
Omar M.E. Abdel Salam
Department of Pharmacology, National Research Centre, Tahrir St., Dokki, Cairo, Egypt
Siham El-Batran
Department of Pharmacology, National Research Centre, Tahrir St., Dokki, Cairo, Egypt
Ayman R. Baiuomy
Department of Pharmacology, National Research Centre, Tahrir St., Dokki, Cairo, Egypt
Journal of Pharmacology and Toxicology
Year: 2006 | Volume: 1 | Issue: 3 | Page No.: 259-269







ABSTRACT
The anti-nociceptive effect of melatonin on visceral pain evoked by i.p. acetic acid injection in mice was investigated. Melatonin (1-8 mg kg-1, s.c.) caused a dose-dependent inhibition of the acetic acid-induced writhes (by 84 to 98.6%). The inhibition of writhing response by melatonin (4 mg kg-1) was not reduced by co-administration of theα-2 adrenoceptor antagonist yohimbine (4 mg kg-1, i.p.). In contrast, the α-1 adrenoceptor antagonist doxazocin (16 mg kg-1, s.c.) reduced the anti-writhing effect of melatonin, whereas the α 1-adrenoceptor antagonist prazosin (2 mg kg-1, i.p.) was without effect. The effect of melatonin was slightly reduced by beta-adrenoceptor blockade with propranolol (2 mg kg-1, i.p.) or adrenergic neuron blockade with guanethidine (8 or 16 mg kg-1, i.p.). The muscarinic receptor antagonist atropine (2 mg kg-1 i.p.), the potassium channel blocker glibenclamide (5 mg kg-1, i.p.), the opioid antagonist naloxone (5 mg kg-1, i.p.) or the non-selective adenosine receptor antagonist, theophylline (10 mg kg-1 i.p.), failed to reduce the anti-writhing effect of melatonin. The antinociceptive effect of melatonin was slightly reduced by co-administration of the central dopamine D2 receptor antagonist sulpiride (15 mg kg-1, i.p.) (though not by haloperidol, 2 mg kg-1, i.p.) and to more extent by the dopamine D2 receptor agonist bromocryptine (1.5 mg kg-1, i.p.), whereas metoclopramide (3 mg kg-1, i.p.), chlorpromazine (3 mg kg-1, i.p.), domperidone (10 mg kg-1, i.p.) or blockade of dopamine D1 receptors by clozapine (5 mg kg-1, i.p.) had no effect. In conclusion, melatonin exerts potent anti-nociceptive effects on visceral pain in mice. The antinociception caused by melatonin is subject to modulation by noradrenergic and dopaminergic neural pathways.
PDF Abstract XML References
How to cite this article
Omar M.E. Abdel Salam, Siham El-Batran and Ayman R. Baiuomy, 2006. Noradrenergic and Dopaminergic Modulation of Melatonin Visceral Antinociception in Mice. Journal of Pharmacology and Toxicology, 1: 259-269.
DOI: 10.3923/jpt.2006.259.269
URL: https://scialert.net/abstract/?doi=jpt.2006.259.269
DOI: 10.3923/jpt.2006.259.269
URL: https://scialert.net/abstract/?doi=jpt.2006.259.269
INTRODUCTION
Melatonin (N-acetyl-5-methoxytryptamine) is a product of the pineal gland as well as other organs such as the gut (Reiter, 1995; Messner et al., 2001). Melatonin and its oxidation products are direct free radical scavengers and potent indirect antioxidants (Rodriguez et al., 2004; Allegra et al., 2003; Silva et al., 2004). In experimental models, melatonin exhibited marked anti-inflammatory and analgesic properties and poteniated those of non-steroidal anti-inflammatory drugs (Cuzzocrea et al., 1999; El-Shenawy et al., 2002). Visceral pain is a common form of pain but is poorly understood. The polymodal peripheral receptors initiate unpleasant sensations that can be modulated in the dorsal horn and the anterior columns of the spinal cord before reaching the cerebral cortex (Al-Chaer and Traub, 2002). The complexity of pain perception reflects the presence of both nociceptive and antinociceptive systems that modulate nociceptive input at many levels of the neuraxis. Descending brainstem systems influence nociceptive stimuli spinally while other systems modulate nociceptive input supraspinally (Pasternak, 1998).
There is evidence to suggest that melatonin-induced antinociception is mediated through melatonin receptors within the central nervous system. Intraperitoneal administration of either 1.25, 2.5, 5.0 or 10.0 mg kg-1 melatonin induced a dose-dependent inhibition of spinal wind-up activity, probably through hyperpolarization of dorsal horn neurons subsequent to melatonin binding to membrane receptors and/or via intracellular interference with a NMDA receptor-dependent nitric oxide generating pathway (Laurido et al., 2002). It has been suggested that melatonin interferes with the NMDA-mediated glutamatergic component of pain transmission in rat spinal cord by acting on MT2 receptors (Noseda et al., 2004). The aim of this study thus was to investigate the effect of melatonin in the model of visceral pain induced by acetic acid injection in mice and to pharmacologically characterize and investigate the possible neural pathways involved.
MATERIALS AND METHODS
Animals
Swiss male albino mice weighing 20-22 g were used in all experiments. Animals were acclimatized to the laboratory for at least 1h before testing and were used once during the experiments. Procedures were in accordance with the Institutional Animal Care Committee and followed guidelines on ethical standards for investigations of experimental pain in conscious animals (Zimmermann, 1983).
Acetic Acid-induced Writhing
Separate groups of 6 mice each were administered vehicle and/or melatonin (1, 2, 4 or 8 mg kg-1, s.c.). After the appropriate pretreatment interval, an i.p. injection of 0.6% acetic acid was administered (0.2 mL/mice) (Koster, 1959). Each mouse was then placed in an individual clear plastic observational chamber and the total number of writhes made by each mouse was counted for 30 min after acetic acid administration. In addition, the effect of melatonin at 4 mg kg-1was compared to that of equimolar dose of indomethacin (5.16 mg kg-1, s.c.).
In an attempt to elucidate the mechanisms by which melatonin exerts its anti-nociceptive effect, the use of selective antagonists of receptors or drugs that interfere with second messengers and ion channels were employed. The dose of 4 mg kg-1 of melatonin was selected as the standard dose of melatonin for studies on effects of noradrenergic, cholinergic, dopaminergic drug or other drugs.
The effects of the following agents on melatonin-induced anti-writhing were examined; α-1 adrenoceptor antagonist prazosin (2 mg kg-1, i.p.), α-2 adrenoceptor antagonist yohimbine (4 mg kg-1, i.p.), the adrenergic blocker guanethidine (8 mg kg-1 or 16 mg kg-1, i.p.), the beta adrenoceptor antagonist, propranolol (2 mg kg-1, i.p.), the muscarinic acetylcholine receptor antagonist atropine (2 mg kg-1 i.p.), the non-selective opioid receptor antagonist naloxone (5 mg kg-1 i.p.), the non-selective adenosine receptor antagonists theophylline (10 mg kg-1 i.p.), the potassium channel blocker glibenclamide (5 mg kg-1, i.p.), the dopamine D2 receptor antagonists sulpiride (15 mg kg-1, i.p.), haloperidol (2 mg kg-1, i.p.) and domperidone (10 mg kg-1, i.p.), metoclopramide (3 mg kg-1, i.p.) and chlorpromazine (3 mg kg-1, i.p.), the dopamine D2 receptor agonist bromocryptine (1.5 mg kg-1, i.p.) or the dopamine D1 receptor antagonist clozapine (5 mg kg-1, i.p.). The doses of different antagonists and agonists used are those reported in other studies (Gleeson and Atrens, 1982; Ohkubo et al., 1991; Ghelardini et al., 2000; Takeshita and Yamaguchi, 1997; Kowaluk et al., 2000). All drugs except guanethidine were administered together with melatonin 30 min prior to the injection of acetic acid. Guanethidine was administered 1 h prior to the abdominal constriction assay.
Drugs and Chemicals
Melatonin (Sigma, St. Louis, USA), indomethacin (Kahira Pharm and Chem. IND Co., Cairo, A.R.E.), atropine sulfate, yohimbine hydrochloride, propranolol hydrochloride, naloxone hydrochloride (Sigma, St. Louis, USA) were used. Analytical-grade glacial acetic acid (Sigma, St. Louis, USA) was diluted with pyrogen-free saline to provide a 0.6% solution for i.p. injection. All drugs were dissolved in isotonic (0.9% NaCl) saline solution immediately before use, except indomethacin which was dissolved in a 5% solution of sodium bicarbonate.
Statistical Analyses
Data are expressed as mean±SE. Data were analyzed by one way analysis of variance, followed by a Tukey's Multiple Range Test for post hoc comparison of group means. When there were only two groups a two-tailed Student's t-test was used. For all tests, effects with a probability of p<0.05 were considered to be significant.
RESULTS
The intraperitoneal injection in mice of 0.1 mL/10 g body weight of a 0.6% (v/v) solution of acetic acid induced a writhing response with 0 and 30 min. Melatonin (1-8 mg kg-1) injected s.c. 30 min before acetic acid significantly inhibited the nociceptive response in a dose-dependent manner by 84 to 98.6%.
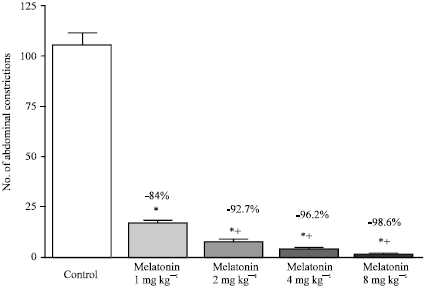 | |
| Fig. 1: | Effect of different doses of melatonin administered as 30 min pretreatment on the number of writhes in the abdominal constriction assay. Six mice were used per each group. Asterisks indicate significant change from the control group. The plus (+) sign indicates significant change from the 1 mg kg-1 treated group |
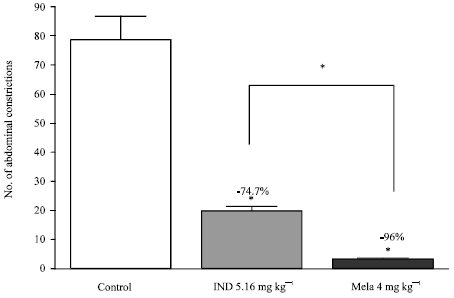 | |
| Fig. 2: | Effect of equimolar doses of indomethacin and melatonin administered as 30 min pretreatment on the number of writhes in the abdominal constriction assay. Six mice were used per each group. * p<0.05 compared to control and between indomethacin and melatonin treated groups |
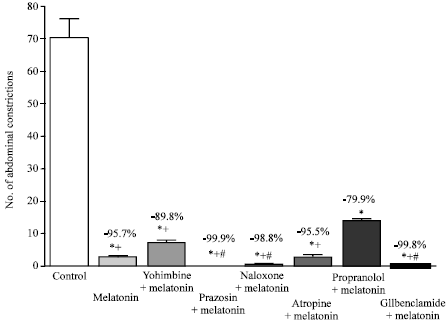 | |
| Fig. 3: | Effect of yohimbine (4 mg kg-1, i.p.), prazosin (2 mg kg-1, i.p.), naloxone (5 mg kg-1, i.p.), atropine (2 mg kg-1, i.p.), propranolol (2 mg kg-1, i.p.) and glibenclamide (5 mg kg-1, i.p.) on anti-nociception induced by melatonin (4 mg kg-1, s.c.) in the abdominal constriction assay. Six mice were used per each group. Asterisks indicate significant change from the control group. The plus (+) sign indicates significant change from the propranolol treated group. The # sign indicates significant change from the yohimbine treated group |
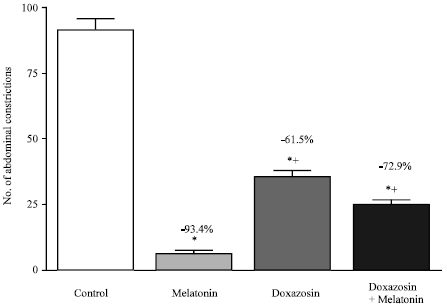 | |
| Fig. 4: | Effect of doxazosin on melatonin (4 mg kg-1, s.c.)-induced anti-nociception in the abdominal constriction assay. Six mice were used per each group. Asterisks indicate significant change from the control group. The plus (+) sign indicates significant change from the melatonin treated group. |
Significant inhibition of the writhing response by 2, 4 or 8 mg kg-1 melatonin was observed compared to 1 mg kg-1treated group (Fig. 1). On a molar basis, melatonin was more potent than indomethacin, a dose of 4 mg kg-1 inhibiting the writhing response by 96% compared to 74.7% inhibition by indomethacin at equimolar dose of 5.16 mg kg-1 (Fig. 2).
Figure 3 shows that treatment with the α-2 adrenoceptor antagonist yohimbine or the α-1 adrenoceptor antagonist prazosin did not significantly change the antinociceptive effect caused by melatonin in the abdominal constriction assay. The non-specific opioid receptor antagonist naloxone, the muscarinic acetylcholine receptor antagonist atropine and the KATP-channel blocker glibenclamide were also without effect. The melatonin-induced anti-writhing effect, however, was reduced by the beta adrenoceptor antagonist, propranolol. Furthermore, mice treated with propranolol exhibited significantly more writhes than yohimbine, prazosin, naloxone, atropine or glibenclamide-treated mice. Meanwhile, mice treated with yohimbine showed significantly more abdominal constrictions than prazosin, naloxone or glibenclamide- treated mice (Fig. 3).
The anti-writhing effect of melatonin was, in addition, reduced in the presense of doxazosin, another α-1 adrenoceptor antagonist. Treatment of mice with doxazosin or doxazosin plus melatonin resulted in significantly more writhes than the melatonin-treated group (Fig. 4). Furthermore, depletion of endogenous catecholamines with guanethidine (8 or 16 mg kg-1) slightly reduced the melatonin-induced anti-writhing (Fig. 5 and 6), although a combination of guanethidine (16 mg kg-1, i.p.) and propranolol (2 mg kg-1, i.p.) failed to alter antinociception by melatonin (Fig. 6). Guanethidine itself resulted in 16.1% (p<0.05) reduction in the number of abdominal constrictions compared with the control group (Fig. 5).
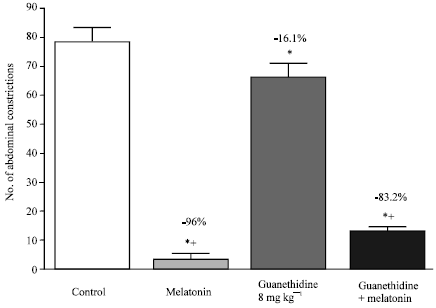 | |
| Fig. 5: | Effect of guanethidine (8 mg kg-1, s.c.) on melatonin (4 mg kg-1, s.c.)-induced anti-nociception in the abdominal constriction assay. Six mice were used per each group. Asterisks indicate significant change from the control group. The plus (+) sign indicates significant change from the melatonin treated group |
 | |
| Fig. 6: | Effect of guanethidine (16 mg kg-1, i.p.) or guanethidine (16 mg kg-1, i.p.) combined with propranolol (2 mg kg-1, i.p.) on anti-nociception induced by melatonin (4 mg kg-1, s.c.) in the abdominal constriction assay. Six mice were used per each group. Asterisks indicate significant change from the control group. The plus (+) sign indicates significant change from the guanethidine treated group |
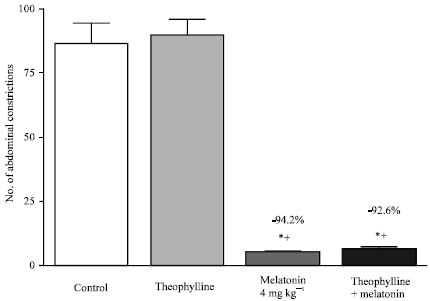 | |
| Fig. 7: | Effect of theophylline (10 mg kg-1, i.p.) on anti-nociception induced by melatonin (4 mg kg-1, s.c.) in the abdominal constriction assay. Six mice were used per each group. Asterisks indicate significant change from the control group. The plus (+) sign indicates significant change from the control or theophylline treated groups |
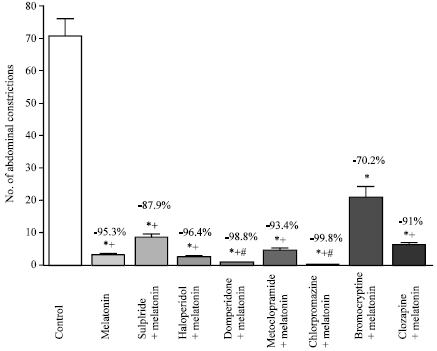 | |
| Fig. 8: | Effect of different dopamine receptor agonists and antagonists on melatonin (4 mg kg-1, s.c.)-induced anti-nociception in the abdominal constriction assay. Six mice were used per each group. Asterisks indicate significant change from the control group. The plus (+) sign indicates significant change from the bromocryptine treated group. The # sign indicates significant change from the sulpiride treated group |
Figure 7 shows that the non-selective adenosine receptor antagonists theophylline failed to reduce the anti-nociceptive effect of melatonin. Figure 8 shows that the dopamine D1 receptor antagonist clozapine and the non-selective dopamine receptor antagonist chlorpromazine had no effect on the anti-writhing effect of melatonin, whereas blockade of central dopamine D2 receptors by sulpiride (though not haloperidol, metoclopramide or the peripherally acting dopamine antagonist, domperidone) or treatment with the dopamine D2 receptor agonist bromocryptine reduced the antinociceptive effect of melatonin. Mice treated with bromocryptine exhibited significantly more abdominal constrictions than those treated with melatonin, sulpiride, haloperidol, domperidone, metoclopramide, chlorpromazine or clozapine. Meanwhile, treatement with sulpiride was associated with significantly more abdominal constrictions than after treatment with domperidone or chlorpromazine.
DISCUSSION
Chemical stimuli applied topically have been employed in studies of visceral pain. A standard pharmaceutical screening tool, the writhing test consists of i.p. injection of dilute (0.6%) acetic acid followed by subsequent counting of “writhes” characteristic contraction of abdominal muscles accompanied by a hind limb extensor motion (Koster, 1959). In the present study we examined the effect of melatonin on visceral nociception evoked in mice by i.p. administration of acetic acid. Results indicate that melatonin potently inhibits visceral nociception. In further steps, we analyzed the possible neural pathways involved in the anti-nociceptive action of melatonin.
Certain pain conditions involve the sympathetic nervous system, e.g., visceral pain due to abdominal and pelvic cancers, ischaemic pain from peripheral vascular disease, arterial spasm or frostbite and others. Sympathetcomy or intravenous regional sympathetic block with guanethidine can be carried out to reduce pain (Serpell, 2005). In the present study, we evaulated the anti-nociceptive effect of melatonin in mice treated with the non-selective β-adrenoceptor blocker propranolol or the adrenergic neurone blocking drug guanethidine. The latter is taken up into adrenergic neurons, where it binds to the storage vesicles and prevent release of neurotransmitter in response to a neuronal impulse, which results in generalized decrease in sympathetic tone. The administration of guanethidine alone resulted in around 16% reduction in the writhing response. In addition, the anti-writhing effect of melatonin was found to be partially reduced in mice treated with propranolol or guanethidine. The results suggest that the analgesic action of melatonin is at least in part due to the enhancement of sympathetic neurotransmission.
The administration of α-2 adrenoceptor agonists produce anti-nociception in rodents by inhibiting synpatic transmission in the spinal cord dorsal horn and there is an evidence of descending noradrenergic system, the stimulation of which, results in the activation of spinal α-2 adrenergic receptors and anti-nociception (Gutierrez et al., 2003; Tanabe et al., 2005). However, the participation of α-2 adrenoceptor involvement in the observed melatonin anti-nociception is unlikely, since yohimbine treatment failed to block the antinociceptive effect of melatonin. The anti-nociception caused by melatonin was reduced by α-1 adrenoceptor antagonism with doxazosin, which might suggest the involvement of α-1 adrenoceptor mechanism (s). Prazosin, another selective α-1 adrenoceptor antagonist, however, failed to reduce the melatonin effect. In another study, the antinociception induced by melatonin (120 mg kg-1, i.p.) in thermally-induced pain in rats was unaffected by intracerebroventricularly administered prazosin, but inhibited by the selective MT2 melatonin receptor antagonist luzindole administered via thec same route (Yu et al., 2000).
The spinal cholinergic system and muscarinic receptors are important for regulation of nociception. Activation of spinal muscarinic receptors produces analgesia and inhibits dorsal horn neurons through inhibition of the glutamatergic synaptic input (Li et al., 2002) and potentiation of synaptic GABA release in the spinal cord (Zhang et al., 2005). The involvement of opioidergic mechanisms or spinal cholinergic system and muscarinic receptors in the anti-nociceptive action of melatonin are ruled out in view of naloxone and atropine-insenstive anti-writhing effect of melatonin. Adenosine is an inhibitory neuromodulator that can increase nociceptive thresholds in response to noxious stimulation (Sawynok, 1998; Dunwiddie and Masino, 2001) and blockade of adenosine receptors by theophylline, a nonselective adenosine receptor antagonists at A1 and A2 (Bruns et al., 1983) is known to induce hyperalgesia (Paalzow, 1994). In the present study, data, however, do not suggest that an adenosine receptor mechanism (s) is involved in the mediation of antinociception induced by melatonin, since theophylline failed to influence the melatonin effect on visceral pain.
Voltage-gated potassium channels, can also contribute to the sensitization of primary afferents observed in gastrointestinal pain states (Cervero and Laird, 2003). The possible involvement of K-channels in the mediation of antinociception induced by melatonin is ruled out, in view of the inability of glibenclamide an ATP-sensitive potassium (KATP) channels blocker to reduce the melatonin effect.
The antinociceptive response of melatonin was also studied in combination with selective dopamine receptor agonists and antagonists. Dopamine D-2 receptors are involved in nociception and analgesic mechanisms and dopamine D2-receptor antagonists e.g., sulpiride caused antinociception in different pain models (Ben-Sreti et al., 1983; Rooney and Sewell, 1989; Michael-Titus et al., 1990; Frussa-Filho et al., 1996; Forman, 1999). In the present study, the analgesic response to melatonin was also partially reduced by administration of the dopamine D2 receptor antagonist, sulpiride and the dopamine D2 receptor agonist bromocryptine, suggesting an interaction at the level of dopamine D2 receptors. Thus it appears that antinociception induced by melatonin results, at least in part, from activation of stimulation of D2 subtypes of the dopamine receptor.
In summary, visceral nociception evoked by i.p. injection acetic acid in mice was markedly inhibited by the administration of melatonin. The antinociception caused by melatonin is subject to modulation by noradrenergic and dopaminergic neural pathways.
REFERENCES
- Al-Chear, E.D. and R.J. Traub, 2002. Biological basis of visceral pain: Recent development. Pain, 96: 221-225.
PubMed - Allegra, M., R.J. Reiter, D.X. Tan, C. Gentile, L. Tesoriere and M.A. Livrea, 2003. The chemistry of melatonin's interaction with reactive species. J. Pineal Res., 34: 1-10.
CrossRefDirect Link - Dunwiddie, T.V. and S.A. Masino, 2001. The role and regulation of adenosine in the central nervous system. Ann. Rev. Neurosci., 24: 31-55.
CrossRefPubMedDirect Link - El-Shenawy, S.M., O.M.E. Abdel-Salam, A.R. Baiuomy, S. El-Batran and M.S. Arbid, 2002. Studies on the anti-inflammatory and anti-nociceptive effects of melatonin in the rat. Pharmacol. Res., 46: 235-243.
CrossRefDirect Link - Koster, R., M. Anderson and E.J. de Beer, 1959. Acetic acid-induced analgesic screening. Fed. Proc., 18: 412-412.
Direct Link - Kowaluk, E.A., J. Mikusa, C.T. Wismer, C.Z. Zhu and E. Schweitzer et al., 2000. ABT-702 (4-Amino-5-(3-bromophenyl) -7-(6- morpholino-pyridin- 3-yl)pyrido[2,3-d]pyrimidine), a novel orally effective adenosine kinase inhibitor with analgesic and anti-inflammatory properties. II. In vivo characterization in the rat. J. Pharmacol. Exp. Ther., 295: 1165-1174.
PubMed - Laurido, C., T. Pelissie, R. Sotomoyano, L. Valladares, F. Flores and A. Hernandez, 2002. Effect of melatonin on rat spinal cord nociceptive transmission. Neuroreport, 13: 89-91.
Direct Link - Rodriguez, C., J.C. Mayo, R.M. Sainz, I. Antolin, F. Herrera, V. Martin and R.J. Reiter, 2004. Regulation of antioxidant enzymes: A significant role for melatonin. J. Pineal. Res., 36: 1-9.
CrossRefPubMedDirect Link - Tanabe, M., K. Takasu, N. Kasuya, S. Shimizu, M. Honda and H. Ono, 2005. Role of descending noradrenergic system and spinal α2-adrenergic receptors in the effects of gabapentin on thermal and mechanical nociception after partial nerve injury in the mouse. Br. J. Pharmacol., 144: 703-714.
CrossRef - Zimmermann, M., 1983. Ethical guidelines for investigations of experimental pain in conscious animals. Pain, 16: 109-110.
CrossRefPubMedDirect Link