
Shiv Kumar Dubey
Center for Biotechnology, University of Allahabad, Allahabad-21 1, India
Archana Pandey
CMP Degree College, University of Allahabad, Allahabad-21 1, India
Ashok Kumar Bajaj
Bajaj Skin Clinic, Panna Lal Road, Allahabad-211002, India
Krishna Misra
Nucleic Acid Research Lab, Department of Chemisfry
University of Allahabad, Allahabad-211002, India
Journal of Pharmacology and Toxicology
Year: 2007 | Volume: 2 | Issue: 8 | Page No.: 718-724







ABSTRACT
The colour of mammalian skin is determined by many factors and mainly by the degree and distribution of melanin pigmentation. Tyrosinase is the key enzyme for melanin biosynthesis. Contact with different azo dyes is well known to produce contact dermatitis and some times depigmentation of the skin. In the present study we have reported the effect of known depigmenting dyes as well as some food, cosmetic and drug dyes on the activity of tyrosinase enzyme. The activity of the enzyme has been assessed in terms of oxidation of DOPA. The inhibitory effect of PPD was found to be maximum i.e., 85% while solvent yellow 3 and Brilliant crocin showed inhibition of 70 and 60%, respectively. The PPD and solvent yellow 3 were identified as noncompetitive inhibitors of tyrosinase by Linweaver Burk plot.
PDF Abstract XML References
How to cite this article
Shiv Kumar Dubey, Archana Pandey, Ashok Kumar Bajaj and Krishna Misra, 2007. Some Commercial Azo Dyes as Inhibitors of Mushroom Tyrosinase DOPA Oxidase Activity. Journal of Pharmacology and Toxicology, 2: 718-724.
DOI: 10.3923/jpt.2007.718.724
URL: https://scialert.net/abstract/?doi=jpt.2007.718.724
DOI: 10.3923/jpt.2007.718.724
URL: https://scialert.net/abstract/?doi=jpt.2007.718.724
INTRODUCTION
The color of human skin depends on a number of factors which include the thickness of epidermis, the amount of blood supply, the degree of vasodilation and the amount of pigment present. Different endogenous pigments, such as melanin, oxyhaemoglobin, reduced haemoglobin and carotene present in the dermis, epidermis and subcutaneous tissue may contribute to the color of the normal human skin (Riley, 2003; Schroder, 1995). However, the major factor responsible for the degree of coloration of skin is the melanin content of melanocytes, the specific epidermal cells containing the organelle, the melanosome where melanin is synthesized. Melanin is secreted by melanocyte, which are distributed in basal layers of the dermis (Bolognia and Orlow, 2003; Sulaimon and Kitchell, 2003).
Tyrosinase (EC 1.14.18.1) is a melanogenic copper containing enzyme that catalyzes the transformation of tyrosine to DOPAquinone (Solano et al., 2006; Xie et al., 2003), thus plays important role in the formation of melanin pigment during melanogenesis (Ando et al., 2007; Marmol and Beermen, 1996; Jimbow et al., 2000; Riley, 1993). Melanogenesis is mainly regulated by tyrosinase and dopachrome tautomerase. Tyrosinase is a key enzyme for melanin biosynthesis in plants and animals. Tyrosinase inhibitors can therefore be clinically useful for the treatment of some dermatological disorders associated with melanin hyperpigmentation. They have become increasingly important as cosmetic (Schurink et al., 2007) and medicinal products (Hwang and Lee, 2007; Miao et al., 1997) primarily to control melanin pigmentation.
Significant information is available from literature on the modification of skin pigmentation due to contact with variety of physical and chemical agents.
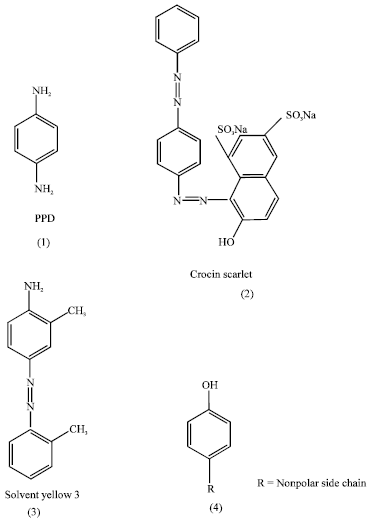 | |
| Fig. 1: | p,p’-benzene substituted azo dyes interacting with CT-DNA and having potent tyrosinase inhibitory effect causing depigmentation |
Paraphenylendiamine (PPD) (Fig. 1) a building block for azo dyes and major component of hair dyes has recently been implicated in producing depigmentation (Taylor et al., 1993; Bajaj et al., 1996). Black Henna touted to be a herbal product also contains approximately 16% PPD leading to depigmentation at the site of patch test after 1-3 months. Alta, a scarlet red solution applied by a certain sect of Indian woman on their feet during religious and social functions does produce the depigmentation at the site of constant application as was reported by our research group (Bajaj et al., 1998). We reported that an alta component viz., Crocin scarlet MOO, an azo dye (Fig. 1 and 2) to produce depigmentation at the application site. Later it was found that Solvent yellow 3 (Fig. 1and 3), another azo dye (4’-amino-2’, 3’-dimethyl azo benzene or o-amino azo toluene), known as Fast Garnet GBC and used for textile dying also produces depigmentation at patch test site (Bajaj et al., 2000). The p-substituted phenols e.g., p-tertiary butyl phenol (PTBP) also produces depigmentation (Angeline et al., 1983; Bajaj et al., 1990).
Regarding mechanism of change in pigmentation of skin by contact with chemicals nothing significant has been reported in literature. In order to study the effect of these dyes on human skin we have conducted experiments. Earlier we have studied (Bajaj et al., 2004) the interaction of these dyes with Calf Thymus DNA in vitro in order to find out whether these pigments interact with DNA at molecular level and proposed a simple spectroscopic technique for scanning the depigmenting potential of azo dyes.
 | |
| Fig. 2: | Chemical structure of azo dyes screened for tyrosinase inhibitory studies |
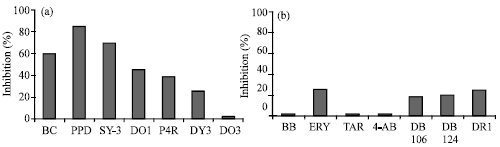 | |
| Fig. 3a and b: | Comparative inhibitory effect of azo dyes on tyrosinase enzyme |
In this communication we have studied the inhibitory effect of these dyes on tyrosinase enzyme responsible for melanogenesis by assessing Dopa formation i.e., evaluation of tyrosinase activity by assessing Dopa quantitatively.
MATERIALS AND METHODS
Materials
Mushroom tyrosinase and L-dopa (3, 4-dihydroxy phenyl alanine) were purchased from Aldrich Chemicals (USA). Azo dyes and PPD were purchased from Aldrich and Fluka. All chemicals used were of analytical grade.
Assay for Tyrosinase Activity
Inhibition of tyrosinase activity was measured by determining change in its DOPA oxidase activity using a modification of the method reported by Shono and Toda (1981). Test substances (dyes) were dissolved in methanol to a concentration of 1 mM. L-DOPA, 600 μL (8 mM, dissolved in 67 mM phosphate buffer, pH 6.8) and 200 μL of each dye solution, 100 μL of buffer, 100 μL of mushroom tyrosinase was added to make the volume to 1 mL.
The reaction mixture was incubated at 37°C for 30 min and the amount of dopachrome formed was determined spectrophotometricaly at 475 nm and inhibitory activity was calculated according to following formula:
Where, C:The absorbance of the control at 475 nm and S is the absorbance of the sample at the same wavelength.
Inhibition Pattern of Mushroom Tyrosinase by PPD and Solvent Yellow 3
The kinetic behaviour of dyes causing inhibition of the enzyme i.e PPD (85%) and Solvent yellow-3 (70%) was studied by assaying the oxidation of L-DOPA. When different concentratios of L-DOPA were used as substrate, PPD and SY-3 decreased Vmax value of tyrosinase in a dose dependent manner but did not change the Km value. Therefore, PPD and Solvent Yellow-3 were identified as non competitive inhibitors by Linweaver-Burk plot analysis (Fig. 4 and 5).
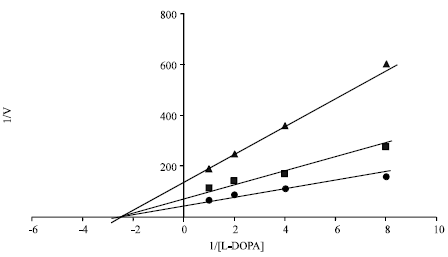 | |
| Fig. 4: | Linweaver-Burk plots of mushroom tyrosinase activity changes caused by PPD 200 μM (▲), 100 μM (■) and blank (●) |
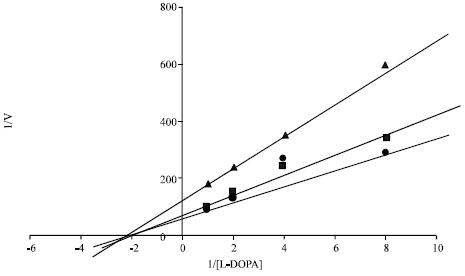 | |
| Fig. 5: | Linweaver-Burk plots of mushroom tyrosinase activity changes caused by Solvent Yellow 3 200 μM (▲), 100 μM (■) and blank (●) |
| Table 1: | Inhibitory effect of azo dyes on mushroom tyrosinase |
 | |
RESULTS
A number of dyes were screened for their inhibitory potential which is presented in Table 1. The majority of azo dyes inhibited tyrosinase activity. PPD has highest inhibitory effect, while Solvent Yellow-3 and Brilliant Crocin shows, respectively 70 and 60% inhibition of the enzyme. Some other azo dyes also showed inhibitory activities (Table 1) while some had no change at conc. of 200 μM (Bar diagrams are shown in Fig. 3a and b).
DISCUSSION
The primary material of melanin biosynthesis is tyrosine which is subsequently oxidized to dopaquinone, catalysed by tyrosinase enzyme. Thus inhibition of tyrosinase can regulate melanin synthesis. The non-production of melanin may be due to mimicking (Isosterism) of tyrosine by the p-substituted phenols or amines (PPD) or the PPD unit of azo dyes Although p-substituted benzene nucleus is an essential structural component of all azo dyes as evident from Fig. 1 and 2 the structural similarity between all the azo dyes used and PPD may explain the cross sensitization of some dyes of azo group which are reported to cross react with para amino benzene compound and cause group sensitization. The p-substitution of benzene ring may be the minimum requirement for enzyme binding resulting in decoloration of skin. It appears that optimal depigmentation from substituted phenols occur when one position of an aromatic ring is hydroxylated and position-4 has a nonpolar side chain (Mcguvie and Hendee, 1971) (Fig. 1 and 4). Their structural similarity (Fig. 1, (4) and Fig. 2, (1-8)) to tyrosine, the building block of melanin is important. This can further be evaluated by seeing the effect of dyes on cultured melanocytes from human skin.
ACKNOWLEDGMENT
One of author (S.K.D) wishes to thank UGC. India for providing financial assistance by a scholarship during the course of this study.
REFERENCES
- Ando, H., K. Hirofumi, M. Ichihashi and V.J. Hearing, 2007. Approaches to identify inhibitors of melanin biosynthesis via the quality control of tyrosinase. J. Invest. Dermatol., 127: 751-761.
CrossRefDirect Link - Anqeline, E., O.C. Marineno, A.M. Carrozzo, L. Bianchi, A. Delogu, G. Giannelo and G. Nini, 1983. Allergic contact dermatitis of the lip margins from p-tertiarybutylphenol in a lipliner. Contact Dermatol., 28: 146-148.
CrossRef - Bajaj, A.K., A. Misra, K. Misra and S. Rastogi, 2000. The azo dye Solvent Yellow3 produces depigmentation. Contact Dermatitis, 42: 237-238.
Direct Link - Bajaj, A.K., S.K. Dubey and K. Misra, 2004. A simple spectroscopic technique proposed for scanning the depigmenting potential of azo dyes. Contact Dermatitis, 51: 97-98.
CrossRefDirect Link - Del Marmol, V. and F. Beermen, 1996. Tyrosinase and related proteins in mammalian pigmentation. FEBS Lett., 381: 165-168.
CrossRef - Hwang, J.H. and B.M. Lee, 2007. Inhibitory effects of plant extracts on tyrosinase, L-DOPA oxidation and melanin synthesis. J. Toxicol. Environ. Health, 70: 393-407.
Direct Link - Jimbow, K., J.S. Park, F. Kato, K. Hirosaki, K. Toyofuku, C. Hua and T. Yamashita, 2000. Assembly, target-signaling and intracellular transport of tyrosinase gene family proteins in the initial stage of melanosome biogenesis. Pigment Cell Res., 13: 222-229.
CrossRefDirect Link - Mcguvie, J. and J. Hendee, 1971. Biochemical basis for depigmentation of skin by phenolic germicides. J. Invest. Dermatol., 57: 256-261.
CrossRefDirect Link - Riley, P.A., 1993. Mechanistic aspect of the control of tyrosinase activity. Pigment Cell Res., 6: 182-185.
PubMedDirect Link - Schurink, M., W.J. van Berkel, H.J. Wichers and C.G. Boeriu, 2007. Novel peptides with tyrosinase inhibitory activity. Peptides, 28: 485-495.
CrossRefDirect Link - Solano, F., S. Briganti, M. Picardo and G. Ghanem, 2006. Hypopigmenting agents: An updated review on biological, chemical and clinical aspects. Pigment Cell Res., 19: 550-571.
CrossRefDirect Link - Taylor, J.S., H.I. Maibach, A.A. Fisher and W.F. Bergfeld, 1993. Contact leucoderma associated with the use of hair colors. Cutis, 52: 273-280.
PubMed