
W.E. Gato
Department of Chemistry and Biochemistry, Southern Illinois University, IL 62901, Carbondale
J.C. Means
Department of Chemistry and Biochemistry, Southern Illinois University, IL 62901, Carbondale
Journal of Pharmacology and Toxicology
Year: 2011 | Volume: 6 | Issue: 3 | Page No.: 234-248







ABSTRACT
The mutagen 2-aminoanthracene (2-AA) is an aromatic amine or arylamine, which belongs to a class of polycyclic aromatic hydrocarbons. 2-AA is used in the manufacturing of a wide variety of chemicals, drugs, dyes and polymers. Non-occupational sources include tobacco smoking and cooked foods. The goal of this study is to evaluate pancreatic gene expression patterns in Fischer-344 (F-344) male rats exposed to 2-AA. As a first step the effects of 2-aminoanthracene exposure on the pancreas with particular interest in genes that relate to insulin and insulin metabolism. To achieve this goal, twenty-four post-weaning, 3-4 week old F-344 male rats were exposed to 0 mg kg-1-diet (control), 50 mg kg-1-diet (low dose), 75 mg kg-1-diet (medium dose) and 100 mg kg-1-diet (high dose) 2-AA for 14 and 28 days followed by analysis of the pancreas for broad gene expression changes. Results obtained from our study suggest most of the mRNA transcripts that were differentially expressed are involved in energy metabolism in the pancreas, protein digestion and some that play an active role in pancreatitis and pancreatic cancer. Some of these genes include: insulin, colipase pancreatic, carboxypeptidase, chymotrypsinogen B1 and chymotrypsin C (caldecrin). These findings seem to point to the role of 2-AA in the dysregulation of several pancreatic genes that regulate lipid and protein metabolism in a way that involves a feedback mechanism which may ultimately lead to insulin resistance and tissue autolysis. Quantitative PCR determination of fold changes in selected genes show similar trends to global expression determined via microarray analyses.
PDF Abstract XML References Citation
Received: June 14, 2010;
Accepted: August 16, 2010;
Published: November 12, 2010
How to cite this article
W.E. Gato and J.C. Means, 2011. Pancreatic Gene Expression Altered Following Dietary Exposure to 2-Aminoanthracene: Links to Diabetogenic Activity. Journal of Pharmacology and Toxicology, 6: 234-248.
DOI: 10.3923/jpt.2011.234.248
URL: https://scialert.net/abstract/?doi=jpt.2011.234.248
DOI: 10.3923/jpt.2011.234.248
URL: https://scialert.net/abstract/?doi=jpt.2011.234.248
INTRODUCTION
Cigarette smoking is one of the most important preventable causes of disease and death around the world. It is estimated that more than 2.4 million deaths annually are attributable to cigarette smoke. Cigarette smoking is strongly associated with all kinds of diseases including cancers, heart disease and diabetes. It is associated with cancers such as lung, bladder, pancreatic and esophageal cancer. Pancreatic cancer is the fourth or fifth leading cause of cancer-related death and its mortality rate is at 98% in the US, hence understanding its etiology and as well developing biomarkers will be crucial in overcoming it. When a cigarette is combusted, more than 4000 chemicals are generated in the process. Some of these are large free radicals; alkyl, alkoxyl, peroxyl and quinine/hydroxyquinone, Polycyclic Aromatic Hydrocarbons (PAH) (benzo[a]pyrene (BaP), N-nitrosamine, aldehydes, nicotine, arylamines and nitric oxide (Cerami et al., 1997; Wang and Wang, 2005; Soleimani et al., 2007; Chipitsyna et al., 2009; Tseng, 2009).
Similarly, cigarette smoking has been associated with insulin sensitivity and insulin absorption in type 2 diabetes. Reports have shown insulin resistance development and changes in markers of insulin resistance syndrome due to cigarette smoke. Diabetes affects about 194 million people worldwide with estimations in the US between 16-17 million. This figure is projected to reach 300 million by the year 2025. In the US, lost productivity due to diabetes was estimated at $120 billion in 2000. It was $60 billion in 1992. Type 2 diabetes mellitus affects about 7% of adults in the US. Insulin-dependent diabetes mellitus which results from specific disruption of the pancreatic islet insulin-secreting beta cells is induced in experimental animals by toxic agents including chemicals that are byproducts of cigarette smoking (Eliasson et al., 1997; Bott et al., 2005; Boudreau et al., 2006; Elayat et al., 1995; Wang et al., 2007).
Arylamines are known mutagens and carcinogens that occur both naturally and synthetic form. They are employed in the manufacture of dyes, drugs, inks, rubber antioxidants, plastics and agricultural chemicals. These aromatic amines are also used as curing agents in synthesizing epoxy resins and polyurethanes and are found in road tars and synthetic fuels. 2-amino anthracene is the benchmark aromatic amine used for toxicity studies. It is a model aryl amine because relatively, 2-amino anthracene is potent direct-acting carcinogen and induces mutations in both eukaryotic and prokaryotic cells (Boudreau et al., 2006; Snyderwine et al., 1992; Zhu et al., 1995).
Previous research in this laboratory examined the effect of 2-AA on growth, tissue histological, immunocytochemical and clinical pathological end points in the pancreas. Cytological, immunocytochemical and histological results demonstrate alterations in the endocrine and exocrine pancreas cellular morphology. Following these findings, further research was conducted to determine the extent of hepatic toxicity of 2-AA (the primary site for metabolic activation of aromatic amines) as well as to determine whether the cessation of 2-AA exposure reduces or eliminates clinical, pathologic and histologic effects reported. After 5 weeks, clinical chemistry returned to normal while histopathologic lesions in the liver were common suggesting sub-cellular changes (such as DNA mutations) might have occurred that did not become phenotypically evident until hepatocytes had undergone several replication cycles. Although results from this laboratory and other researchers have led to the inference that there is subcellular and molecular injury due to 2-AA exposure, this has yet to be demonstrated directly by global gene expression alterations (Boudreau et al., 2006; Baker et al., 2001).
It is therefore the goal of this study to evaluate pancreatic gene expression patterns in Fischer 344 male rats. As a first step, this research examines the effects of sub-chronic 2-aminoanthracene dietary exposure on the pancreas global gene expression flowed by focused quantitative gene expression studies with particular emphasis upon genes that relate to insulin biosynthesis and insulin metabolism and organ system function.
MATERIALS AND METHODS
Experimental design: Twenty-four post-weaning, 3-4 week old Fisher 344 male rats were purchased from Harlan Laboratories and randomly assigned to four dose regimens of 0 mg kg-1-diet, 50 mg kg-1-diet, 75 mg kg-1-diet and 100 mg kg-1-diet 2-aminoanthracene (2-AA) for 14 and 28 days. There were three animals per group. Rats were individually housed at the Southern Illinois University Animal Facility and acclimated in an AAALAC certified animal facility for two weeks prior to exposure. This experiment was performed between 2009 and 2010, rats were provided distilled water ad libitum. The animals were treated according to the principles outlined in the NIH and Southern Illinois University Guide for the Care and Use of Laboratory Animals. The temperature was held at 20±1°C and a 12/12 h light/dark cycle was maintained in the exposure room. Animals were weighed and food consumption measured and recorded. At the end of each exposure period (14 or 28 days), rats were euthanized with CO2 and blood was collected by cardiac puncture. Livers and kidneys were excised and snap frozen in liquid nitrogen. Excised pancreas tissue was stored in RNALater stabilization solution until total RNA was subsequently extracted from the pancreas.
Diet preparation: 2-AA (CAS# 613-13-8) [98+% Pure] was obtained from Sigma-Aldrich (St. Louis, MO) and used without further purification. A kilogram of rat diet supplied by PMI Nutrition International, LLC (Brentwood, MO) was immersed in 1 L molecular grade ethyl alcohol containing the mass of 2-AA necessary to yield the target dose concentrations in the diets and the ethyl alcohol evaporated under the hood with periodic thorough mixing to assure homogeneity. The diet was stored in the freezer and protected from light until given to the animals.
Total RNA extraction: Total RNA was isolated from rat pancreas using a Qiagen RNA Isolation kit (Qiagen, 2006 Valencia, CA. Approximately 30 mg pancreas samples) were homogenized in RLT buffer. RLT buffer denatures and inactivates RNases. The RNA is then allowed to bind to a silica-gel membrane and finally eluted with RNase-free water. Total RNA was quantified using the UV-VIS spectrophotometer at 260 and 280 nm absorbance and control electrophoretic gels were run for RNA quality assurance purposes to examine purity and the absence of genomic DNA in the samples.
Microarray gene expression analysis: Total RNA extracts were prepared for microarray analysis using the manufacturer’s protocol (Affymetrix, 2008/9). Double stranded cDNA was synthesized from total RNA samples using reverse transcriptase and oligo dT primers. The synthesized cDNA served as a template for an in vitro transcription (IVT) reaction in which amplified RNA or aRNA is synthesized and involves the incorporation of biotin-conjugated nucleotide to produce cRNA molecules. These cRNA molecules were purified to remove NTPs, salts, enzymes and inorganic phosphate and then fragmented using array fragmentation buffer and hybridized to GeneChip® Rat 230 2.0 Array and subsequently scanned as described in Affymetrix GeneChip™ Wash, Stain and Scan protocol (Affymetrix, 2008/9). The Rat 230 2.0 chip contains over 31,000 probe sets analyzing more than 30,000 transcripts and 28,000 variants representing more than 28,000.
Data analysis: Significant differences in body weight gain and organ weight during the treatment period were analyzed by ANOVA and student t-test using EXCEL and were expressed as mean±SE.
Affymetrix GeneChip Command Console software (AGCC) controlled the fluidic station and the scanner. Intensity data was generated from the image data (.dat) acquired from the scanner. Using the Affymetrix expression console, single intensity values was computed for every probe locus on the arrays from the image data (.dat files) and saved as .cel files. Also, quality control data was generated using the plier algorithm in the expression console. The fluorescence intensity due to proper hybridization of each target gene to the chip was estimated by examining the difference in fluorescence intensities in perfect match and mismatch probe pairs present at each locus on the chip. These intensities were scaled for all valid probes using a default target signal threshold.
The .cel files were imported into Biometric Research Branch-Array Tools (BRB-ArrayTools) software (Simon and Lam, 2005) for data collation, filtering, normalization and gene sub-setting. Genes which passed through the above quality assurance process were then analyzed for differential expression by scatter-plot of phenotype averages. Data collation involved importing data and aligning genes. BRB-ArrayTools converts either EXCEL or CHP files into a tab-delimited format. Individual arrays were filtered using spot filters. Spots on arrays which had signal intensities less than 10 were considered below a threshold and not analyzed further. A log base 2 transformation was applied to all data and each array data set was normalized to a reference array such that log-intensity differences between any experimental array and the reference array equaled zero over all the genes on the array. The reference was chosen to be the median array. Minimum gene fold-change filter was used to exclude probe sets from all arrays which did not meet the following criteria: the minimum fold change less than 20% of expression data values at least a 1.5-fold change in either direction from the gene’s median value (array that has median log-intensity value to be the median over all the median log-intensity values for the complete set of arrays). Over 600 genes met these criteria and thus were used in our subsequent analysis of differential gene expression and hierarchical clustering.
Quantitative real-time polymerase chain reaction (qRT-PCR): To validate gene expression data obtained from microarray experiments, quantitative real-time polymerase chain reaction (qRT-PCR) was employed to quantify the specific mRNA expression changes of six selected genes and a control gene β-Actin. Using the National Center for Biotechnology Information (NCBI) database, the FASTA mRNA sequence of β-Actin, Pnlip, Cpa2, Ins1, Ctrc, Cel and Clps were obtained for Rattus norvegicus. These sequences were employed in the PrimerQuest Custom Design Tool to design primers (Table 1) that were ordered from the Integrated DNA Technologies Inc. (IDT, Coralville, IA). The primers were combined with iScript one-step RT-PCR kit with SYBR green (Bio-Rad, 2005) and the quantitative PCR product determined on a Chromo4 System for real-time PCR detection (Bio-Rad, 2010).
Fluorescent intensity values were analyzed using relative expression software tool (REST©) which were implemented in Excel (Pfaffl et al., 2002).
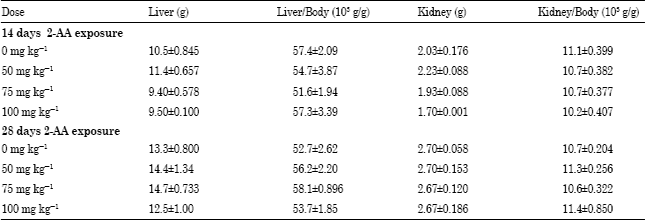
As presented in Table 2, liver and kidney weights and their corresponding organosomatic indices were compared between treated and untreated 2-AA rats. Organ weights as well as hepatosomatic and renal somatic indices were not significantly altered either for the 14 or 28-day treatment groups, though these indices are slightly less for the medium and high dose groups.
| Table 1: | qRT-PCR primer pair sequence and related product size |
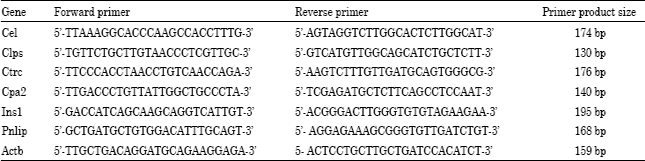 | |
| Table 2: | Selected organ weights (g) and their indices (organ wt. x 103/b.wt.) in response to 2-AA exposure. The data points represent the averages of 3 male F-344 rats fed 0, 50, 75 and 100 mg kg-1 2-AA diet for 14 and 28 days |
 | |
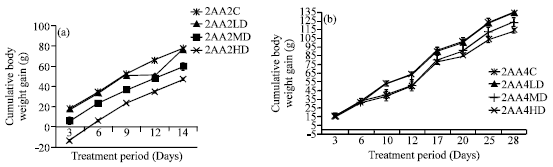 | |
| Fig. 1: | The effects of 2-AA on F-344 body weight. Each data point is the mean of 3 F-344 male rats given untreated (0 mg kg-1-diet) and treated (50, 75 and 100 mg kg-1-diet) 2-AA diet. Body weight gain is significantly reduced at p<0.01 for both exposure time points in the medium and high dose treatments when compared with the controls |
In contrast, cumulative body weight gain was significantly reduced at the medium (75 mg kg-1-diet) and high (100 mg kg-1-diet) dose groups relative to control animals for both exposure time points as shown in Fig. 1a and b. For the two-week study, body weights were decreased 39 and 23%, respectively in the high and medium dose relative to controls. Similarly, the high and medium dose body weights were reduced 15 and 8% correspondingly with respect to the controls. For the low dose (50 mg kg-1-diet) exposure diet group, no significant effect on the body weights of animals was detected in either treatment exposure period group.
Global gene expression changes: In order to examine the effects of 2-AA on pancreatic tissues with particular interest in insulin regulation, global gene expression levels in the pancreas were determined in each exposure and control group. Several quality control metrics derived from Affymetrix Expression Console that help to ensure data integrity and validity. For the current study, each of these quality control parameters was found to be within normal operating ranges and did not warrant exclusion of any array or repeating any assays.
Figure 2, 3 and Table 3 show the effects of 2-AA on pancreatic gene expression. Figure 3 and 4 represent all the genes that were at least 2-fold differentially expressed for the 14 and 28 day treatment periods, respectively. In both instances, arrays from treated animals were compared with the corresponding controls in the A, B and C panels. In the case of the D and E panels, the medium and high dose transcripts were compared with the low dose responses.
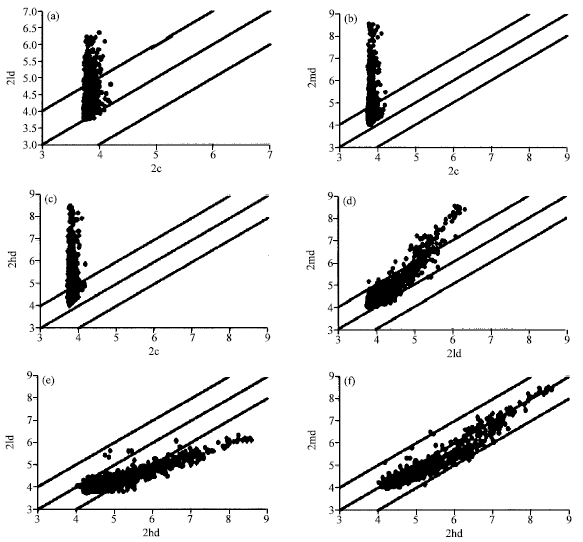 | |
| Fig. 2: | Scatterplots showing differentially expressed genes due to 2-AA exposure at two-fold change for 14 days time frame. (a-c) represent comparison between 50, 75 and 100 mg kg-1 2-AA and the control respectively, whereas, (d, e) were medium and high dose 2-AA compared with low dose and finally f was the scatterplot between medium and high dose 2-AA. There were three arrays in each phenotype class |
Finally, panel F represents a comparison between the medium and high dose treatment regimens. The points that were equal to or greater than two-fold up- or down-regulated genes were those falling outside of the pair of marker lines as demonstrated in Fig. 2 and 3. Table 3 lists the number of genes that were >2-fold, >3-fold and >5-fold up- or down-regulated for each dose and exposure period group. More genes were found to be either up-regulated in the 14-day group than in the 28-day group as compared with their controls. In contrast, for the 28-day exposure group, pronounced down-regulation in gene expression was observed for the low and medium dose groups when related to their controls than between the control and high dose animals. In general, a dose-dependent increase in the number of up-regulated genes was observed at the 14 days exposure period relative to controls but not at 28 days, while an inverse dose-dependence in down-regulated genes was observed in the 28 d exposure group relative to controls but not in the 14 days exposure group.
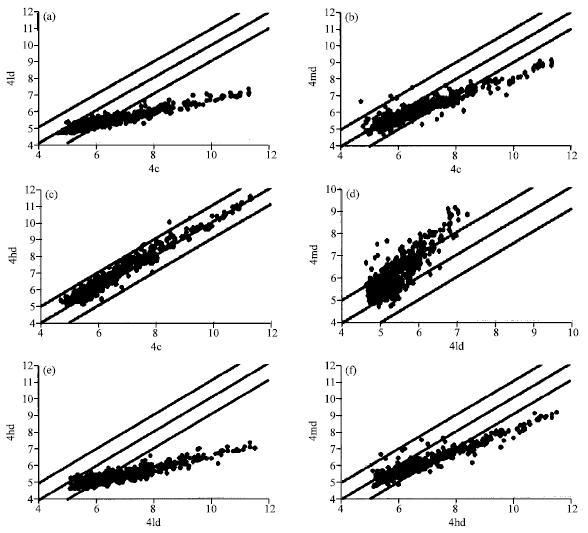 | |
| Fig. 3: | (a-f) Two-fold gene expression changes in the pancreas of F-344 rats exposed to 2-AA through diet for 28 days. (a-c) compares low, medium and high dose to the control while, compared medium and high to low dose and medium to high dose, respectively |
| Table 3: | The number of mRNA transcripts whose expression levels were either suppressed or enhanced as a result of 2-AA intoxication that are represented by C (0 mg kg-1), LD (50 mg kg-1), MD (75 mg kg-1) and HD (100 mg kg-1). Transcript numbers were extracted from Figures 3 and 4 at 2-, 3- and 5-fold differential expression. There were three arrays per group |
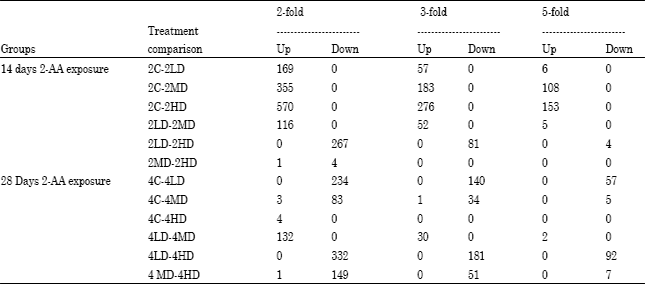 | |
Figure 2, scatterplots showing differentially expressed genes due to 2-AA exposure at two fold change for 14 days time frame. (a-c) represent comparison between 50 mg kg-1, 75 mg kg-1 and 100 mg kg-1 2-AA and the control respectively, whereas, (d, e) were medium and high dose 2-AA compared with low dose and finally f was the scatterplot between medium and high dose 2-AA. There were three arrays in each phenotype class.
Figure 3, two-fold gene expression changes in the pancreas of F-344 rats exposed to 2-AA through diet for 28 days. a, b and c compares low, medium and high dose to the control while d, E compared medium and high to low dose and medium to high dose respectively.
A select set of genes that were at least 5-fold differentially altered are presented in Table 4-6 for different dose/time groups and Table 7 presents a list of gene differentially altered by >10-fold. Many of these genes are related to energy metabolism in the pancreas, while others such as chymotrypsinogen C are documented to be involved in pancreatitis, which is an inflammation of the pancreas. Among the genes differentially expresses are those that control hydrolysis of fat molecules such as acylglycerols, phospholipids and ceramide. Others genes such as carboxypeptidase and chymotrypsinogen B1 are involved in protein digestion.
Microarray data were validated by a series of quantitative polymerase chain reaction assays for select genes. The selected transcripts reflect the observation of genes that are important in energy metabolism in the pancreas, inflammation or pancreatitis, or in control of lipid metabolism.
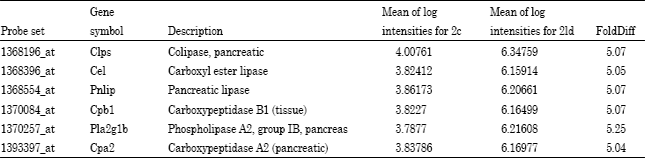
| Table 4: | Genes that were either up-regulated by 5-fold comparing control and low dose for the 14 day exposure group |
 | |
| Table 5: | Genes that were down-regulated by 5-fold comparing control and medium dose groups at the 28-day treatment period |
 | |
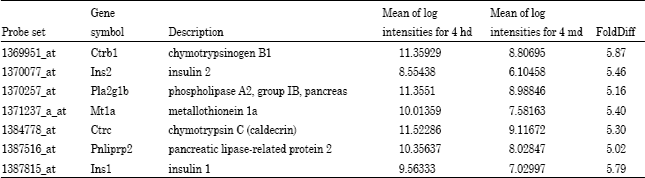
| Table 6: | Genes that were down-regulated by 5-fold comparing high dose and medium dose at the 28-day treatment period |
 | |
| Table 7: | A ten-fold change of gene expression comparing control and low dose treatment groups for 28 days |
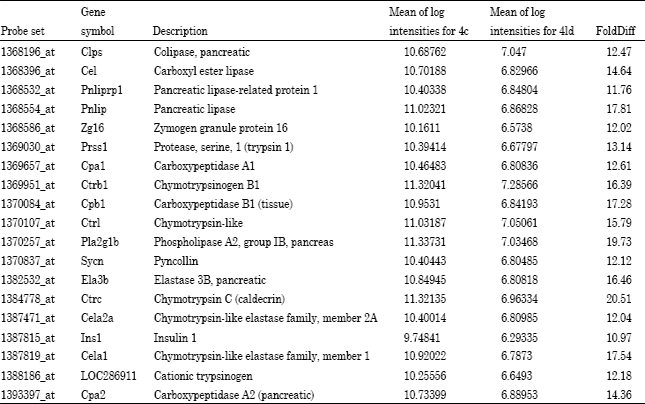 | |
| Table 8: | Gene expression of selected genes determined by quantitative real-time PCR. These fold changes were calculated relative to β-actin and paired control sample |
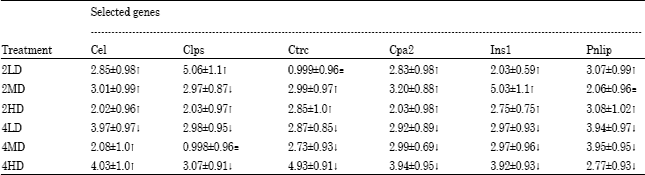 | |
The genes chosen for mRNA transcript validation included: Pnlip (pancreatic lipase), Cpa2 (Carboxypeptidase A2 [pancreatic]), Ins1 (Insulin 1), Ctrc (Chymotrypsin C [caldecrin]), Cel (Carboxyl ester lipase) and Clps (Colipase, pancreatic) and β-Act (beta Actin) as shown in Table 1. Although the quantitative fold differences observed from the quantitative PCR data were smaller when compared with the fold difference values obtained from the microarray data, they nevertheless show the same differential expression trends. The results are presented in Table 8. Most of the two-week study genes were up-regulated when compared with their control whereas in contrast, the four-week study genes were down-regulated when compared with the controls. A full listing of all genes that were up- or down regulated in the microarray data sets in this study is available as supplementary data posted at http://opensiuc.lib.siu.edu
DISCUSSION
The current study investigated the gene expression responses in the pancreas of F-344 male rats after dietary exposure to the suspected diabetogenic agent 2-AA. Body weight gain results were consistent with previous study from this research group. Also no significant hepatic or renal tissue weight reductions or alterations in the somatic indices for these organs (Table 2) were observed.
The primary focus of this study was to elucidate the potential molecular basis for the role of 2-AA in causing diabetes and potentially pancreatic cancer using both global gene expression analysis and qPCR of pancreas tissues in F-344 male rats. As a first step, we analyzed genes that were differentially expressed by at least 5-fold level up or down relative to controls in order to find any unique transcripts that relate to tumor initiation and/or the onset of diabetes. Results obtained from our study suggest most of the RNA transcripts that meet these criteria were divided into groups of genes primarily involved in energy metabolism in the pancreas, protein digestion and some that were reported to play an active role in inflammatory responses associated with either pancreatitis or pancreatic cancer. These are discussed in greater detail below. Gene ontology, clustering and pathway analyses will be published as a separate follow-up study to the current one.
Trends in global gene expression: The major observation in global changes in gene expression patterns observed in this study were the presence of an apparent dose response in the number of genes over-expressed in the 14 day exposure group and the dramatic shift to an inverse dose response in the number of genes suppressed in the 28 day exposure. These trends held true for all genes that were up- or down-regulated by >2-fold, >3-fold and >5-fold in this experiment (Table 3). This is consistent with a stress response during a short-term exposure followed by a pattern suggesting that gene expression is first suppressed at low doses and an adaptive recovery of gene expression at increasing doses over a longer period of exposure.
Among the genes discovered that were up-regulated by >5-fold by the low dose during the 14 day exposure were the six genes: Clps, Cel, Pnlip, Ppb1, Pla2g1b and Cpa2 (Table 4). These lipases and proteases each have a role in the exocrine function of the pancreas. Boudreau et al. (2006) noted the 2-AA caused histologically evident damage to the exocrine cells of the pancreas with longer-term exposures. These results may suggest that damage is occurring in these cells during initial exposure and that gene expression is being up-regulated to compensate for this damage.
A much larger array of genes were down-regulated by >5-fold in animals exposed at the low dose for 28 days (Table 7). Among them are all of those which were up-regulated in the short-term exposure group. In addition to these lipases and proteases and a few new ones such as pancreatic lipase-related protein1 [Pnliprp1], zymogen granule protein 16 [Zg 16], protease serine 1 [Prss1], carboxypeptidase A1, B1 [Ppa1 and Cpb 1], chymotrypsin-like [Ctrl], chymotrypsin C or caldecrin [Ctrc], several structural genes associated with zymogen granules like pancreatic elastin 3B [Ela3B] and syncollin [Sycn] and finally insulin1 [Ins1] were affected by the longer exposure to the 2-AA low dose (25 mg kg-1-diet). Elastin 3B is used as a clinical indicator of overall pancreatic exocrine function.
Whitcomb and Lowe, 2007 noted that the pancreas is primary organ responsible for producing digestive enzymes that are then transported to the small intestine for the hydrolysis of complex nutrients. Some of the mRNA transcripts that were found to relate to the metabolism of sugars and lipids include: insulin, colipase pancreatic, carboxyl ester lipase and phospholipases. Phospholipases are known by their unique ability to hydrolyze sn-2 ester bond of the phospholipid substrate to yield free fatty acids and lysophospholipids and are also known to influence inflammatory responses such as recruitment of neutrophils and macrophages. The process by which this happens leads to the production of essential second messengers that play vital physiological roles (Burke and Dennis, 2009; Funk, 2001). One of the by-products of phospholipase action is the release of free arachidonic acid, an eicosanoid, which involved in prostaglandin biosynthesis and is therefore very important mediator and regulator of many physiological and pathophysiological states (Perez-Chacon et al., 2009; Chilton et al., 1996). Carboxyl ester lipase (Cel) previously called pancreatic cholesterol esterase (bile salt-dependent lipase) is a non-specific lipolytic enzyme that can catalyze the hydrolysis of cholesteroyl esters, tri, di and mono-acylglycerols, phospholipids, lysophospholipids and ceramides (Hui and Howles, 2002; Hui, 1996). This transcript is also reported to participate in chylomicron assembly and secretion. In cultured cells, Cel was found to mediate lipoprotein metabolism and oxidized LDL-induced atherosclerosis (Hui and Howles, 2002). Similarly, colipase and its protein cofactor Pancreatic Triglyceride Lipase (PTL) work together to efficiently digest dietary triglycerides (Lowe, 2002).
Insulin is one of the primary protein hormones that regulates blood glucose levels (Bansal and Wang, 2008). The importance of this hormone relates to its ability to regulate cellular energy supply and macronutrient balance as well as direct anabolic processes in the fed state. Insulin is required for intra-cellular transport of glucose into insulin-dependent tissues including muscle and adipose tissues (Wilcox, 2005; Bevan, 2001). Insulin is produced in the β-cells of the pancreatic islets of the Langerhans (Wilcox, 2005). Binding of insulin to insulin receptors initiates its action. This involves insulin binding to the extracellular á subunit of the insulin receptor which leads to conformational change that enables ATP to bind to the intracellular component of the of β subunit of the receptor. The ATP-binding results in tyrosine phosphorylation that activates various signaling pathways such phosphatidylinosytol 3-kinase/Akt signaling and mitogen-activated protein kinase activation. These pathways are coordinated in such a way as to regulate glucose transport, protein and lipid biosynthesis and mitogenic processes (Wilcox, 2005; Kido et al., 2001; Wolever, 1990).
The major source of proteases required for digestion of ingested proteins are the cells of the exocrine pancreas. These proteins typically are bio-synthesized in their inactive or pro-enzyme forms (zymogens) including trypsin, chymotrypsinogen A and B, proelastase, procarboxypeptidase A1, A2, B1, B2 and others (Whitcomb and Lowe, 2007). Some of the other mRNA transcripts found to be involved in protein digestion from our study include: carboxypeptidase A2, carboxypeptidase B1, chymotrypsin-like cationic trypsinogen and chymotrypsin C (caldecrin). The chymotrypsin-related proteins observed to be modulated by 2-AA administration in this study are primarily serine proteases (Hedstrom, 1996; Ma et al., 2005). Chymotrypsin acts preferentially to cleave dietary proteins at aromatic amino acid substrate residues such as phenylalanine, tyrosine and tryptophan (Ma et al., 2005). The catalytic mechanism of action involves chymotrypsin binding to the substrate to form a Michaelis complex, which is nucleophilically attacked by Ser 195 that result in the formation of tetrahedral intermediate. This intermediate then decomposes to acyl-enzyme intermediate that is quickly deacylated to regenerate the enzyme and a carboxylated product (Voet and Voet, 2004). Carboxypeptidases are known to catalyze the hydrolysis of C-terminal amino acid residues of polypeptides. These enzymes are highly specific for the various chemical reactions they catalyze (Voet and Voet, 2004). Carboxypeptidase B is documented to play a vital role in the conversion of pro-insulin into insulin. Also, it is reported to aid in the processing of various insulin precursors (Zuhlke et al., 1976). Thus, down-regulation of Cpb could lead to a reduced conversion of pro-insulin to insulin leading to dysregulation of glucose blood levels as observed in an earlier study (Boudreau et al., 2006).
Various studies have shown that some of the mRNA transcripts reported in our study to play significant role in either pancreatitis or pancreatic cancer (Witt and Bhatia, 2008; Chen et al., 2007a, b; Al-Bahrani and Ammori, 2005; Chen et al., 2007b). Pancreatitis is an inflammation of the pancreas. Recent studies by Chen et al. (2007a,b), reported that chymotrypsinogen b proteins were present in both pancreatic cancer juice and pancreatitis juice in proteomic analysis of inflamed tissue. In both cases, this protein was up-regulated. Witt and Bhatia (2008) noted that genetic changes in the loci of genes coding for trypsinogens and the trypsin-degrading transcript chymotrypsin C was associated with the prevalence of idiopathic chronic pancreatitis in the Western world.
The current study was undertaken to better understand the role of 2-AA in the induction of type 2 or insulin-dependent diabetes and potentially pancreatic cancer. There is strong evidence to suggest an association between diabetes and pancreatic cancer (Chari, 2007). Although pancreatic cancer is observed to cause glucose intolerance, the diabetes induced due to pancreatic cancer is of short duration and does not persist. Boudreau et al. (2006) published histopathological data that demonstrated that there was a severe dose/time-related disruption of the cellular architecture of pancreas at higher doses of 2-AA. This disruption was characterized pathologically as necrosis. The disruption was evident in both the endocrine (β cells) and exocrine (acinar cells) pancreas. They also reported the absence of a baseline expression response of the oncogene ras while a slight increase in the oncogene c-myc was observed in the pancreas tissues. In the present study, no activation of the oncogenes ras or c-myc was observed. Further, a suite of genes typically associated with the initiation of apoptosis (controlled cell death), as contrasted with necrosis, was also not found to be affected by 2-AA administration. This confirms the previous findings that necrosis of the pancreas tissues appears to be occurring and may be the results of up-regulation of a broad spectrum of genes which yield proteases, lipases and other digestive enzymes causing auto-digestion of the pancreas leading to symptomology of diabetes and uncontrolled necrosis of the pancreas tissues. Genes associated with tumor cell proliferation such as PCNA and p53 were also not affected in the current study. Cytokines typically associated with an inflammatory response were also not modulated in the present experiments.
It was postulated more than one hundred years ago that pancreatitis may be the result of pancreatic auto-digestion. Recent research seems to validate this theory (Witt and Bhatia, 2008; Nemoda and Sahin-Toth, 2006; Chem and Ferec, 2009). Nemoda and Sahin-Toth (2006) observed that chymotrypsin C employs a novel positive feedback mechanism in its auto-activation of human cationic trypsinogen, thereby facilitating the trypsinogen enzyme cascade. Active trypsin is capable of activating trypsinogen (auto-activation) and other pancreatic protease zymogens such as chymotrypsin C. When activated chymotrypsin C catalyzes the conversion of N-terminus of still un-activated trypsinogens and as a result enhances auto-activation of the most abundant cationic isoform. Chem and Ferec, 2009 support the observation that auto-activation and premature activation of trypsin in the pancreas may play a central role in the etiology of pancreatitis. The premature-activated trypsin, if not inhibited, could initiate auto-digestion within the pancreas and thus trigger the activation cascade of digestion zymogens which include chymotrypsin, elastase, kallikrein, carboxypeptidase A, carboxypeptidase B, phospholipase A2 and colipase (Chem and Ferec, 2009). Expressions of all the genes associated with these enzymes except kallikrein were found in our study to be differentially altered.
Gene expression data was validated by analyzing quantitative PCR of mRNA transcripts found to be involved in processes such as energy metabolism in the pancreas, protein digestion and others noted to play an active role in inflammatory responses associated with both pancreatitis and pancreatic cancer. The selected genes include pancreatic lipase (Pnlip), carboxypeptidase A2 (pancreatic) (Cpa2), insulin 1 (Ins1), chymotrypsin C (caldecrin) (Ctrc), carboxyl ester lipase (Cel), Colipase, pancreatic (Clps). The expression levels of these transcripts were determined relative to β-actin (Actb). The values range from a two- to five-fold change across all treatment groups when compared with the controls. Ctrc in low dose (50 mg kg-1) and clps in medium dose (75 mg kg-1) genes were not significantly altered. Although, these expression levels were much smaller than those noted from microarray data, the qPCR data supports trends observed in global gene expression analysis.
Earlier research from our group that examined cytological, immuno-cytochemical and histological data, we suggested that 2-AA exerts its toxicity effects via an independent mechanism that was yet to be defined (Boudreau et al., 2006). Our current research findings seem to point to the role of 2-AA in the dysregulation of several pancreatic genes that regulate lipid and protein metabolism in a way that involves a feedback mechanism which may ultimately lead to insulin resistance (Funaki, 2009) and tissue autolysis. We also report that 2-AA directly suppresses Ins1 and Ins2 gene expression with prolonged exposure leading to symptomology associated with insulin-dependent diabetes. Further, we hypothesize that dysregulation of these lipases produces an excess of free fatty acids in pancreatic cells from their catalytic activity, which in turn also alters glucose metabolism and its subsequent insulin signaling. Riserus et al. (2009) notes that glucose metabolism is influenced by free fatty acids since they can alter cell membrane function, enzyme activity, insulin signaling and gene expression. Boudreau et al. (2006) reported necrosis of both the endocine and exocrine tissues of the pancreas. The over-expression of proteases reported here could lead to such non-specific tissue necrosis. Modulations of genes associated with oncogene activation, apoptosis and inflammatory responses were not observed in the present experiments. Further analysis is ongoing to examine gene ontology as part of quantitative trait analysis, class comparison analyses and hierarchical cluster analysis in order to discern other pathways that may be important in elucidating 2-AA pancreas toxicity and diabetogenic activity.
ACKNOWLEDGMENT
The authors would like to thank the staff of the Southern Illinois University’s Laboratory Animal Program. We are grateful to Dr. Richard Thomas for allowing us to use his Chromo4 system instrument for determining relative expression values by qPCR. We are also grateful to Ms. Emilia Zargham for helping with the feeding study.
REFERENCES
- Al-Bahrani, A.Z. and B.J. Ammori, 2005. Clinical laboratory assessment of acute pancreatitis. Clin. Chim. Acta, 362: 26-48.
PubMed - Baker, D.G., H.W. Taylor, S.P. Lee, S.A. Barker, M.E.P. Goad and J.C. Means, 2001. Hepatic toxicity and recovery of Fischer 344 rats following exposure to 2-aminoanthracene by intraperitoneal injection. Toxicol. Pathol., 29: 328-332.
PubMed - Bansal, P. and Q. Wang, 2008. Insulin as a physiological modulator of glucagon secretion. Am. J. Physiol. Endocrynol. Metab., 295: E751-E761.
CrossRefDirect Link - Bott, S., Y.A. Shafagoj, P.T. Sawicki and T. Heise, 2005. Impact of smoking on the metabolic action of subcutaneous regular insulin in type 2 diabetic patients. Horm. Metab. Res., 37: 445-449.
PubMed - Boudreau, M.D., D.G. Baker, H.W. Taylor, S.A. Barker and J.C. Means, 2001. Suppression of arylamine toxicity in the Fischer-344 rat following ingestion of a complex mixture. Toxicol. Pathol., 29: 333-343.
PubMed - Boudreau, M.D., H.W. Taylor, D.G. Baker and J.C. Means, 2006. Dietary exposure to 2-aminoanthracene induces morphological and immunocytochemical changes in pancreatic tissues of Fisher-344 rats. Toxicol. Sci., 93: 50-61.
CrossRefDirect Link - Burke, J.E. and E.A. Dennis, 2009. Phospholipase A2 biochemistry. Cardiovasc. Drugs Ther., 23: 49-59.
CrossRefPubMedDirect Link - Cerami, C., H. Founds, I. Nicholl, T. Mitshuhashi and D. Giordano et al., 1997. Tobacco smoke is a source of toxic reactive glycation products. Proc. Natl. Acad. Sci. USA., 94: 13915-13920.
PubMed - Chari, S.T., 2007. Detecting early pancreatic cancer: Problems and prospects. Semin. Oncol., 34: 284-294.
CrossRefPubMedDirect Link - Chen, R., S. Pan, R. Aebersold and T.A. Brentnall, 2007. Proteomics studies of pancreatic cancer. Proteomics Clin. Applied, 1: 1582-1591.
Direct Link - Chen, R., S. Pan, K. Cooke, K.W. Moyes and M.P. Bronner et al., 2007. Comparison of pancreas juice proteins from cancer versus pancreatitis using quantitative proteomic analysis. Pancreas, 34: 70-79.
PubMed - Chilton, F.H., A.N. Fonteh, M.E. Surette, M. Triggiani and J.D. Winkler, 1996. Control of arachidonate levels within inflammatory cells. Biochim. Biophys. Acta, 1299: 1-15.
PubMed - Chipitsyna, G., Q. Gong, R. Anandanadesan, A. Alnajar and S.K. Batra et al., 2009. Induction of osteopontin expression by nicotine and cigarette smoke in the pancreas and pancreatic ductal adenocarcinoma cells. Int. J. Cancer, 125: 276-285.
PubMed - Elayat, A.A., M.M. El-Naggar and M. Tahir, 1995. An immunocytochemical and morphomometric study of the rat pancreatic islets. J. Anat., 186: 629-637.
Direct Link - Eliasson, B., S. Attvall, M.R. Taskinen and U. Smith, 1997. Smoking cessation improves insulin sensitivity in healthy middle-aged men. Eur. J. Clin. Invest., 27: 450-456.
PubMed - Hedstrom, L., 1996. Trypsin: A case study in the structural determinants of enzyme specificity. Biol. Chem., 377: 465-470.
Direct Link - Hui, D.Y., 1996. Molecular biology of enzymes involved with cholesterol ester hydrolysis in mammalian tissues. Biochim. Biophys. Acta, 1303: 1-14.
PubMed - Hui, D.Y. and P.N. Howles, 2002. Carboxyl ester lipase: Structure-function relationship and physiological role in lipoprotein metabolism and atherosclerosis. J. Lipid Res., 43: 2017-2030.
CrossRefDirect Link - Kido, Y., J. Nakae and D. Accili, 2001. The insulin receptor and its cellular targets. J. Clin. Endocrinol. Metabol, 86: 972-979.
Direct Link - Ma, W., C. Tang and L. Lai, 2005. Specificity of trypsin and chymotrypsin: Loop motion controlled dynamic correlation as a determinant. Biophys. J., 89: 1183-1193.
CrossRefDirect Link - Nemoda, Z. and M. Sahin-Toth, 2006. Chymotrypsin C (caldecrin) stimulates autoactivation of human cationic trypsinogen. J. Biol. Chem., 281: 11879-11886.
CrossRefDirect Link - Perez-Chacon, G., A.M. Astudillo, D. Balgoma, M.A. Balboa and J. Balsinde, 2009. Control of free arachidonic acid levels by phospholipase A2 and lysophospholipid acyltransferases. Biochim. Biophys. Acta, 1791: 1103-1113.
CrossRef - Riserus, U., W.C. Willet and F.B. Hu, 2009. Dietary fats and prevention of type 2 diabetes. Prog. Lipid Res., 48: 44-51.
CrossRefPubMedDirect Link - Soleimani, S., F.F. Azarbaizani and V. Nejati, 2007. The effect of Equisetum arvense L. (Equisetaceae) in histological changes of pancreatic β-cells in streptozotocin-induced diabetic in rats. Pak. J. Biol. Sci., 10: 4236-4240.
CrossRefPubMedDirect Link - Snyderwine, E.G., H.A.J. Schut, R.H. Adamson, U.P. Thorgeirsson and S.S. Thorgeirsson, 1992. Metabolic activation and genotoxicity of heterocyclic arylamines. Cancer Res., 52: 2099s-2102s.
Direct Link - Tseng, P.J., 2009. Human exposure to perfluorinated compounds via smoking and second-hand smoke. J. Applied Sci., 9: 2475-2479.
CrossRefDirect Link - Wang, T., K. Shankar, M.J. Ronis and H.M. Mehendale, 2007. Mechanisms and outcomes of drug- and toxicant-induced liver toxicity in diabetes. Crit. Rev. Toxicol., 37: 413-459.
PubMed - Wang, X.L. and J. Wang, 2005. Smoking-gene intereaction and disease development: Relevance to pancreatic cancer and atherosclerosis. World J. Surg., 29: 344-353.
PubMed - Whitcomb, D.C. and M.E. Lowe, 2007. Human pancreatic digestive enzymes. Dig. Dis. Sci., 52: 1-17.
CrossRefDirect Link - Witt, H. and E. Bhatia, 2008. Genetic aspects of tropical calcific pancreatitis. Rev. Endocr. Metab. Disord., 9: 213-226.
PubMed - Zhu, Y., S. Bye, P.J. Stambrook and J.A. Tischfield, 1995. Aflatoxin b1 2-aminoanthracene and 7,12-dimethylbenz[a]anthracene-induced frameshift mutations in human APRT. Environ. Mol. Mutagen., 26: 234-239.
CrossRefDirect Link - Zuhlke, H., D.F. Steiner, A. Lernmark and C. Lipsey, 1976. Carboxypeptidase B-like and trypsin-like activities in isolated rat pancreatic islets. Ciba. Found Symp., 41: 183-195.
PubMed - Funk, C.D., 2001. Prostaglandins and leukotrienes: Advances in eicosanoid biology. Science, 294: 1871-1875.
CrossRef - Pfaffl, M.W., G.W. Horgan and L. Dempfle, 2002. Relative Expression Software Tool (REST©) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res., Vol. 30, No. 9e36.
CrossRefDirect Link