
Omar M.E. Abdel-Salam
Department of Pharmacology, National Research Centre, Tahrir St., Dokki, Cairo, Egypt
Amany Ameen Sleem
Department of Pharmacology, National Research Centre, Tahrir St., Dokki, Cairo, Egypt
Nabila S. Hassan
Department of Pathology, National Research Centre, Tahrir St., Dokki, Cairo, Egypt
Hafiza A Sharaf
Department of Pathology, National Research Centre, Tahrir St., Dokki, Cairo, Egypt
Mozsik Gy
Department of Internal Medicine, Medical University ofPecs, Hungary
Journal of Pharmacology and Toxicology
Year: 2006 | Volume: 1 | Issue: 2 | Page No.: 147-156







ABSTRACT
The effect of capsaicin, the pungent principle of red hot pepper and a sensory stimulant on the development of hepatic injury in rats treated with carbon tetrachloride was investigated. CCl4 was given orally (2 mL kg-1 followed by 1 mL kg-1 after one week). Capsaicin at three dose levels (10, 100 and 1000 μg kg-1; 2.5, 25 and 250 μg mL-1) or silymarin (22 mg kg-1) was administered orally daily for 10 days, starting at time of administration of CCl4. The daily administration of capsaicin conferred significant protection against the hepatotoxic effects of CCl4 in rats. It decreased the increases in serum alanine aminotransferase (ALT) and aspartate aminotransferase (AST) and also prevented the development of histological hepatic necrosis caused by CCl4 as determined 10 days after drug administration. Thus, compared with the CCl4 control group, serum ALT decreased by 39.3, 59.3 and 71.1%, while AST decreased by 14.3, 21.5 and 23.3%, after the administration of capsaicin at the aforementioned doses, respectively. Serum bilirubin was decreased by capsaicin at 10 or 100 μg kg-1 (46.4 and 66.5% reduction, respectively), but an increased bilirubin and ALP was observed after the highest dose of capsaicin. Meanwhile, silymarin reduced serum ALT by 65.3%, AST by 18.9%, ALP by 22% and bilirubin by 13.4%, compared to CCl4 control. Serum proteins were significantly increased by 16.9-22.9% after treatment with capsaicin compared, whilst marked increased in serum glucose by 66.9% was observed after the highest dose of capsaicin compared with vehicle-treated group. Quantitative analysis of the area of damage by image analysis technique showed a reduced area of damage from 13.6% to 7.5, 4.3 and 2.8% by the aforementioned doses of capsaicin, respectively and H and E staining also showed markedly less hepatic necrosis in rats treated with capsaicin or silymarin. Histochemical alterations such as decreased nuclear DNA, cell glycogen and protein contents caused by CCl4 in hepatocytes were prevented by capsaicin as well as by silymarin. It is concluded that orally administered capsaicin exerts beneficial effects on liver histopathologic changes and enzymatic release caused by CCl4 in rats, but high doses are likely to result in cholestasis.
PDF Abstract XML References
How to cite this article
Omar M.E. Abdel-Salam, Amany Ameen Sleem, Nabila S. Hassan, Hafiza A Sharaf and Mozsik Gy, 2006. Capsaicin Ameliorates Hepatic Injury Caused by Carbon Tetrachloride in the Rat. Journal of Pharmacology and Toxicology, 1: 147-156.
DOI: 10.3923/jpt.2006.147.156
URL: https://scialert.net/abstract/?doi=jpt.2006.147.156
DOI: 10.3923/jpt.2006.147.156
URL: https://scialert.net/abstract/?doi=jpt.2006.147.156
INTRODUCTION
Capsaicin (8-methyl-N-vanillyl-6-nonenamide), the pungent principle of red hot pepper excites and in high doses desensitizes a subset of mammalian primary afferents with C and Aδ thin fibers. Owing to the remarkable neuroselective site of action of the drug, it has been considered invaluable probe for delineating neural responses mediated by this subgroup of afferents (Szolcsányi, 1977, 1984, 1993; Holzer, 1991). In the rat stomach, capsaicin introduced at small doses and low concentrations, which excited the peripheral terminals of capsaicin-sensitive sensory nerves, resulted in mucosal protection. Higher desensitizing concentrations of the agent, impaired mucosal defense, with the resultant aggravation of gastric mucosal damage (Szolcsányi and Barthó, 1980; Holzer et al., 1989; Abdel-Salam et al., 1995). In addition, defunctionalization of capsaicin-sensitive sensory nerves with systemic treatment of rats with high doses of capsaicin resulted in enhancement of gastric mucosal damage evoked by several noxious agents such as ethanol (Holzer and Lippe, 1988), cysteamine (Holzer and Sametz, 1986) or indomethaicin (Gray et al., 1994). The release of vasodilator mediators from sensory nerve endings by capsaicin accounted for the protection by low concentrations of capsaicin, whereas failure of peptide release, impaired one of the most important mucosal defenses, that is the increase in gastric blood flow in face of noxious challenge, with the resultant aggravation of gastric mucosal damage upon exposure to acid (Szolcsányi and Barthó, 1980; 2001; Holzer, 1991).
The mucosal protective effect of sensory stimulant doses of capsaicin is not confined to the stomach, but also is observed in other organs such as the colon (Evangelista and Meli, 1989) and pancreas (Dembinski et al., 2003). Meanwhile and despite a surge of publications dealing with the effect of the agent in the stomach, fewer studies have investigated the effect of capsaicin on liver injury. As early as 1962, Lee, found that rabbits fed diet that contained high levels of ground red peppers developed liver cirrhosis, as well as gastric ulcers. In more recent studies, in which ablation of capsaicin sensitive sensory nerves by systemic capsaicin pretreatment (capsaicin desensitization) was used as a tool to investigate the role of capsaicin-sensitive sensory nerves, decreased hepatic fibrosis was reported in rats subjected to bile-duct ligation (Casini et al., 1990). In addition, depletion of primary afferent nerve fibers by neonatal capsaicin treatment prevented immune-mediated liver injury in mice (Bang et al., 2003). In contrast, the attenuation of CCl4-induced acute liver injury by central administration of thyrotropin-releasing hormone depended on intact capsaicin-sensitive afferent neurons and CGRP release (Yoneda et al., 2003). Furthermore, administration of capsaicin and CGRP ameliorated liver injury caused by ischaemia/reperfusion in rats (Harada et al., 2002).
Capsaicin is a popular food ingredient and highly seasoned food is consumed in many parts of the world. Capsaicin can be effectively absorbed after intragastric administration so as to reach hepatobiliary tree at levels sufficient to excite the peripheral terminals of capsaicin-sensitive nerves (Abdel-Salam et al., 2005). Therefore, it looked pertinent to test the effect of oral capsaicin administration upon the development of liver cell injury caused by CCl4 in rats. In the present study, capsaicin was given in three different concentrations of 10, 100 or 1000 μg mL-1. The concentrations used were based upon previous studies in the stomach and bile.
Materials and Methods
Animals
Adult Sprague-Dawley rats of either sex, weighing 120-130 g of body weight were used throughout the experiments and fed with standard laboratory chow and water ad libitum. The study was done at the Departments of Pharmacology and Pathology, National Research Centre, Cairo, between January and March 2005. All animal procedures were performed according to approved protocols and in accordance with the recommendations for the proper care and use of laboratory animals.
Drugs and Chemicals
Carbon tertrachloride (BDH Chemicals, England), Silymarin (Sedico Pharmaceutical Co. Cairo, A.R.E.) and capsaicin (Sigma, USA) were used in the experiments. Stock solutions of capsaicin (10 mg mL-1) contained 10% ethanol, 10% Tween 80, 80% saline solution.
The Carbon Tetrachloride Model of Hepatic Damage
Different groups of rats (n = 7-8 per group) were used. Hepatotoxicity was induced by treating rats with CCl4-olive oil, given orally (1:1, 2 mL kg followed by 1 mL kg after one week). Control rats were treated with olive oil. Capsaicin at three dose levels (10, 100 and 1000 μg kg-1; 2.5, 25 and 250 μg mL-1; 0.5 mL rat-1) or silymarin (22 mg kg-1) were administered orally daily for 10 days, starting at time of administration of CCl4. Rats had free access to food and drinking water during the study.
Biochemical Assessment
Blood samples were obtained from the retro-orbital vein plexus at the end of the experiment under light ether anesthesia. Alanine aminotransferase (ALT) and aspartate aminotransferase (AST) activities in serum were measured according to Reitman-Frankel colorimetric transaminase procedure (Crowley, 1967), whereas colorimetric determination of alkaline phosphatase (ALP) activity was done according to the method of Belfield and Goldberg (1971), using commercially available kits (BioMérieux, France). Total proteins in serum were measured spectrophotometrically (Bradford, 1976). Glucose concentrations in serum were measured enzymatically (Bauer, 1982).
Histopathological and Histochemical Studies
At the end of the treatment period, rats were killed; livers were excised and fixed in 10% formalin saline, Bouin’s and Carnoy’s fluids. Sections were prepared and stained with hematoxylin and eosin-stained (H and E) for the histological investigations. Periodic acid-Schiff’s, mercury and bromophenol and Feulgen techniques were used for intracellular glycogen, proteins and nuclear DNA determinations. Further histopathgological evaluation was done with morphometry. The percentage of necrotic areas surrounded by fatty degeneration was measured by a computerized image analyzer. Qwin Leica image processing and analysis system (Cambridge-England) was used for interactive automatic measurement of the percentage of damaged areas on slides stained by H and E by analyzing 15 random fields per slide.
Statistical Analysis
All results are expressed as means±SE. Comparison between two independent groups was made by Mann-Whitney U-test. Comparison of the values before and after CCl4 was made by paired Student's t-test. Multiple group comparisons were performed by ANOVA followed by Duncan test. p<0.05 was considered statistically significant.
RESULTS
Biochemical Results
In the CCl4-induced acute liver injury, the levels of alanine aminotransferase (ALT) and aspartate aminotransferase (AST) activities were markedly raised in plasma indicating the severity of hepatic injury caused by CCl4. The serum ALT and AST levels were significantly elevated from 8.8±0.7 and 47.7±4.2 (olive oil) to 30±2.9 and 114.0±6.7 IU L-1, respectively (p<0.01) (Table 1).
| Table 1: | Effect of capsaicin or silymarin on serum alanine aminotransferase (ALT), aspartate aminotransferase (AST), alkaline phosphatase (ALP), total bilirubin, total proteins, glucose and area of damage in CCl4-treated rats |
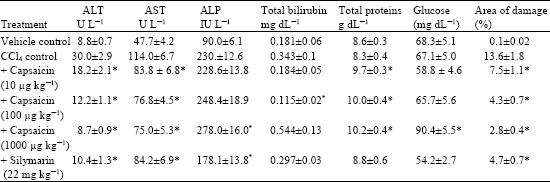 | |
| Results are means±SE. Data were analyzed by one way ANOVA and means of different groups were compared by Duncan’s multiple range test. Two-tailed probabilities of less than 0.05 were considered significant. *p<0.05 compared with the CCl4 control group | |
Alkaline phosphatase (ALP) activity was also raised in serum of CCl4-treated rats, suggesting congestion and cholestasis (90.0±6.1 (olive oil) to 230±12.7 IU L-1). Serum bilirubin increased from 0.18±0.06 (olive oil) to 0.34±0.1 mg dL). The administration of capsaicin at doses of 10, 100 and 1000 μg kg-1 caused a dose-dependent reduction in serum ALT and AST levels, compared with the CCl4 control group. Thus, serum ALT decreased by 39.3, 59.3 and 71.1%, while AST decreased by 14.3, 21.5 and 23.3%, by the above mentioned doses of capsaicin, respectively. Serum bilirubin was reduced by 46.4 and 66.5% after 10 or 100 μg kg-1 of capsaicin. In contrast, an increase in ALP (by 21%; 278±16 vs 230±12.7 IU L) and bilirubin (by 58.8%; 0.54±0.13 vs 0.34±0.1 mg dL) was observed with the higher dose of capsaicin (1000 μg kg-1). Meanwhile, silymarin reduced serum ALT by 65.3%, AST by 18.9%, ALP by 22.5% and bilirubin by 13.4%, compared to CCl4 control group. Serum proteins were significantly increased by 16.9-22.9% after treatment with capsaicin compared with vehicle-treated group. Serum glucose was also markedly increased by 66.9% after the highest dose of capsaicin compared with vehicle-treated group.
Image Analysis
Quantitative analysis of hepatic tissue damage by image analysis technique showed a reduced area of damage from 13.6% to 7.5, 4.3 and 2.8% by the aforementioned doses of capsaicin, respectively (p<0.01) and to 4.7% by silymarin (Table 1).
Histopathological Results
The livers of saline control rats revealed the characteristic hepatic architecture (Fig. 1A). The livers of rats subjected to CCl4 showed fibrosis, dilated blood sinusoids, mononuclear cell infiltration, increase in Kupffer cells and nuclear ploymorphism (Fig. 1B). In rats treated with CCl4 + silymarin, liver sections showed marked diminution of fibrosis as compared to the group treated with CCl4 only (Fig. 1C). The liver sections of rats treated with CCl4 + capsaicin (1000 μg kg-1) showed that the liver tissue restored its structure to almost normal picture (Fig. 1D).
Histochemical Changes
Figure 2 shows the distribution of glycogen contents in liver tissue stained by periodic acid-Schiff’s (PAS) reagent, characterized in normal rats by deeply stained reddish granules in the cytoplasm of hepatocytes (Fig. 2A).
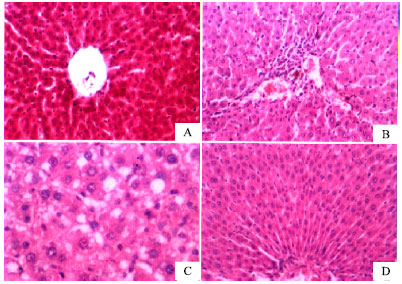 | |
| Fig. 1: | Hematoxylin and eosin stained liver sections from (A) normal saline-treated rat. (B) CCl4 (vehicle-treated) control showing fibrosis, dilated blood sinusoids, mononuclear cell infiltration, increased Kupffer cells and nuclear ploymorphism. © CCl4 + silymarin treatment showing marked diminution of fibrosis. (D) CCl4 + capsaicin (1000 μg kg-1) treatment showing almost preserved architecture |
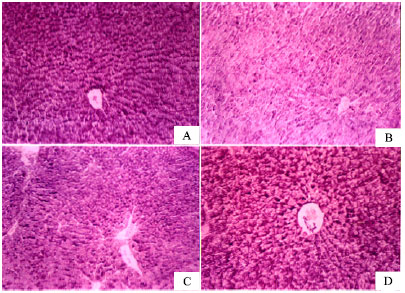 | |
| Fig. 2: | Distribution of glycogen contents in liver tissue stained by Periodic acid-Schiff’s (PAS). (A) Normal distribution of PAS reaction in liver cells from normal saline-treated rat characterized by deeply stained reddish granules in the cytoplasm of hepatocytes. (B) CCl4 (vehicle-treated) control showing sharp decrease in glycogen content. © CCl4 + silymarin treatment showing glycogen content almost restored to normal. (D) CCl4 + capsaicin (1000 μg kg-1)-treated rats, showing marked increase in PAS reaction in the heptacyte cytoplasm |
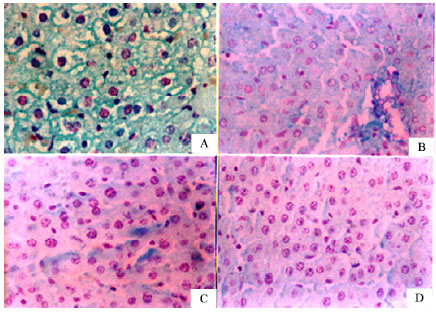 | |
| Fig. 3: | Liver sections stained with Feulgen technique. (A) Normal saline-treated rat; nuclear chromatin substances of the cells were stained red by the Feulgen technique. (B) CCl4 (vehicle-treated) control showing marked decrease in DNA contents in hepatic nuclei. © CCl4 + silymarin treatment and (D) CCl4 + capsaicin (1000 μg kg-1)-treated rats, marked improvement in DNA content was evident compared with CCl4 control rats |
The glycogen content was sharply reduced in CCl4-treated control rats (Fig. 2B), indicating reduced glycogen synthesis and glycogen stores. After treatment with silymarin, glycogen content was almost restored to normal (Fig. 2C). In CCl4 + capsaicin (1000 μg kg-1)-treated rats, marked increase in PAS reaction in the hepatocyte cytoplasm was seen in Fig. 2D.
Figure 3 shows the histological appearance of liver sections stained with Feulgen technique. The histochemical investigation of liver in the control rats showed that nuclear chromatin substances of the cells were stained red by the Feulgen technique. The DNA containing bodies were moderately dense in the nuclei (Fig. 3A). The histochemical investigation of DNA in CCl4-treated control rats revealed significant decrease in DNA contents in hepatic nuclei (Fig. 3B). In liver sections of rats treated with silymarin (Fig. 3C) or capsaicin (1000 μg kg-1) (Fig. 3D), marked improvement in DNA content was evident compared with CCl4 control rats.
DISCUSSION
The present study provides the first evidence that oral administration of capsaicin reduces hepatic injury caused by the hepatotoxin CCl4 in rats. Liver enzymes (alanine transaminase, aspartate transaminase) in the serum and liver necrosis evaluated histologically were significantly reduced in the capsaicin-treated rats. The CCl4-related disturbances of cellular dysfunction indicated by reduced glycogen and protein content and DNA alterations were ameliorated by capsaicin. The significance of these observations derives from the fact that capsaicin is a popular food ingredient and is consumed worldwide and thus its effect on the liver, the largest metabolic organ and on metabolism is of utmost importance. A number of studies have examined the effects of capsaicin on the hepatic architecture. In these studies where high and possibly desensitizing doses of capsaicin or neonatal capsaicin treatment were employed, the authors reported increased (Lee, 1963) or decreased hepatic fibrosis (Casini et al., 1990; Bang et al., 2003), or failure to develop hyperkinetic circulation when cirrhosis or portal hypertension was induced (Lee and Sharkey, 1993; Li et al., 2003) or even complete inhibition of stress-induced cellular damage (Erin et al., 2000). In clear contrast to the above findings, administration of capsaicin and CGRP ameliorated liver injury caused by ischaemia/reperfusion in rats (Harada et al., 2002), while capsaicin-sensitive afferent appear to be involved in the mechanism by which central administration of TRH attenuates the CCl4-induced acute liver injury (Yoneda et al., 2003). This discrepancy in results is probably due to the different regimens concerning capsaicin desensitization. It is also worth mentioning that after neonatal treatment during development, secondary changes in the periphery and in the nervous system result in a qualitatively different situation as compared to adult treated animals (Szolcsányi, 1993; Szoke et al., 2002).
Capsaicin is widely used as a probe for sensory neuron mechanisms. At low doses (in the μg kg range) or concentrations (in the μg kg-1 range), capsaicin exerts a powerful excitatory effect on peripheral sensory nerve endings, this effect being confined to unmyelinated afferents (C-fibers) (Szolcsányi 1977, 1984; Holzer, 1988; Mózsik et al., 2001). Capsaicin sensitive afferent nerve endings are not only sensory receptors for initiation of nerve impulses, but also effector sites, where the mediators are released and which in turn elicit local tissue responses (Szolcsányi 1984, 1990; Holzer, 1988). Capsaicin evokes the release of several neuropeptides from capsaicin-sensitive nerve endings such as calcitonin gene-related peptide (CGRP), somatostatin and neurokinins (Holzer et al., 1990). The former is a potent vasodilator and much of the observed protection by capsaicin in the stomach has been suggested to be mediated by CGRP (Evangelista, 1994). Capsaicin introduced into the rat stomach (Lippe et al., 1989; Holzer et al., 199l), or applied on the serosal surface of the stomach or jejunum (Abdel-Salam et al., 1996), led to a marked increase in gastric mucosal blood flow, while submucosal application of the agent caused dilatation of submucosal arterioles (Chen et al., 1992). In peripheral blood vessels, the application of capsaicin produces a sustained vasodilatation, which is mediated by the release of neuropeptides from sensory nerve endings (Fujimori et al., 1990).
The rat hepatobiliary tract is densely innervated by CGRP-containing fibers form dense networks in the fibromuscular layer of the biliary tree and surrounding the portal vein. This suggests the involvement of these peptidergic visceral afferents in regulating hepatobiliary activities, including hemodynamic functions of the hepatic vasculature (Goehler and Sternini, 1996). Studies indicated that vasodilatation in the vascular bed of the hepatic artery induced by reduction of the portal inflow to the liver by 90% is partially mediated by activation of capsaicin-sensitive sensory fibers in the liver, probably due to local tissue ischemia and hypoxia (Biernat et al., 2005). In rats subjected to hepatic I/R, antithrombin increased hepatic tissue levels of 6-keto-PGF1alpha, hepatic-tissue blood flow and attenuated both hepatic local inflammatory responses and liver injury by a mechanism dependent upon intact sensory innervation, where CGRP-induced activation of both endothelial nitric oxide synthase and cyclooxygenase-1 might be critically involved (Harada et al., 2005). Capsaicin is likely to ameliorate hepatic injury through increasing/maintaining microvascular perfusion. Early vascular events were suggested to have a role in liver damage caused by CCl4 (Tanaka et al., 1999) and increased hepatic arterial blood flow may act to lessen CCl4-induced acute hepatic injury (Tanaka et al., 1999; Hsu et al., 1993). Capsaicinoids are readily absorbed from gut and intact molecules are found in portal vein blood (Srinivasan et al., 1980; Kawada et al., 1984; Kawada and Iwai, 1985; Donnerer et al., 1990). Capsaicin administered by intragastric or intraduodenal routes (4-400 μg mL-1; 10-1000 μg kg-1) altered bile flow and was readily excreted into bile at concentrations that are likely to have excitatory effects on sensory nerve endings (Abdel-Salam et al., 2005). Findings in the present study, indicates that orally administered capsaicin exerts beneficial effects on liver histopathologic changes and enzymatic release caused by CCl4 in rats.
REFERENCES
- Belfield, A. and D.M. Goldberg, 1971. Revised assay for serum phenyl phosphatase activity using 4-amino-antipyrine. Enzyme, 12: 561-573.
PubMedDirect Link - Biernat, J., W.W. Pawlik, R. Sendur, A. Dembinski, T. Brzozowski and S.J. Konturek, 2005. Role of afferent nerves and sensory peptides in the mediation of hepatic artery buffer response. J. Physiol. Pharmacol., 56: 133-145.
Direct Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Casini, A., I.T. Lippe, S. Evangelista, P. Geppetti and P. Santicioli et al., 1990. Effect of sensory denervation with capsaicin on liver fibrosis induced by common bile duct ligation in rat. J. Hepatol., 11: 302-312.
PubMed - Dembinski, A., Z. Warzecha, P. Ceranowicz, J. Jaworek and R. Sendur et al., 2003. Stimulation of sensory nerves and CGRP attenuate pancreatic damage in ischemia/reperfusion induced pancreatitis. Med. Sci. Monit., 9: 418-425.
Direct Link - Donnerer, J., R. amann, R. Schuligoi and F. Lembeck, 1990. Absorption and metabolism of capsaicinoids following intragastric administration in rats. Naunym-Schmiedeberg's Arch. Pharamcol., 342: 357-361.
CrossRef - Erin, N., F. Ercan, B.C. Yegen, S. Arbak, I. Okar and S. Oktay, 2000. Role of capsaicin-sensitive nerves in gastric and hepatic injury induced by cold-restraint stress. Dig. Dis. Sci., 45: 1889-1899.
Direct Link - Fujimori, A., A. Saito, S. Kimura and K. Goto, 1990. . Release of calcitonin gene-related peptide (CGRP) from capsaicin-sensitive vasodilator neurons in the rat mesenteric artery. Neurosci. Lett., 112: 173-178.
PubMed - Goehler, L.E. and C. Sternini, 1996. Calcitonin gene-related peptide innervation of the rat hepatobiliary system. Peptides, 17: 209-217.
PubMed - Gray, J.L., N.W. Bunnett, S.L. Orloff, S.J. Mulvihill and H.T. Debas, 1994. Role for calcitonin gene-related peptide in protection against gastric ulceration. Ann. Surg., 219: 58-64.
PubMedDirect Link - Harada, N., K. Okajima, M. Uchiba and T. Katsuragi, 2002. Ischemia/reperfusion-induced increase in the hepatic level of prostacyclin is mainly mediated by activation of capsaicin-sensitive sensory neurons in rats. J. Lab. Clin. Med., 139: 218-226.
Direct Link - Harada, N., K. Okajima, M. Yuksel and H. Isobe, 2005. Contribution of capsaicin-sensitive sensory neurons to antithrombin-induced reduction of ischemia reperfusion-induced liver injury in rats. Thromb. Haemost., 93: 48-56.
Direct Link - Holzer, P. and W. Sametz, 1986. Gastric mucosal protection against ulcerogenic factors in the rat mediated by capsaicin-sensitive afferent neurons. Gastroenterology, 91: 975-981.
PubMed - Holzer, P., 1988. Local effector functions of capsaicin sensitive sensory nerve endings involvement of tachykinins calcitonin gene related peptide and other neuropeptides. Neuroscience, 42: 739-768.
PubMed - Holzer, P. and I.T. Lippe, 1988. Stimulation of afferent nerve endings by intragastric capsaicin protects against ethanol induced damage of gastric mucosa. Neuroscience, 27: 981-987.
PubMed - Holzer, P., M.A. Pabst and I.T. Lippe, 1989. Intragastric capsaicin protects against aspirin induced lesion formation and bleeding in the rat gastric mucosa. Gastroenterology, 96: 1425-1433.
PubMed - Holzer, P., B.M. Peskar, B.A. Peskar and R. Amann, 1990. Release of calcitonin gene-related peptide induced by capsaicin in the vascularly perfused rat stomach. Neurosci. Lett., 108: 195-200.
Direct Link - Holzer, P., 1991. Capsaicin: Cellular targets, mechanisms of action and selectivity for thin sensory neurons. Pharmacol. Rev., 43: 144-201.
Direct Link - Holzer, P., E.H. Livingston, A. Saria and P.H. Guth, 1991. Sensory neurons mediate protective vasodilatation in rat gastric mucosa. Am. J. Physiol., 260: 363-370.
Direct Link - Hsu, C.T., K. Schichijo, M. Ito and I. Sekine, 1993. The effect of chemical sympathectomy on acute liver injury induced by carbon tetrachloride in spontaneously hypertensive rats. J. Auton Nerv Syst., 43: 91-96.
Direct Link - Kawada, T., T. Suzuki, M. Takahashi and K. Iwai, 1984. Gastrointestinal absorption and metabolism of capsaicin and dihydrocapsaicin in rats. Toxicol. Applied Pharmacol., 72: 449-456.
CrossRefDirect Link - Kawada, T. and K. Iwai, 1985. In vivo and in vitro metabolism of dihydrocapsaicin, a pungent principle of hot pepper in rats. Agric. Biol. Chem., 49: 441-448.
Direct Link - Lee, S.O., 1963. [Studies on the influence of diets and lipotropic substances upon the various organs and metabolic changes in rabbits on long-term feeding with red pepper. II. Histopathological changes of liver and spleen]. Korean J. Int. Med., 6: 383-400.
PubMed - Lee, S.S. and K.A. Sharkey, 1993. Capsaicin treatment blocks development of hyperkinetic circulation in portal hypertensive and cirrhotic rats. Am. J. Physiol., 264: 868-873.
Direct Link - Li, Y., D. Song, Y. Zhang and S.S. Lee, 2003. Effect of neonatal capsaicin treatment on haemodynamics and renal function in cirrhotic rats. Gut, 52: 293-299.
CrossRef - Lippe, I.T., M.A. Pabst and P. Holzer, 1989. Intragastric capsaicin enhances rat gastric elimination and mucosal blood flow by afferent nerve stimulation. Br. J. Pharmacol., 96: 91-100.
PubMed - Mozsik, G., A. Vincze and J. Szolcsanyi, 2001. Four response stages of capsaicin-sensitive primary afferent neurons to capsaicin and its analog: Gastric acid secretion, gastric mucosal damage and protection. J. Gastroenterol. Hepatol., 16: 1093-1097.
Direct Link - Szoke, E., L. Seress and J. Szolcsanyi, 2002. Neonatal capsaicin treatment results in prolonged mitochondrial damage and delayed cell death of B cells in the rat trigeminal ganglia. Neuroscience, 113: 925-937.
Direct Link - Szolcsanyi, J., 1977. A pharmacological approach to elucidation of the role of different nerve fibers and receptor endings in mediation of pain. J. Physiol., 73: 251-259.
PubMed - Szolcsanyi, J. and L. Bartho, 1980. Impaired Defense Mechanism to Peptic Ulcer in the Capsaicin Desensitized Rat. In: Advances in Physiological Sciences. Gastrointestinal Defense Mechanisms, Mozsik, G.Y., O. Hanninen and T. Javor (Eds.). Pergamon Press and Akademiai Kiado, Oxford and Budapest, pp: 39-51.
- Szolcsanyi, J. and L. Bartho, 2001. Capsaicin-sensitive afferents and their role in gastroprotection: An update. J. Physiol. Paris, 95: 181-188.
CrossRef - Tanaka, N., K. Tanaka, Y. Nagashima, M. Kondo and H. Sekihara, 1999. Nitric oxide increases hepatic arterial blood flow in rats with carbon tetrachloride-induced acute hepatic injury. Gastroenterology, 117: 173-180.
Direct Link - Yoneda, M., Y. Sato, K. Nakamura, S. Yokohama, T. Kono, H. Watanobe and A. Terano, 2003. Involvement of calcitonin gene-related peptide and capsaicin-sensitive afferents in central thyrotropin-releasing hormone-induced hepatic cytoprotection. Eur. J. Pharmacol., 478: 173-177.
Direct Link