
M. Najafi
Department of Biochemistry, College of Medicine
Isfahan University of Medical Sciences, Isfahan, Iran
A.A. Moshtaghie M. Ani
Department of Biochemistry, College of Medicine
Isfahan University of Medical Sciences, Isfahan, Iran
M. Shabani
Department of Biochemistry, School of Medicine
Iran University of Medical Sciences, Hemmat High Way, Tehran, Iran
Journal of Pharmacology and Toxicology
Year: 2008 | Volume: 3 | Issue: 2 | Page No.: 163-167







ABSTRACT
The current study was designed to investigate the effects of cadmium administration on the AST (Aspartate aminotransferase) and its isoenzyme activities in the serum and liver for durations of 15 and 60 days, respectively. AST isoenzymes were separated by gel filtration chromatography technique and evaluated kinetically. Results showed significant increases in the serum AST activities up to 47 and 38.35% upon Cd administrations of 0.25 and 0.5 mg kg-1, respectively. This increase was not time and dose dependent in the long period. At the end of each period, the specific activity both isoenzymes in the serum increased significantly (p< 0.05) while in the liver, mitochondrial AST activity increased as compared to cytosolic AST activity. We concluded that the total serum AST activity was not dose and time dependent. However, the changes of liver AST isoenzymes in the short and long periods might be due to hepatotoxicity following oxidative stress and delayed synthesis of AST isoenzymes, respectively.
PDF Abstract XML References Citation
How to cite this article
M. Najafi, A.A. Moshtaghie M. Ani and M. Shabani, 2008. Study of Aspartate Aminotransferase Activity in Intoxified Rat by Cadmium. Journal of Pharmacology and Toxicology, 3: 163-167.
DOI: 10.3923/jpt.2008.163.167
URL: https://scialert.net/abstract/?doi=jpt.2008.163.167
DOI: 10.3923/jpt.2008.163.167
URL: https://scialert.net/abstract/?doi=jpt.2008.163.167
INTRODUCTION
It is well documented that Cadmium (Cd) is a heavy transition metal involved in the human intoxication (Zhou et al., 2004; Joseph et al., 2001; Pulido and Parrish, 2003). It's high distribution along with other chemicals in environment come from industrial processes. It has been reported that Cd is readily absorbed through the lung and gastrointestinal tract and metabolized in the body (Bressler et al., 2004; Bridges and Zalups, 2005). It's presence in hepatocytes interfere with cellular function (Karmakar et al., 1999; Shaikh et al., 1999). The amount of trapped Cd in liver is excreted via bile (Klaassen et al., 1999). It is also returned into blood and transferred into the kidneys (Satarug et al., 2006), so that microalbuminuria, glucosuria and phosphaturia occur following Cd overload in the kidneys (Järup, 2003). The damaging effect of Cd on cell organelles is primarily manifested in membrane deterioration, resulting in the release of some important enzymes into the serum (Blasco and Puppo, 1999; Boguszewska and Pasternak, 2004). One of these enzymes is Aspartate aminotransferase (AST, EC 2.6.1.1) that has two isoenzymes namely cytosolic aspartate aminotransferase (c-AST) and mitochondrial aspartate aminotransferase (m-AST) (Hiroyasu and Hiroyasu, 1978). It has been reported that CdCl2 administration in rat model led to the changes in AST activity (El-demerdash et al., 2004; Kowalczyk et al., 2003).
Since AST isoenzymes are important from clinical point of view thus, the present study was designed to investigate the effects of different concentrations of cadmium on AST activity and its isoforms in the rat serum and liver by intraperitoneal administration in short and long periods.
MATERIALS AND METHODS
A total of 40 adult male albino wistar rats weighing 200-250 g were purchased from Isfahan University of Medical Sciences, housed for two weeks and fed with standard chow and water ad libitum.
All the chemical (analytical grades) and enzymes were purchased from Sigma Chemical Co. (USA) and Man Co. (Iran). In each period of experiment, the rats were divided into three groups (5 each). Treated groups received intraperitoneally cadmium (doses of 0.25 and 0.5 mg kg-1 body weight) as cadmium chloride in 0.2 mL saline and control group injected only saline solution daily. At the end of each period, blood samples were collected in pre-washed plastic tubes for determination of the total serum AST activity and separation of the serum AST isoenzymes. For preparation of rat liver subcellular fractions, the livers were immediately dissected out, trimmed, washed in ice cold saline. A known weight of the tissue was homogenized in 5 mL of ice cold sucrose (0.25 M) and centrifuged at 12000 g for 15 min. The supernatant (post-mitochondrial fraction) was stored at -20°C. Pellet (mitochondrial fraction) was also washed in ice cold saline several times and incubated in Triton X-100 (1%). Then, it was diluted in Tris-HCl buffer (0.025 M, pH 7.5) and stored at -20°C. The AST isoenzymes were separated by an improved gel filtration chromatography technique, using sephacryl S-300 column (1x50 cm) (Moshtaghie et al., 1995) and evaluated kinetically (concentrations for L-Aspartate were 0.025 by 0.1 mM and for α-Ketoglutarate were 0.13 by 1 mM). After loading the serum and liver subcellular fractions, the column was eluted with the same Tris-HCl buffer and 30 collected tubes were contained 2 mL of the buffer each. The AST activity assayed spectrophotometrically at 505 nm (Reitman and Frankek, 1957) and 340 nm (Karmen, 1995). Protein was also determined colorimetrically (Lowry et al., 1951).
Statistical Analysis
Data were expressed as mean±SD. The student unpaired t-test was used to estimate differences in means of the total serum AST activity and changes of isoenzymes activity between the groups, using Epi Ver. 6.1. A value of p<0.05 was considered statistically significant.
RESULTS AND DISCUSSION
In vitro finding showed that concentration of cadmium less than 5 mM had no any effect on AST activity (data not shown). Two peaks for serum AST activities were obtained while mitochondrial fraction showed one peak (Fig. 1). The Kmapp for both substrates α-Ketoglutarate and L-Aspartate (first peak) were 0.12 and 0.37 mM, respectively. Furthermore, the Kmapp for the same substrates (second peak) were 0.24 and 0.1 mM, respectively.
| Table 1: | Effects of cadmium administrations on serum AST activity |
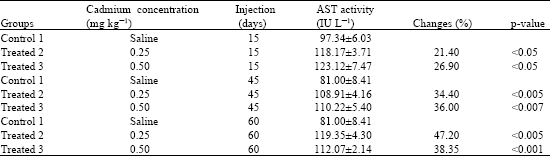 | |
| Values are expressed as mean±SD; p<0.05 means significant. Group 1 as control. Group 2 treated with 0.25 and Group 3 treated with 0.5 mg kg-1 of cadmium. | |
| Table 2: | Changes in AST isoenzymes activities in serum and liver |
 | |
| m-AST: Mitochondrial AST, c-AST: Cytosolic AST | |
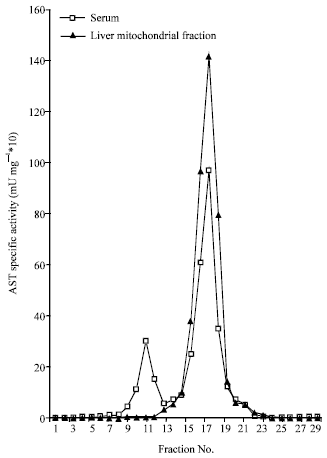 | |
| Fig. 1: | Specific activity profiles of AST isoenzymes |
As shown in Table 1, there was a significant (p<0.05) increase in the level of total AST serum activity for the periods of 15, 45 and 60 days as compared to the control group. Cadmium administrations of 0.25 and 0.5 mg for 60 days increased the serum AST activity up to 47.2 and 38.35%, respectively. There was a significant (p<0.05) increase in the c-AST activity of serum for both 0.25 and 0.5 mg concentrations of cadmium on 15 days period as compared to the 45 and 60 days periods (Table 2). In addition, the c-AST activity in liver had a significant (p<0.05) increase from days 45 to 60. Also, the m-AST activity of serum increased during the study, but, its changes in the liver was insignificant (p>0.05).
Several investigators have evaluated the effects of Cd on cellular dysfunction in different tissues (Zhou et al., 2004; Joseph et al., 2001; Goering et al., 1994). Cd promotes an oxidative stress and forms molecular complexes that would cause to change function of intracellular organelles leading to cell dead (Pulido and Parrish, 2003; Shaikh et al., 1999). There is no any report on changes of AST isoenzymes after Cd administration in short and long periods.
In vitro results based on kinetic data in agreement with Terrance (1874) revealed that the first and second peaks were corresponded with c-AST and m-AST isoenzymes, respectively.
Present data showed an increase in the serum AST activity which supports the finding of Kowalczyk et al. (2004). This increase might be due to the enzyme leakage from cells into blood. This increase was not dose and time dependent, because, higher dose (0.5 mg kg-1) of cadmium showed slight activity as compared with the lower dose (0.25 mg kg-1) of cadmium for 60 days period. Since, cadmium in concentrations of less than 5 mM did not affect the activity of AST, thus we suggest that the decreased AST in 60 days period, might be due to urinary excretion of AST from the kidneys. It is important to mention that delayed failure of the kidneys by cadmium administration was already described (Järup, 2002).
We assume the increase of serum m-AST and c-AST isoenzymes of serum during the continuation of treatment might be due to cell damage leading to mitochondrial dysfunction. In addition, its synthesis mainly in the hepatocytes may contribute to increased level of m-AST. It should be noted that cadmium toxicity in the liver was previously described (Rana et al., 1996). The decreased specific activity changes in the liver c-AST as compared to the liver m-AST in the short term (15 days treatment) might be due to impairment in the membrane of hepatocytes. On the other hand, in the long terms (45 and 60 days), both isoenzymes were increased in the serum and hepatocytes. Probably, some inductive mechanisms involved in anabolic reactions in the liver that have led to inhibition of oxidative stress (Hollis and Hogstrand, 2001) and excretion of cadmium (Klassen et al., 1999) might have been increased activity of AST isoenzymes in hepatocytes.
In conclusion, the present data suggest that increased serum AST activity was not dependent on time and dose of administrated cadmium. Furthermore, the changes of liver AST isoenzymes might be due to early hepatotoxicity following oxidative stress and delayed synthesis of AST isoenzymes.
REFERENCES
- Blasco, J. and J. Puppo, 1999. Effect of heavy metals (Cu, Cd and Pb) on aspartate and alanine aminotransferase in Ruditapes philippinarum (Mollusca: Bivalvia). Comp. Biochem. Physiol. C, 122: 253-263.
Direct Link - Boguszewska, A. and K. Pasternak, 2004. Cadmium-influence on biochemical processes of the human organism. Biometals, 17: 519-523.
Direct Link - Bressler, J.P., L. Olivi, J.H. Cheong, Y. Kim and D. Bannona, 2004. Divalent metal transporter 1 in lead and cadmium transport. Ann. N. Y. Acad. Sci., 1012: 142-152.
Direct Link - Bridges, C.C. and R.K. Zalups, 2005. Molecular and ionic mimicry and the transport of toxic metals. Toxicol. Appl. Pharmacol., 204: 274-308.
CrossRefDirect Link - El-Demerdash, F.M., M.I. Yousef, F.S. Kedwany and H.H. Baghdadi, 2004. Cadmium-induced changes in lipid peroxidation, blood hematology, biochemical parameters and semen quality of male rats: Protective role of vitamin E and β-carotene. Food Chem. Toxicol., 42: 1563-1571.
CrossRefPubMedDirect Link - Hollis, L., C. Hogstrand and C.M. Wood, 2001. Tissue-specific cadmium accumulation, metallothionein induction and tissue zinc and copper levels during chronic sublethal cadmium exposure in juvenile rainbow trout. Arch. Environ. Contam. Toxicol., 41: 468-474.
CrossRefDirect Link - Järup, L., 2003. Hazards of heavy metal contamination. Br. Med. Bull., 68: 167-182.
CrossRefDirect Link - Joseph, P., T.K. Muchnok, M.L. Klishis, J.R. Roberts, J.M. Antonini, W.Z. Whong and T. Ong, 2001. Cadmium-induced cell transformation and tumorigenesis are associated with transcriptional activation of c-fos, c-jun and c-myc proto-onco genes: Role of cellular calcium and reactive oxygen species. Toxicol. Sci., 61: 295-303.
Direct Link - Karmakar, R., A. Banerjee, S. Datta and M. Chatterjee, 1999. Influence of cadmium intoxication on hepatic lipid peroxidation, glutathione level and glutathione S-transferase and gamma-glutamyl transpeptidase activities: Correlation with chromosome aberrations in bone marrow cells. J. Environ. Pathol. Toxicol. Oncol., 18: 277-287.
- Klaassen, C.D., J. Liu and S. Choudhuri, 1999. Metallothionein: An intracellular protein to protect against cadmium toxicity. Annu. Rev. Pharmacol. Toxicol., 39: 267-274.
CrossRefDirect Link - Kowalczyk, E., A. Kopff, P. Fijalkowski, M. Kopff and J. Niedworok, 2003. Effect of anthocyonins on selected biochemical parameters in rats exposed to cadmium. Acta Biochem. Pollut., 50: 543-548.
Direct Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Pulido, M.D. and A.R. Parrish, 2003. Metal-induced apoptosis: Mechanisms. Mutat. Res./Fundam. Mol. Mech. Mutagen., 533: 227-241.
CrossRefDirect Link - Rana, S.V., R. Singh and S. Verma, 1996. Protective effects of few antioxidants on liver function in rats treated with cadmium and mercury. Indian J. Exp. Biol., 34: 177-179.
PubMed - Reitman, S. and S. Frankel, 1957. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol., 28: 56-63.
CrossRefPubMedDirect Link - Satarug, S., M. Nishijo, J.M. Lasker, R.J. Edwards and M.R. Moore, 2006. Kidney dysfunction and hypertension: Role for cadmium, P450 and heme oxygenases? Tohoku J. Exp. Med., 208: 179-202.
Direct Link - Shaikh, Z.A., T.T. Vu and K. Zaman, 1999. Oxidative stress as a mechanism of chronic cadmium-induced hepatotoxicity and renal toxicity and protection by antioxidants. Toxicol. Applied Pharmacol., 154: 256-263.
CrossRefPubMedDirect Link - Zhou, T., X. Jia, R.E. Chapin, R.R. Maronpot, M.W. Harris, J. Liu, M.P. Waalkes and E.M. Eddy, 2004. Cadmium at a non-toxic dose alters gene expression in mouse testes. Toxicol. Lett., 154: 191-200.
Direct Link