. Applied Pharmacol., 128: 169-181.
Direct Link - Dhanasekaran, J.J. and M. Ganapathy, 2011. Hepatoprotective effect of Cassia auriculata L. leaf extract on carbon tetrachloride intoxicated liver damage in wister albino rats. Asian J. Biochem., 6: 104-112.
CrossRefDirect Link - Darzynkiewicz, Z., G. Juan, X. Li, W. Gorczyca, T. Murakami and F. Traganos, 1997. Cytometry in cell necrobiology: Analysis of apoptosis and accidental cell death (necrosis). Cytometry, 27: 1-20.
PubMed - El-Enany, A.A., 2000. Phytochelatins as a biomarker for cu-toxicity in soybean callus cultures. Pak. J. Biol. Sci., 3: 799-803.
CrossRefDirect Link - Proksch E., 2001. Review toxicological evaluation of nitrosamines in condoms. Int. J. Hyg. Environ. Health, 204: 103-110.
PubMedDirect Link - Garcia, A., P. Morales, N. Arranz, E. Delgado and A.I. Haza, 2007. Induction of apoptosis and reactive oxygen species production by n-nitrosopiperidine and n-nitrosodibutylamine in human leukemia cells. Toxicol. Lett., 172: S66-S66.
CrossRef - Garcia, A., P. Morales, J. Rafter and A.I. Haza, 2009. N-nitrosopiperidine and n-nitrosodibutylamine induce apoptosis in hepg2 cells via the caspase dependent pathway. Cell Biol. Int., 33: 1280-1286.
PubMedDirect Link - Gajula, D., M. Verghese, J. Boateng, L. Shackelford and S.R. Mentreddy et al., 2010. Basil (Ocimum basilicum and Ocimum tenuiflorum) reduces azoxymethane induced colon tumors in fisher 344 male rats. Res. J. Phytochem., 4: 136-145.
CrossRefDirect Link - Goff, E.U. and D.H. Fine, 1979. Analysis of volatile n-nitrosamines in alcoholic beverages. Food Cosmet. Toxicol., 17: 569-573.
Direct Link - Griesenbeck, J.S., M.D. Steck, J.C. Huber, J.R. Sharkey, A.A. Rene and J.D. Brender, 2009. Development of estimates of dietary nitrates, nitrites, and nitrosamines for use with the short willet food frequency questionnaire. Nutr. J., Vol. 8.
CrossRef - Hebels, D.G., J.J. Briede, R. Khampang, J.C. Kleinjans and T.M. de Kok, 2010. Radical mechanisms in nitrosamine- and nitrosamide-induced whole-genome gene expression modulations in caco-2 cells. Toxicolog. Sci.: Off. J. Soc. Toxicol., 116: 194-205.
PubMedDirect Link - Jagerstad, M. and K. Skog, 2005. Genotoxicity of heat-processed foods. Mutat. Res./Fundam. Mol. Mech. Mutagen., 574: 156-172.
CrossRefPubMedDirect Link - Jansson, C., A. Paccou and B.G. Osterdahl, 2003. Analysis of tobacco-specific n-nitrosamines in snuff by ethyl acetate extraction and liquid chromatography-tandem mass spectrometry. J. Chromatogr. A, 1008: 135-143.
PubMedDirect Link - Jeyachandran, R., A. Mahesh and L. Cindrella, 2007. DEN-Induced cancer and its alleviation by Anisomeles malabarica (L.) R.Br. ethanolic leaf extract in male albino mice. Int. J. Cancer Res., 3: 174-179.
CrossRefDirect Link - Khan, A.A., 2010. Intracellular mechanisms of apoptosis. J. Biol. Sci., 10: 291-305.
CrossRefDirect Link - Krauss, M., P. Longree, F. Dorusch, C. Ort and J. Hollender, 2009. Occurrence and removal of N-nitrosamines in wastewater treatment plants. Water Res., 43: 4381-4391.
PubMedDirect Link - Kushida, H., K.I. Fujita, A. Suzuki, M. Yamada, T. Endo, T. Nohmi and T. Kamataki, 2009. Metabolic activation of N-alkylnitrosamines in genetically engineered Salmonella typhimurium expressing cyp2e1 or cyp2a6 together with human nadph-cytochrome p450 reductase. Carcinogenesis, 21: 1227-1232.
CrossRefDirect Link - Levallois, P., P. Ayotte, J.M. van Maanen, T. Desrosiers and S. Gingras et al., 2000. Excretion of volatile nitrosamines in a rural population in relation to food and drinking water consumption. Food Chem. Toxicol., 38: 1013-1019.
PubMedDirect Link - Levy, D.T., E.A. Mumford, K.M. Cummings, E.A. Gilpin and G.A. Giovino et al., 2006. The potential impact of a low-nitrosamine smokeless tobacco product on cigarette smoking in the united states: Estimates of a panel of experts. Addictive Behaviors, 31: 1190-1200.
PubMedDirect Link - Lijinsky, W., 1999. N-Nitroso compounds in the diet. Mutation Res. Genet. Toxicol. Environ. Mutagenesis, 443: 129-138.
CrossRef - Lin, H.L., L.A. Parsels, J. Maybaum and P.F. Hollenberg, 1999. N-nitrosodimethylamine-mediated cytotoxicity in a cell line expressing p450 2e1: Evidence for apoptotic cell death. Toxicol Applied Pharm., 157: 117-124.
PubMedDirect Link - Liska, D., M. Lyon and D.S. Jones, 2006. Detoxification and biotransformational imbalances. Explore: J. Sci. Healing, 2: 122-140.
CrossRefDirect Link - Manivasagam, T., S. Perumal, E. Musthafa Mohamed, S.S. Ganapathy Suthakar and S. Ramar, 2006. Effects of s-allyl cysteine on n-nitrosodiethylamine induced hepatocarcinogenesis: variations in temporal patterns of circulatory tumor marker enzymes. Int. J. Pharmacol., 2: 136-141.
CrossRefDirect Link - Miller, B.J., S.M. Billedeau and D.W. Miller, 1989. Formation of n-nitrosamines in microwaved versus skillet-fried bacon containing nitrite. Food Chem. Toxicol., 27: 295-299.
PubMed - Mitacek, E.J., K.D. Brunnemann, M. Suttajit, N. Martin, T. Limsila, H. Ohshima and L.S. Caplan, 1999. Exposure to N-nitroso compounds in a population of high liver cancer regions in Thailand: Volatile nitrosamine (vna) levels in thai food. Food Chem. Toxicol., 37: 297-305.
PubMedDirect Link - Montesano, R. and H. Bartsch, 1976. Mutagenic and carcinogenic n-nitroso compounds: Possible environmental hazards. Mutation Res. Rev. Genet. Toxicol., 32: 179-227.
CrossRef - Ripolles, C., E. Pitarch, J.V. Sancho, F.J. Lopez and F. Hernandez, 2011. Determination of eight nitrosamines in water at the ngl−1 levels by liquid chromatography coupled to atmospheric pressure chemical ionization tandem mass spectrometry. Anal. Chim. Acta, 702: 62-71.
CrossRef - Robichova, S. and D. Slamenova, 2002. Effects of vitamins C and E on cytotoxicity induced by n-nitroso compounds, n-nitrosomorpholine and n-methyl-n'-nitro-n-nitrosoguanidine in caco-2 and v79 cell lines. Cancer Lett., 182: 11-18.
PubMedDirect Link - Rostkowska, K., K. Zweirz, A. Rozanki, J. Moniuszko-Jakoniuk and A. Roszczenko, 1998. Formation and metabolism of N-nitrosamines. Polish J. Environ. Stud., 7: 321-325.
Direct Link - Rudell, D.R., J.P. Mattheis and J.K. Fellman, 2005. Evaluation of diphenylamine derivatives in apple peel using gradient reversed-phase liquid chromatography with ultraviolet-visible absorption and atmospheric pressure chemical ionization mass selective detection. J. Chromatogr. A, 1081: 202-209.
Direct Link - Sen, N.P., S.W. Seaman and S.C. Kushwaha, 1989. Determination of non-volatile n-nitrosamines in baby bottle rubber nipples and pacifiers by high-performance liquid chromatography-thermal energy analysis. J. Chromatogr. A, 463: 419-428.
Direct Link - Sheweita, S. and M. Mostafa, 1995. Recovery of the hepatic carcinogen-metabolizing capacity in schistosome-infected mice after treatment with the antischistosomal praziquantel. Oncol. Rep., 1: 155-159.
PubMedDirect Link - Sheweita, S.A. and M.H. Mostafa, 1996. N-nitrosamines and their effects on the level of glutathione, glutathione reductase and glutathione s-transferase activities in the liver of male mice. Cancer Lett., 99: 29-34.
PubMedDirect Link - Shirazi, S.H., H. Yazdanpanah, F. Khoshjoo and L. Hosseinzadeh, 2006. Pistachio extracts effects on the aflatoxin B1 cytotoxicity in HepG2 cells. Int. J. Pharmacol., 2: 233-239.
CrossRefDirect Link - Song, P.J. and J.F. Hu, 1988. N-nitrosamines in chinese foods. Food Chem. Toxicol., 26: 205-208.
CrossRefPubMedDirect Link - Verghese, M., D.R. Rao, C.B. Chawan, L.T. Walker and L. Shackelford, 2006. Anticarcinogenic effect of phytic acid (IP6): Apoptosis as a possible mechanism of action. LWT-Food Sci. Technol., 39: 1093-1098.
CrossRefDirect Link - Williams, D.S., M. Verghese, L.T. Walker, J. Boateng and L.A. Shackelford et al., 2008. Chemopreventive effects of flax seed oil and flax seed meal on azoxymethane-induced colon tumors in fisher 344 male rats. Int. J. Cancer Res., 4: 28-40.
CrossRefDirect Link - Yamamoto, M., R. Iwata, H. Ishiwata, T. Yamada and A. Tanimura, 1984. Determination of volatile nitrosamine levels in foods and estimation of their daily intake in japan. Food Chem. Toxicol., 22: 61-64.
CrossRef - Yurchenko, S. and U. Molder, 2006. Volatile N-nitrosamines in various fish products. Food Chem., 96: 325-333.
CrossRefDirect Link - Aebi, H., 1984. Catalase in vitro. In: Methods in Enzymology, Packer, L., Academic Press, Cambridge, Massachusetts, United States, ISBN: 9780121820053, pp: 121-126.
CrossRefDirect Link - Ogunlade, B., L.C. Saalu, O.S. Ogunmodede, G.G. Akunna, O.A. Adeeyo and G.O. Ajayi, 2012. The salutary role of Allium cepa extract on the liver histology, liver oxidative status and liver marker enzymes of rabbits submitted to alcohol-induced toxicity. Am. J. Biochem. Mol. Biol., 2: 67-81.
CrossRefDirect Link
sunita Reply
good