
R.P. Jaya Prasanthi
Department of Zoology, SV University, SVTJ Campus, Tirupati, AP 51 7 SC)2 India
G. Hariprasad Reddy
Department of Zoology, SV University, SVTJ Campus, Tirupati, AP 51 7 SC)2 India
C.S. Chetty
Department of Natural Sciences and Mathematics, Savannah State University, Savannah, GA 31404
G. Rajarami Reddy
Department of Zoology, SV University, SVTJ Campus, Tirupati, AP 51 7 SC)2 India
Journal of Pharmacology and Toxicology
Year: 2006 | Volume: 1 | Issue: 3 | Page No.: 270-277







ABSTRACT
Developing nervous system has long been recognized as a primary target for lead (Pb2+) induced toxicity. We have examined the effect of calcium (Ca2+) and zinc (Zn2+) supplementation on Pb2+-induced alterations in adenosine triphosphatase (ATPase) activities in the developing mouse brain from postnatal day (PND) 14 to 90. Albino mice were lactationally exposed to low (0.2%) and high (1%) levels of lead acetate via the drinking water of the mother. Pb2+-exposure was commenced on PND 1, continued up to PND 21 and stopped at weaning. Ca2+ and Zn2+ (0.02% in 0.2% Pb2+-water or 0.1% in 1% Pb2+-water) were supplemented separately to the mother up to PND 21. The activities of Mg2+ATPase in mitochondrial fraction and Na+K+ATPase in P2-fraction of the cerebral cortex, hippocampus, cerebellum and medulla were assayed. The specific activity of both ATPases increased gradually in an age-dependent manner in the control brain. Pb2+-exposure significantly inhibited the activities of both Mg2+ATPase and Na+K+ATPase in different brain regions of the developing brain. The inhibition was more pronounced at PND 28 in the brain regions of mice exposed to 1% Pb2+. Ca2+ or Zn2+-supplementation significantly reversed the Pb2+-inhibited activity of both ATPases. These results indicate that dietary Ca2+/Zn2+-supplementation decreases intestinal absorption of Pb2+ and there by reduces the neurotoxic effects of Pb2+.
PDF Abstract XML References
How to cite this article
R.P. Jaya Prasanthi, G. Hariprasad Reddy, C.S. Chetty and G. Rajarami Reddy, 2006. Influence of Calcium and Zinc on Lead-induced Alterations in ATPases in the Developing Mouse Brain. Journal of Pharmacology and Toxicology, 1: 270-277.
DOI: 10.3923/jpt.2006.270.277
URL: https://scialert.net/abstract/?doi=jpt.2006.270.277
DOI: 10.3923/jpt.2006.270.277
URL: https://scialert.net/abstract/?doi=jpt.2006.270.277
Introduction
Na+K+ATPase and Mg2+ATPase have a relatively high sensitivity to certain classes of heavy metals and other pollutants and it has been shown that toxicosis from pollutants may develop primarily from ATPase inhibition (Sushma, 1999). A number of studies have shown that ATPase activity was inhibited by heavy metals such as Pb2+ (Singerman, 1976), cadmium (Valle and Ulmer, 1972; Tucker and Matte, 1980) and methyl mercury (Schmidt-Niensen, 1974). Heavy metal binding to sulfhydryl (-SH) groups has often been implicated in the inhibition of Na+K+ATPase which maintains cellular fluid balance and provides the electrochemical gradients essential for synaptic potentials and action potentials (Sweadner and Goldin, 1980; Carfagna et al., 1996).
Pb2+ is known to exert its neurotoxic effects by competing with Ca2+ for its receptors coupled with second messenger functions (Hammond et al., 1984; Bressler and Goldstein, 1991) and in some cases, inhibits the actions of Ca2+ as a regulator of cell function (Bressler and Goldstein, 1991). Bogden et al. (1992) have demonstrated that a dietary excess of Ca2+ reduces Pb2+ -absorption. However, this effect is not as dramatic as the effect of Ca2+ insufficiency on Pb2+ uptake and retention. Therefore, the achievement of adequate rather than excessive dietary Ca2+ seems to be more useful in combating Pb2+-intoxication (Peraza et al., 1998). Similar to Ca2+, Zn2+ also competes for Pb2+ and effectively reduces the availability of binding sites for its uptake (Flora and Tandon, 1990). Therefore, present study was designed to examine the influence of Ca2+ and Zn2+ on Pb2+-induced alterations in the specific activities of Mg2+ ATPase and Na+K+ ATPase in different regions of developing mouse brain.
MATERIALS AND METHODS
Chemicals
Chemicals used in this study were obtained from Sigma, St. Louis, MO, USA.
Animal Exposure
Swiss albino pregnant mice were purchased from Indian Institute of Science, Bangalore, India. All pups were pooled on PND 1 and new litters consisting of eight males were randomly selected and placed with each dam. Young mice were lactationally exposed to low-level (0.2%) and high-level (1%) Pb2+ by adding lead acetate to deionized drinking water of the mother. Pb2+-exposure was commenced on PND 1, continued up to PND 21 and stopped at weaning. Control mice received only deionized water without Pb2+.
Calcium and Zinc-Supplementation
Ca2+/Zn2+-supplemented to Pb2+ (0.02% in 0.2% Pb2+-water and 0.1% in 1% Pb2+-water) was separately given to mothers upto PND 21 and stopped at weaning.
Isolation of Tissues
The control, Pb2+-treated and Ca2+/Zn2+-supplemented mice were sacrificed at PND 14, PND 21, PND 28 and at 3 months age. The different brain regions such as cortex, hippocampus, cerebellum and medulla were quickly isolated under ice cold conditions and were stored at -80°C for biochemical analysis.
Preparation of Mitochondrial and P2 fractions
The mitochondrial and P2 fractions of cortex, hippocampus, cerebellum and medulla were prepared using ficoll-sucrose gradients as described by Cotman and Matthews (1971).
Determination of ATPase Activity
The specific activities of Na+K+ ATPase in P2 fraction and mitochondrial Mg2+ATPase were determined as described by Tirri et al. (1973). The enzyme activity was expressed as Fmoles of inorganic phosphate (Pi) formed/mg protein/hour. The Pi was estimated by the method of Fiske and Subba Row (1925). The protein content was estimated by the method of Lowry et al. (1951).
Statistical analysis
Standard statistical procedures such as student t-test and ANOVA were used to analyze the data for significance level.
RESULTS
The specific activities of Mg2+ ATPase and Na+ K+ ATPase in brain regions of the control mice increased with increase in age from PND 14 to 3 months (Table 1 and 2). The cortex region documented higher enzyme activities followed by cerebellum, hippocampus and medulla. Pb2+ decreased the specific activities of both Mg2+ ATPase and Na+K+ ATPase in all the brain regions examined. The inhibition was greater in the brain of mice exposed to 1% Pb2+ as compared to the mice exposed to 0.2% Pb2+. The decrease in Mg2+ ATPase was more pronounced in the hippocampus of mice exposed to 1% Pb2+ where as the maximum inhibition of Na+K+ ATPase was observed in cerebellum (Table 1 and 2). The decrease in enzyme activities was found to be higher at PND 14. However, the enzyme activity slowly increased and reached almost control levels in three months old brain.
The data show that the Ca2+/Zn2+-supplementation reduced the Pb2+-inhibition of Mg2+ ATPase and Na+ K+ ATPase activities observed in all the brain regions of mice (Table 1 and 2). The protective effect of Ca2+/Zn2+-supplementation from Pb2+-toxicity was greater in the mice exposed to 0.2% Pb2+ as compared to 1% Pb2+.
| Table 1: | Alterations in Mg2+ATPase activities in brain of young and adult mice exposed to Pb2+ and supplemented with different concentrations of Ca2+ or Zn2+ |
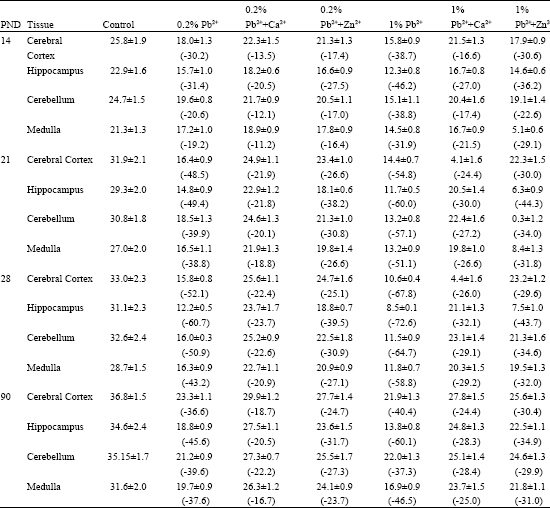 | |
| (Each value is mean±6 observations. PND: Postnatal day. Values in parentheses are % change over control. All changes over respective controls are statistically significant (p<0.5)) | |
| Table 2: | Alterations in Na+K+ATPase activities in brain of young and adult mice exposed to Pb2+ and supplemented with different concentrations of Ca2+ or Zn2+ |
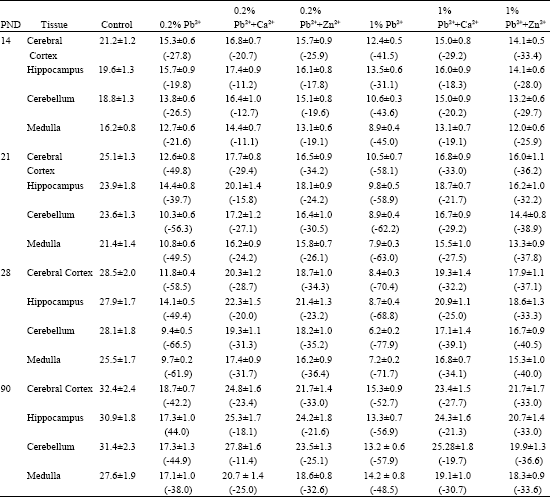 | |
| (Each value is mean ± 6 observations. PND: Postnatal day. Values in parentheses are % change over control. All changes over respective controls are statistically significant (p<0.5)) | |
The mice supplemented with Ca2+ exhibited greater recovery of Pb2+-inhibited ATPase activity as compared to the mice supplemented with Zn2+. The alterations in both the enzyme activities were found to be statistically significant (Table 1 and 2).
DISCUSSION
Heavy metal cations such as Pb2+ can bind to a number of sites on proteins including imidazole, histidyl, carbonyl and especially -SH side chains (Winder and Kitchen, 1984). Pb2+ has been reported to inhibit Na+K+ ATPase of mammalian tissues (Cardone et al., 1971) and also interferes with mitochondrial function and blocks the O2 uptake. In the present study, lactational Pb2+-exposure exerted inhibitory effect on both Mg2+ ATPase and Na+ K+ ATPases in the developing mouse brain in an age- and dose-dependent manner. The decrease in the Mg2+ATPase activity could be attributed to the Pb2+-induced modulations in the oxidative metabolism (Boyer, 1977). Alterations in mitochondrial structure and in neurotransmission were observed in rats chronically exposed to low-doses of Pb2+ (Jablonska et al., 1994; Struzynska et al., 1994). These changes were correlated with changes in energy metabolism of synaptic mitochondria during Pb2+-toxicity.
The data from the present study suggest that Pb2+ significantly inhibited the specific activities of Mg2+ ATPase as well as Na+K+ ATPase which also support the findings of Cardone et al. (1971). Earlier studies have linked the inhibition of ATPases to the binding of heavy metal cations to -SH groups (Barcellos et al., 1994; Battacharya et al., 1997). Chetty et al. (1990) have reported that Pb2+ has high affinity for free –SH groups in enzymes and proteins and its binding can alter their functions.
Pb2+ may exert an inhibitory effect directly on Na+K+ ATPase. It is known that brain Na+k+ ATPase is among several enzymes particularly affected by Pb2+ (Siegel et al., 1977; Fox et al., 1991; Struzynska et al., 1994). Our earlier studies reported that the decrease in Na+K+ ATPase activity can change the gradients of Na+ and K+ across the cell membrane and alter the neurotransmitters levels (Reddy et al., 2003; Devi et al., 2005). Pb2+ is known to produce oxidative stress by generating ROS. Na+K+ ATPase was reported to be inhibited by Reactive Oxygen Species (ROS) in the brain (Rohn et al., 1993; Sagara et al., 1996). Free radicals might destroy –SH groups or may act as inhibitors for these enzymes resulting in decreased enzyme activity.
The high specific activity of ATPases in the cortex, cerebellum and hippocampus regions of the brain suggests the involvement of these regions in different behavioral functions. It is known that the activity levels of ATPases parallel the metabolic demands of different regions of mouse brain and the differential sensitivity to Pb2+-induced neurotoxicity in these brain regions is not due to a preferential Pb2+-accumulation, but possibly could be due to alteration of biochemical or cellular processes that are uniquely associated with, or greatly enhanced in a particular region (Widzowski and Cory-Slechta, 1994; Moreira et al., 2001).
The greater inhibition of ATPase activities in young (up to PND 28) mice could be due to the fact that Pb2+ passes through the blood-brain barrier easily during the early developmental period. The permeability of blood brain barrier is over 1,000 times greater for Pb2+ than for Ca2+ (Bradbury and Deane 1993). A significant decrease in ATPase activities in adult mice even after the withdrawal of Pb2+-exposure may be due to the already accumulated Pb2+ in these regions of brain.
Pb2+ and Ca2+ interactions occur at the cellular and molecular levels and Pb2+ has the ability to mimic or displace Ca2+ during specific physiological processes. Pb2+ competes with Ca2+ resulting in: a) inhibition of neurotransmitter release, b) alterations in the regulation of cell metabolism by binding to second messenger Ca2+ receptors, c) blocking channels and d) inhibition of ATP-dependent Ca2+-and Na+-pumps (Luthman et al., 1994; Bettaiya et al., 1996). Ca2+/Zn2+-supplementation reduced the Pb2+-effects on both ATPases. It is likely that the supplemented Ca2+/Zn2+ may compete for similar binding sites as that of Pb2+. The use of Ca2+ supplements to Pb2+-exposed women during lactation has been shown to blunt the blood Pb2+ levels in children (Pires et al., 2002). In our recent study (Prasanthi et al., 2005), we have also reported the protective effect of Ca2+/Zn2+ against Pb2+-toxicity in mouse brain. It has been shown that supplementation with Ca2+/Zn2+ decreases gastrointestinal absorption of Pb2+ and reduces its tissue accumulation (Cerlewski and Forbes, 1976; Peraza et al., 1998). Thus, Ca2+ and Zn2+ replace Pb2+ in the body and thereby reduce the Pb2+-burden in the body.
Greater recovery of Pb2+-inhibited ATPase activity in mice exposed to 0.2% Pb2+ suggests that the detoxification mechanisms in mice were strong enough to counter the effects of low-level Pb2+-exposure as compared to the mice exposed to 1% Pb2+. From the present study, it is evident that developmental Pb2+-exposure inhibited the Mg2+ATPase and NA+K+ ATPase enzymes in a dose-dependent manner and Ca2+/Zn2+-supplementation significantly reversed the Pb2+-induced alterations in brain Mg2+ATPase and Na+K+ ATPase activities.
ACKNOWLEDGMENTS
This study was supported by a UGC Grant No. F3-113/2003, SR (II) and a CSIR Grant No 37(1136)/03/EMR-II
REFERENCES
- Barcellos, C.K., M.R.C. Schetinger, A.M.O. Battastini, L.B. Silva, R.D. Dias and J.J.F. Sarkis, 1994. Inhibitory effect of cadmium acetate on synaptosomal ATP diphosphohydrolase from adult rat cerebral cortex. Braz. J. Med. Biol. Res., 27: 1111-1115.
Direct Link - Bertoni, J.M. and P.M. Sprenkle, 1991. Low level lead inhibits the human brain cation pump. Life Sci., 48: 2149-2156.
CrossRef - Bettaiya, R., P.R. Yallapragada, E. Hall and S. Rajanna, 1996. In vitro effect of lead on Ca2+-ATPase in synaptic plasma membranes and microsomes of cerebral cortex and cerebellum. Ecotoxicol. Environ. Saf., 33: 157-162.
PubMed - Bhattacharya, S., S. Bose, B. Mukhapadhyay, D. Sarkar and D. Das et al ., 1997. Specific binding of inorganic mercury to Na+K+-ATPase in rat liver plasma membrane and signal transduction. Biometals, 10: 157-162.
CrossRef - Bogden, J.D., S.B. Gertner, S. Christakos, F.W. Kemp, Z. Yang, S.R. Katz and C. Chu, 1992. Dietary calcium modifies concentrations of lead and other metals and renal calbindin in rats. J. Nutr., 122: 1351-1360.
PubMedDirect Link - Bradbury, M.W. and R. Deane, 1993. Permeability of the blood brain barrier to lead. Neurotoxicology, 14: 131-136.
PubMed - Bressler, J.P. and G.W. Goldstein, 1991. Mechanisms of lead neurotoxicity. Biochem. Pharmacol., 41: 479-484.
PubMedDirect Link - Carfagna, M.A., G.D. Ponsler and B.B. Muhoberac, 1996. Inhibition of ATPase activity in rat synaptic plasma membranes by simultaneous exposure to metals. Chem. Biol. Interact., 100: 53-65.
CrossRef - Chetty, C.S., B. Rajanna and S. Rajanna, 1990. Inhibition of rat brain microsomal Na+K+-ATPase and ouabain binding by mercuric chloride. Toxicol. Lett., 51: 109-116.
Direct Link - Devi, C., G. Reddy and C. Chetty, 2005. Developmental lead exposure alters mitochondria monoamine oxidase and synaptosomal catecholamine levels in rat brain. Int. Dev. Neurosci., 23: 375-381.
Direct Link - Fiske, C.H. and Y. Subbarow, 1925. The colorimetric determination of phosphorus. J. Biol. Chem., 66: 375-400.
CrossRefDirect Link - Flora, S.J.S. and S.K. Tandon, 1990. Beneficial effects of zinc supplementation during chelation treatment of lead intoxication in rats. Toxicology, 64: 129-139.
CrossRefPubMedDirect Link - Fox, D.A., S.D. Rubinstein and P. Hsu, 1991. Developmental lead exposure inhibits adult rat retinal, but not kidney, Na+ K+ATPase. Toxicol. Appl. Pharmacol., 109: 482-493.
Direct Link - Hammond, P.B., R.L. Bornschein, H. Zenick, 1984. Toxicology considerations in the assessment of lead exposure. Neurotoxicology, 5: 53-66.
PubMed - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Luthman, J., E. Lindqvist, G.A. Gerhardt, L. Olson and B.H. Hoffer, 1994. Alterations in central monoamine systems after postnatal lead acetate treatment in rats. Environ. Res., 65: 100-118.
PubMed - Moreira, E.G., I. Vassilieff and V.S. Vassilieff, 2001. Developmental lead exposure: Behavioral alterations in the short and long term. Neurotoxicol. Teratol., 23: 489-495.
CrossRef - Pires, J.B., N. Miekeley and C.M. Donangelo, 2002. Calcium supplementation during lactation blunts erythrocyte lead levels and aminolevulinic acid dehydratase zinc reactivation in women non-exposed to lead and with marginal calcium intakes. Toxicology, 175: 247-255.
CrossRef - Prasanthi, R.P.J., G.H. Reddy, C.B. Devi and G.R. Reddy, 2005. Zinc and calcium reduce lead induced perturbations in the aminergic system of developing brain. BioMetals, 18: 615-626.
CrossRefPubMedDirect Link - Reddy, G.R., M.R. Basha, C.B. Devi, A. Suresh and J.L. Baker et al., 2003. Lead induced effects on acetylcholinesterase activity in cerebellum and hippocampus of developing rat. Int. J. Dev. Neurosci., 21: 347-352.
PubMed - Sagara, Y., R. Dargusch, F.G. Klier, D. Schubert and C. Behl, 1996. Increased antioxidant enzyme activity in amyloid beta protein-resistant cells. J. Neurosci., 16: 497-505.
PubMed - Sweadner, K.J. and S.M. Goldin, 1980. Active transport of sodium and potassium ions: Mechanism, function and regulation. N. Engl. J. Med., 302: 777-783.
PubMed