
Shiv Kumar Dubey
Center for Biotechnology, University of Allahabad, Allahabad, 211002, India
Krishna Misra
Department of Chemistry, Nucleic Acid Research Laboratory,
University of Allahabad, Allahabad, 211002, India
Alka Tiwari
Department of Pharmacology, M.L.N. Medical College, Allahabad, 211002, India
A.K. Bajaj
Bajaj Skin Clinic, Panna Lal Road, Allahabad, 211002, India
Journal of Pharmacology and Toxicology
Year: 2006 | Volume: 1 | Issue: 3 | Page No.: 234-247







ABSTRACT
Continued contact of some chemicals with the skin is known to cause contact dermatitis and even in exceptional cases contact leucoderma. In our earlier study some of these chemicals specially azo dyes have been found to interact with human (Calf thymus) DNA, probably causing the inactivation of melanocytes. In an effort to find out the mechanism of interaction of some commercial dyes on human skin, the in vitro studies have been reported on the CT-DNA-azo dyes interaction. In the present article the process of melanogenesis, properties and functions of melanin, the enzymes involved in its synthesis as well as chemically induced leucoderma have been reviewed.
PDF Abstract XML References
How to cite this article
Shiv Kumar Dubey, Krishna Misra, Alka Tiwari and A.K. Bajaj, 2006. Chemically Induced Pigmentary Changes of Human Skin, Interaction of Some Azo Dyes with Human DNA. Journal of Pharmacology and Toxicology, 1: 234-247.
DOI: 10.3923/jpt.2006.234.247
URL: https://scialert.net/abstract/?doi=jpt.2006.234.247
DOI: 10.3923/jpt.2006.234.247
URL: https://scialert.net/abstract/?doi=jpt.2006.234.247
INTRODUCTION
The color of human skin depends on a number of factors which include the thickness of epidermis, the amount of blood supply, the degree of vasodilation and the amount of pigment present as reported by Jeghers (1944) and Edwards and Duntley (1939). Different endogenous pigments, such as melanin, oxyhaemoglobin, reduced haemoglobin and carotene present in the dermis, epidermis and subcutaneous tissue may contribute to the color of the normal human skin. However, the major factor responsible for the degree of coloration of skin is the melanin content of melanocytes, the specific epidermal cells synthesizing the melanin containing organelle, the melanosome. The darkness of human skin is directly proportional to the population density of melanocytes.
Formation of Melanins: Melanogenesis
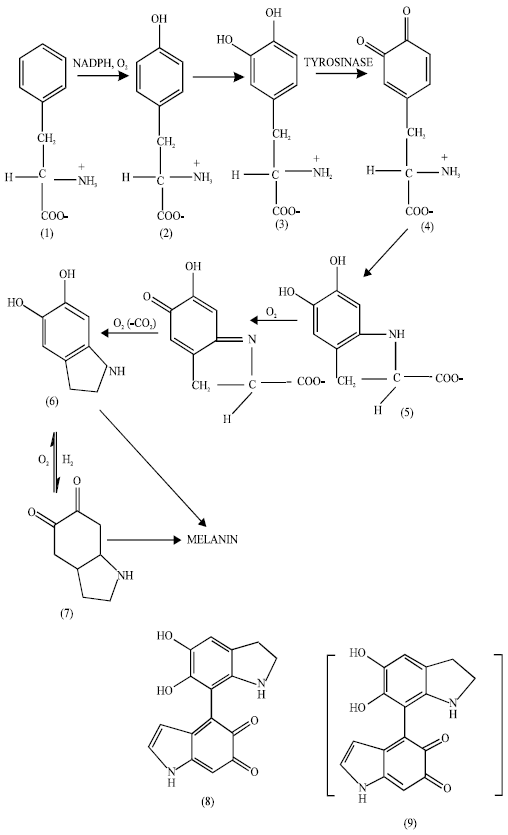
Saji and Iwashita (1963) reported that melanins (latin, black) are synthesized by melanocytes from tyrosine (2) a metabolic product of phenylalanine (1) an essential amino acid (Scheme I). It was shown by Birbeck et al. (1961) that besides tyrosine, a copper containing enzyme, tyrosinase and molecular oxygen also have an important role to play in melanogenesis, a process controlled by pituitary gland. This reaction is also controlled by temperature, pH, redox potential and inhibitors of enzymes.
 | |
| Scheme 1: | Mechanism of formation of melanin: melanogesis |
According to Raper-Mason (Scheme I) tyrosine first gets oxidized to dihydroxy phenylalanine (DOPA) (3), catalyzed by tyrosinase. DOPA, an oxygen activating electron donor, gets further oxidized to DOPAquinone (4). The latter by series of fast spontaneous reactions results in the formation of colorless pigment, leucoDOPAchrome (5) which by subsequent oxidation and decarboxylation forms 5,6-quinone (DOPAchrome) (7) which in turn is converted to 5,6-dihydroxy indole (6) and vice-versa by redox reactions. The various intermediates in this series of fast reactions participate in condensation resulting in the formation of a three-dimensional polymer melanin (Scheme I).
Tyrosine is essential only for the initial stages, the later stages occur in its absence, although their kinetics is influenced by the presence of enzymes. The overall chemical change from tyrosine to melanin requires 5-atoms of oxygen with elimination of 1 mol of CO2. Specific enzymes catalyze each step in the above scheme in vivo.
Extensive studies have been carried out by Fitzpatrick et al. (1961,1963a, 1963b) and Fitzpatrick (1964) on the enzymatic oxidation of DOPA to the final highly insoluble polymeric pigment. On the basis of spectrophotometric studies, it has been suggested that dimer (Scheme I; 8) and oligomer (Scheme I; 9) are the probable intermediates formed during enzymatic oxidation, although these have not been isolated.
However, melanin can be defined as poly-indolequinones of general formula, (C8H302N)n.
Enzymes for Melanogenesis
In lower animals melanin formation is controlled by a polypeptide hormone called melanin-stimulating hormone (α and β-MSH), localized in pituitary gland. In humans, melanogenesis appears to be stimulated by adrenocartico tropic hormone (β-carticotropic; ACTH). The loss of adrenocortical hormones in Addison’s disease leads to a loss of feed back control of ACTH secretion, so that it continues to be secreted at high levels. The melanin stimulating properties of ACTH are attributed to the fact that a large segment of the molecule has the same sequence of amino acids as in MSH.
Tyrosinase is a copper containing mixed function, phenolmonooxygenase and catalyses oxidation of both monophenols (tyrosine) and orthodiphenols (DOPA) to quinones or semiquinones. It utilizes both tyrosine and DOPA as the two electron donors. DOPA being the product of tyrosine oxidation, the result of this enzyme action is the successive oxidation of tyrosine to DOPAquinone.
A radiometric assay for tyrosinase activity has been reported by Chavin in 1963 using enzyme preparations obtained from gold fish skin and labeled tyrosine or DOPA as substrate. The specific activity of tyrosinase is defined as the number of tyrosinase units per Fg of protein nitrogen.
In albinism, which is an inherited disorder of melanin metabolism, there is a decrease or absence of the pigment in the skin and choroids of the eye, due to non-formation of tyrosinase.
Both tyrosine and DOPA have been reported in mammalian skin dialysates, including that of human skin. Sections of human skin turn black when incubated in solutions containing DOPA but not in solutions containing tyrosine. When the epidermis is exposed to ultraviolet radiation, however human skin produces melanin from tyrosine. The radiation probably produces a small amount of DOPA, which primes the tyrosinase reaction producing melanin.
Two tyrosinases have been detected by their behaviour on DEAE- (Diethyl amino ethyl) cellulose and starch gel electrophoresis, but there may be more types in the human skin or human melanoma in all probability.
Properties and Functions of Melanin
The trapping of free radicals by many growing synthetic polymer chains have been reported. It is a common phenomenon in methacrylate polymers. The electro spin resonance (esr) signals shown by natural melanins are attributed to such trapped free radicals. Melanins have been considered as one-dimensional semiconductors with protons acting as electron traps. The biosynthesis of melanins involves a free radical mechanism. DOPA melain shows a strong electron resonance signal at a “g” value of 2.003.The other natural melanins also have “g”value in the same range. Mason (1948) have found that black, brown, red and blond human hair give electron resonance signals at g = 2.003 (black hair has free radical content up to 4.7x1016 spins g-1; dark red hair has 1.8x1015 and blond has only 4.3x1014 spins g-1 dry weight). The spin concentration is increased significantly by sunlight or ultraviolet radiations. Darkening is directly proportional to free radical formation.
The skin of individuals exposed to sunlight for long periods has much more melanins than those living in colder climate, where exposure to sun is much less. This is said to be an important factor for the wide difference in the incidences of skin cancer in white and black skinned people. The former are much more prone to skin cancer (melanomas).
The ultraviolet light which stimulates the synthesis of melanin, is also effectively absorbed by the pigment, resulting in protection of the skin. Melanin acts as an electron exchange polymer capable of protecting tissue against reducing or oxidizing conditions and of trapping such free radicals formed by action of ultraviolet light on skin, which could disturb the metabolism in a normal cell. The melanins protect the skin against the injurious radiations through such mechanism.
Biosynthesis of Melanins
The location and function of a typical melanocyte cell in the human skin has to be considered in view of the different stages of the development of melanin granules.
The polypeptides which subsequently become “tyrosinase” are synthesized in small granules (presumably ribonucleoprotein particles) transferred along endoplasmic reticulum (ER) to the Golgi area and condensed into “protyrosinase” the secondary and tertiary structure of which eventually form tyrosinase. In Golgi area protyrosinase is separated into small units, each of which becomes surrounded by a membranous envelope (ME) and within each envelope the protyrosinase molecule become aligned forming an ordered pattern. This unit is now known as Pro-melanosome (PMS). The protyrosinase then gets activated and forms tyrosinase, melanin biosynthesis begins and the particle is known as melanosome (MS). As melanin gradually accumulates in cytoplasm, the melanosome is eventually transformed into a uniformly dense and structureless particle, the melanin granules (MG) in which no tyrosinase activity has been detected.
Effect of Physical and Chemical Agents on Skin Pigmentation
Significant information is available from literature, as reported by Schwartz (1947), Arndt et al. (1965), Gellin et al. (1970), Kahn et al. (1970), Calnan et al. (1974), Fisher et al. (1976) and Brancaccio et al. (1977), on the modification of skin pigmentation due to contact with a variety of physical and chemical agents. Whether the end result is a gain or loss of melanocyte activity depends on the nature of the inciting agent. Host susceptibility is equally important. Amongst physical causes, heat, cold, ionizing and non ionizing radiations (sunlight, ultraviolet rays) are known to alter skin pigmentation. External contact of skin with certain chemicals or their ingestion in the system may result in the decreased (hypopigmentation) or increased pigmentation (hyperpigmentation), the former leading to contact depigmentation or leucoderma.
Hyperpigmentation
As shown previously by Rook (1951) and Fountain (1967), the most common occupational pigment change is hyperpigmentation. The causes of hyperpigmentation may be chemical and thermal burns (temporary effect), contact with irritants or photosensitivity reactions to fumes of pitch, asphalt and tar. Fitzpatrick et al. (1963) demonstrated that synergistic action by sunlight on these photosensitizes contributes to the tanning effect. Psoralens or furocoumarins present in many fruits and vegetables are reported to be photosensitizers by Pathak et al. (1962). The development of phytophotodermatitis on the sun-exposed skin of harvesters of pink rot fungus (Sclerotina sclerotiorum) infected celery seeds, which release the photoreactive psorolens is well established phenomenon demonstrated by Birmingham et al. (1961). Localised hyperpigmentation has been observed in persons who have had contact with oil of bergamot or limes that contain 5-methoxy psoralen.
Some common etiologic agents causing hyperpigmentation are:
| • | Cancer chemotherapeutic agents |
| • | Antibiotics specially tetracyclines |
| • | Antimalarials |
| • | Metals |
| • | Hormones |
| • | Carotenoids |
| • | Nitro compounds |
| • | Dyes |
| • | Miscellaneous drugs and chemicals |
The use of cancer chemotherapeutic agents has been reported by Losech et al. (1983) to result in hyperpigmentation. All tumor patients using amentanthrone (which is an amino anthroquinone dye) invariably developed diffuse gray-blue color in their skin. However, this color is temporary. The 1, 3-bis (Chlorobutynyl)-1- nitrosourea (BCNU) was reported by Frost et al. (1960) to cause pigmentation in hospital workers who came in contact with it. Blum et al. (1973) and Schuller et al. (1984) reported that bleomycin, a bacterial derived antibiotic caused pigmentary changes in 20% of patients especially when cumulative doses exceed 100 mg. Long term use of tetracyclines particularly minocyclines to treat acne has resulted in numerous reports of skin hyperpigmentation, most commonly blue-black pigmentation of the lower extremities and bluish discoloration of facial acne scars. Sato et al. (1981) reported that the pigmentary change results from a minocycline-melanin complex. The ability of all antimalarials to cause yellow or brown gray pigmentation has been well documented. Chloroquin, hydroxychloroquin and amodiaquine are aminoquinones, which affect approx 25% of patients. However, patients on quinacrine develop a lemon-yellow color which fades in about 4-months after discontinuing the drug.
Exposures or contact with many heavy metals in industry or clinical applications is known to cause hyperpigmentation. Mousels solution (Ferric subsulfate), a cauterizing agent may cause permanent tattooing.
The few hormones that affect pigmentation are well known clinically. The widespread use of estrogen containing oral contraceptive is known to cause melasma.
Carotenoids present in foods are also known to induce skin pigmentation. However, the use of canthaxanthin (orobronze) as an oral “tanning” agent was reported by Suntan et al. (1983) as an exception. However it has been reported that some users of this agent developed crystal deposition in their retina.
There are reports of skin coloration by oral ingestion of some nitro compounds and dyes. Jeghers (1944) reported that during Second World War some serviceman swallowed picric acid to appear ill with jaundice like symptoms to avoid service. Long-term therapy with phenothiazines is a well-known cause of hyperpigmentation.
Hypopigmentation
Hypopigmentation from chemicals and drugs is the result of decrease in the melanin content of the skin. As shown by Cannon et al. (1933) in all cases except for arsenic ingestion, topical use or localized contact of the chemical or drug is the cause of pigment loss. Several chemical and thermal burns (second and third degree) can destroy a sufficient number of melanocytes resulting in loss of pigmentation. Irritants like hydrofluoric acid and caustic dye may cause depigmentation. Skin damage with post inflammatory leucoderma can be caused by a great variety of chemicals. The most important example of allergic contact dermatitis is the contact of skin with poisonivy (Rhus species). In some cases it does not affect pigmentation, in others there may be transient hyperpigmentation, but some cases of pigment loss have been reported by McCarthy et al. (1925). Amongst physical causes, heat, cold, ionizing and non-ionizing radiation (sunlight) is known to alter skin pigmentation.
External contact of skin with certain chemicals or their ingestion in the system may result in decreased (hypopigmentation) or increased pigmentation (hyperpigmentation), the former leading to leucoderma.
Chemically Induced Leucoderma
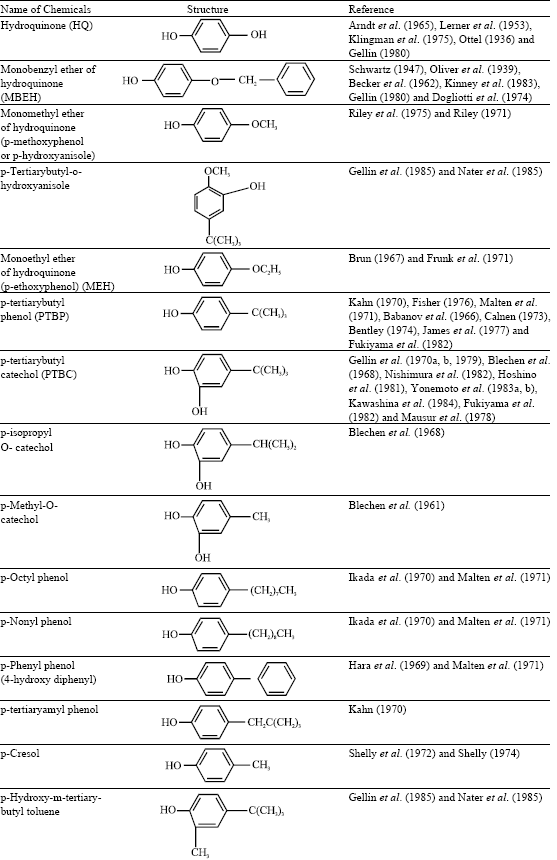
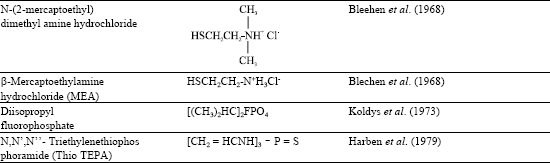
All chemical agents capable of producing contact dermatitis generally leave post inflammatory effects on skin pigment. Pigmentation disorders by occupational hazards have been reported by James et al. (1977) and Gellin et al. (1985, 1987). In 1939 industrially induced depigmentation was first noticed by Oliver et al. on the hands and forearms of workers wearing synthetic rubber gloves containing monobenzyl ether of hydroquinone (MBEH). Based upon the effect, this substance and its parent compound hydroquinone (HQ) have been used therapeutically to depigment the skin as reported by Lerner et al. (1953), Beeker et al. (1962), Arndt et al. (1965), Kligman et al. (1975) and Kinney and Grimes (1983). In 1960’s and 1970’s there were several reports from different countries for hydroquinone and its esters as being responsible for outbreaks of occupational leucoderma (Babanov and Chumakov, 1966; Hara and Nakajima, 1969; Gellin et al., 1970a, b; 1979; Ikeda et al., 1970; Kahn et al., 1970; Malten et al.,1971). Subsequently, many substituted phenols were reported to induce depigmentation in human and animal models, the important ones are tabulated (Table 1). p-Tertiary butyl phenol (TBP), p-tertiary amyl phenol (TAP) were used as antioxidants or rust inhibitors as reported by Gellin et al. (1970 ) and Kahn (1970). These are also used in the manufacture of plastics, resins, lubricants, motor oils, petroleum products, photographic chemicals, insecticides, printing inks, pesticides, disinfectants, synthetic rubber, paints, deodorants and germicides as reported by Gellin et al. (1970a, b, 1979) and Kahn et al. (1970). Depigmentation of breast by wallet and feet by footwear material have been reported by Bajaj et al. (1991).
| Table 1: | Phenols, amines and some phosphorus compounds reported to induce hypopigmentation in human and animal models |
 | |
 | |
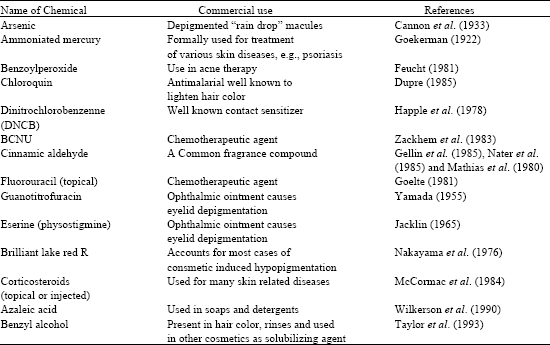
| Table 2: | Miscellaneous depigmenting agents |
 | |
Thin layer chromatography demonstrated presence of MBEH (monobenzyl ether of hydroquinone) as the causative factor. Black gold fish (Fitzpatrick et al., 1963), brown and wild colored guinea pigs (Kahn, 1970; Malten et al., 1971; Riley, 1971; Brun, 1967, 1972; Jimbow et al., 1974; Riley et al., 1975), black guinea pigs (Blechen et al., 1968; Gellin et al. (1970a, b); Malten et al., 1971), black mice (Hara and Nakajima, 1969; Ikeda et al., 1970; Hoshino et al., 1981), white hairless mice (Nishimura et al., 1982; Yonemoto et al., 1983a, b; Kawashima et al., 1984), brown and black cats (Ottel, 1936; Malten et al., 1971) and black rabbits (Babanov and Chumakov, 1966) are the animal models used for studying chemically induced depigmentation (Table 1).
The occupationally related and environmentally associated cases of skin depigmentation generally bear resemblance to vitiligo as reported by James et al. (1977) and are caused by a systemic mechanism. It is morphologically indistinguishable from true vitiligo and its severity is directly proportional to intensity of exposure. In such cases the pigment loss follows contact dermatitis was shown by McCarthy et al. (1925), Fisher (1976) and Gellin et al. (1979). The latter reported that the skin in contact is the first site of depigmentation and the person afflicted is without family history of vitiligo or dermatitis. It is probable that many persons considered to have vitilago have chemically induced leucoderma. Some commercially important miscellaneous depigmenting agents are tabulated in Table 2.
Depigmentation Caused by Some Commercial Azo Dyes
Paraphenylendiamine (PPD) (Fig. I, 1) a building block for azo dyes and major component of hair dyes has recently been implicated in producing depigmentation by Taylor et al. (1993) and Bajaj et al. (1996) and in India such cases of hair dye depigmentation have frequently been observed in the past few years, more so since the introduction of black henna (Kali Mehandi) for dying the hair. Black Henna touted to be an herbal product also contains approximately 16% PPD leading to depigmentation at the site of patch test after 1-3 months. Alta, a scarlet red solution applied by a certain sect of woman on their feet during religious and social function does produce the depigmentation at the site of constant application as was reported by Bajaj et al. (1998). The authors have reported that an alta component viz., Crocein scarlet MOO, an azo dye (Fig. I, 2) to produce depigmentation at the application site. However, Rhodamine-B, a non azo dye present in alta was found to be inert. Later, Bajaj et al. (2000) found that Solvent yellow 3 (Fig. I, 3), another azo dye (4’-amino-2’, 3’-dimethyl azo benzene or o-amino azo toluene), known as Fast Garnet GBC and used for textile dying also produces depigmentation at patch test site.
Although p-substituted benzene nucleus is an essential structural component of all azo dyes as evident from Fig. I, the structural similarity between the two azo dyes (2 and 3) and PPD (1) may explain the cross sensitization of some dyes of azo group which are reported to cross react with para amino benzene compound and cause group sensitization was reported by Anqeline et al. (1983) and Bajaj et al. (1990). The p-substituted phenols e.g. p-tertiary butyl phenol (PTBP) also produces depigmentation, the p-substitution of benzene ring may be the minimum requirement for enzyme binding resulting in decoloration of skin. From a look at Table 1 it appears that optimal depigmentation from substituted phenols occur when one position of an aromatic ring is hydroxylated and position-4 has a nonpolar side chain as was shown by Mcguvie and Hendee (1971). Their structural similarity (Fig. I, 4) to tyrosine (Scheme I, 2), the building block of melanin is important. The inactivation of melanocytes or non-production of melanin may be due to mimicking (Isosterism) of tyrosine by the p-substituted phenols or amines (PPD) or the PPD unit of azo dyes.
Using labeled p-hydroxyanisole it was pointed out by Riley (1975) that it was selectively incorporated into melanocytes grown in tissue culture. It probably reacts with the tyrosinase in the melanosome and then forms a secondary product which diffuses into cytoplasm and kills the cell, since the effect could be reduced or even prevented by tyrosinase inhibitors.
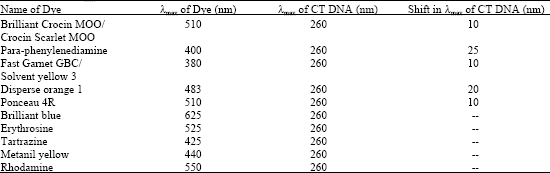
In a series of experiments it has been demonstrated that semiquinine free radicals are formed and initiate lipid peroxidation (a chain reaction) leading to destruction of lipoprotein membranes of the melanocytes and its consequent death was shown by Riley (1971,1975). In order to find out whether this effect is due to the isosterism of these molecules or due to their degradation products viz; ions or free radicals or some other factors we have studied the interaction of some p-substituted azo dyes with mammalian Calf Thymus DNA (CT DNA) by spectroscopic method (Bajaj et al., 2004). A significant shift in λmax of CT DNA was reported by the authors with PPD, Disperse orange 1, Brilliant Crocein MOO, Solvent yellow 3 and Ponceau 4R while no such shift was observed with other dyes (Table 3).
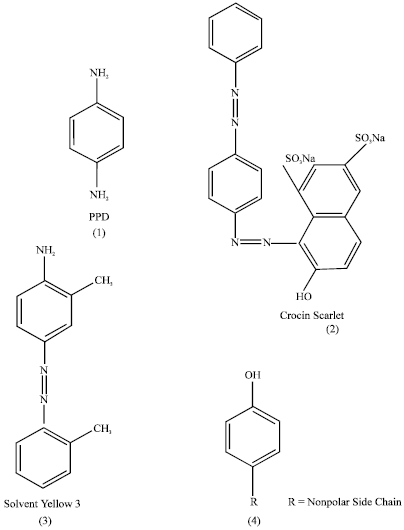 | |
| Fig. I: | Some commercial azo dyes casing depigmentation |
| Table 3: | Shift in λmax on Interaction of azo dyes with CT DNA |
 | |
The interaction of DNA with small molecules (antigens) like drug or dye molecule in its environment may be purely physical like intercalation, which is reversible i.e., reverts back to normal after the antigen is removed. Intercalation involves the insertion of planar molecules between the adjacent DNA base pairs, perpendicular to the double helix backbone. This gives rigidity to the helix. The resulting unwinding of base pairs increases the length of DNA and causes some distortion in back bone. However, in certain cases it may be irreversible too leading to cytotoxicity or cell death. We have found in our preliminary work that PPD and azo dyes like Disperse orange1 and Solvent yellow 3 cause cell death in melanocyte culture. However, these dyes show no interaction with melanoma cell lines L-929 and BF16 except solvent yellow-3. It is however interesting to note that Rhodamine, a non-azo dye which does not give positive patch test, causes apoptosis of melanoma cells (Unpublished results). We have found that dyes like PPD, Disperse orange 1 and Brilliant crocin on incubation with tyrosinase enzyme inhibit its activity. Since this enzyme is significantly responsible for melanin synthesis, these dyes may be the causative factors for hypopigmentation.
ACKNOWLEDGEMENT
One of the authors (SKD) wishes to thank CSIR (India) for a fellowship.
REFERENCES
- Anqeline, E., O.C. Marineno, A.M. Carrozzo, L. Bianchi, A. Delogu, G. Giannelo and G. Nini, 1983. Allergic contact dermatitis of the lip margins from p-tertiarybutylphenol in a lipliner. Contact Dermatol., 28: 146-148.
CrossRef - Arndt, K.A. and T.B. Fitzpatrick, 1965. Topical use of hydroquinone as a depigmenting agent. J. Am. Med. Assoc., 194: 965-967.
Direct Link - Bajaj, A.K., S.C. Gupta and A.K. Chatterji, 1990. Contact depigmentation from free p-tertiary butyl phenol in bindi adhesive. Contact Dermatol., 22: 99-102.
CrossRef - Bajaj, A.K., A. Misra, K. Misra and S. Rastogi, 2000. The azo dye solvent yellow 3 produces depigmentation. Contact Dermatitis, 42: 237-237.
Direct Link - Bajaj, A.K., S.K. Dubey and K. Misra, 2004. A simple spectroscopic technique proposed for scanning the depigmenting potential of azo dyes. Contact Dermatitis, 51: 97-98.
CrossRefDirect Link - Dogliotti, M., I. Caro, R.G. Hartdegen and D.A. Whiting, 1974. Leucomelanoderma in Blacks a recent epidemic. S. Afr. Med. J., 48: 1555-1558.
PubMedDirect Link - Dupre, A., J.P. Ortonne, R. Vibraben and F. Arfeux, 1985. Chloroquin induced hypopigmentation of hair and freckles association with congenital renal failure. Arch. Dermatol., 121: 1164-1166.
Direct Link - Edwards, E.A. and S.Q. Duntley, 1939. The pigment and color of living human skin. Am. J. Anat., 65: 1-33.
CrossRef - Fitzpatrick, T.B., M.A. Pathak and T.A. Magnus, 1963. Abnormal reactions of man to light. Ann. Rev. Med., 14: 195-214.
CrossRef - Jimbow, K., H. Obata, M.A. Pathak and T. B. Fitzpatrick, 1974. Mechanism of depigmentation by hydroquinone. J. Invest. Dermatol., 62: 436-449.
Direct Link - Kawashima, T., K. Yonemoto, G.A. Gellin and K. Fukuyama, 1984. Effect of 4-tertiarybutyl catechol on glutathoine-metabolizing enzymes in vivo and in vitro. J. Invest. Dermatol., 82: 53-56.
Direct Link - Kahn, G., 1970. Depigmentation caused by phenolic detergent germicides. Arch. Dermatol., 102: 177-187.
CrossRefDirect Link - Kligman, A.M. and I. Willis, 1975. A new formula for depigmenting human skin. Arch. Dermatol., 111: 40-48.
CrossRefDirect Link - Lerner, A.B. and T.B. Fitzpatrick, 1953. Treatment of melanin hyperpigmentation. J. Am. Med. Assoc., 152: 577-582.
CrossRefDirect Link - Malten, K.E., E. Seutter and I. Hara, 1971. Occupational vitiligo due to paratertiary butylphenol and homologues. Trans. St. John's Hospital Dermatol. Soc., 57: 115-134.
PubMedDirect Link - Mausur, J.D.S., K. Fukuyama, G.A. Gellin and W.L. Epstein, 1978. Effects of 4-tertiary butyl catechol on tissue cultured melanocytes. J. Invest. Dermatol., 70: 275-279.
CrossRefDirect Link - McCarthy, L.E.E. and L.K. McCafferty, 1925. Leukoderma following dermatitis venenata (Rhus toxicodendron). Arch. Dermatol. Syphilol., 12: 356-359.
CrossRefDirect Link - McCormac, K.P.C., G.N. Ledesma and J.G. Vaillant, 1984. Linear hypopigmentation after intra-articular corticosteroid injection. Arch. Dermatol., 120: 708-709.
CrossRefDirect Link - Mcguvie, J. and J. Hendee, 1971. Biochemical basis for depigmentation of skin by phenolic germicides. J. Invest. Dermatol., 57: 256-261.
CrossRefDirect Link - Nakayama, H., R. Harado and M. Toda, 1976. Pigmented cosmetic dermatitis. Int. J. Dermatol., 15: 673-675.
CrossRefDirect Link - Nishimura, M., G.A. Gellin, S. Hoshino, J.H. Epstein, W.L. Epstein and K. Fukuyama, 1982. Ultrastructural demonstration of chemical modification of melanogenesis in hairless mouse skin. Anal. Rec., 202: 193-201.
CrossRefDirect Link - Seiji, M. and S. Iwashita, 1963. The site of melanin formation in melanocytes. J. Biochem., 54: 103-106.
PubMed - Taylor, J.S., H.I. Maibach, A.A. Fisher and W.F. Bergfeld, 1993. Contact leucoderma associated with the use of hair colors. Cutis, 52: 273-280.
PubMed