
Mohammad Sayedul Islam
Department of Biochemistry and Molecular Biology, University of Chittagong, Chittagong-4331, Bangladesh
LiveDNA: 880.35589
Md Saiful Islam
Department of Biochemistry and Molecular Biology, University of Chittagong, Chittagong-4331, Bangladesh
Journal of Plant Sciences
Year: 2021 | Volume: 16 | Issue: 1 | Page No.: 10-20







ABSTRACT
Background and Objective: Rice is considered one of the most important cereal crops for human nutrition. Based on current predictions of the increasing global population, rice production needs to be increased by more than 50% to meet the future demand of the rice-eating countries. It has already been demonstrated that cereal crop yield could be increased by modifying root architectural traits for production under limited resources like water and nutrition. The main objective of the study is to assess the rooting behaviour of a panel of Boro and Amon rice cultivars collected from the Bangladesh Rice Research Institute (BRRI). Materials and Methods: Two established screening methods, hydroponic and soil-filled glass rhizotron system were used to assess the panel of Boro and Amon rice cultivars. Using these two methods, deep and shallow-rooted cultivars were identified. A panel of 16 rice cultivars was used in this study. Results: Assessing the 16 cultivars in a hydroponic system, 98% variation was observed by the genotype at day 46 (p = 0.001). Soil filled glass rhizotron was used to assess six of these accessions along with two check varieties revealing strong genotypic variation. Analysis of variance (ANOVA) showed a 73.37% variation explained by the genotype (F = 13.20, p = 0.001) at 46 days after sowing. Conclusion: Assessing all the traits, the rhizotron data revealed that BR 22, BR 60, BR 19 and BR 3 has the deepest rooting systems. The trait information could be useful to breed better cultivars, especially in drought-prone regions.
PDF Abstract XML References Citation
How to cite this article
Mohammad Sayedul Islam and Md Saiful Islam, 2021. Assessing Root System Characteristics in a Subset of Rice Accessions Through Hydroponic Rhizotron Systems. Journal of Plant Sciences, 16: 10-20.
DOI: 10.3923/jps.2021.10.20
URL: https://scialert.net/abstract/?doi=jps.2021.10.20
DOI: 10.3923/jps.2021.10.20
URL: https://scialert.net/abstract/?doi=jps.2021.10.20
INTRODUCTION
Rice is one of the oldest domesticated crop species having fed more than 3.5 billion people1 and offering 20% of their daily calories2. To achieve the expected yield, a better understanding of the root systems will be required. Root systems play a vital role in the uptake of water and nutrients from the soil3. Root morphological characteristics are known to be important in the drought resistance of some rice (Oryza sativa L.) varieties. Several mechanisms of drought resistance are already been exploited by rice varieties and potentially the most promising one is the possessions of deep and thick root systems that allow access to water at depth4.
Root System Architecture (RSA) plays a pivotal role in crop performance, particularly for cultivation under non-optimal water and nutrient supply conditions5. Root System Architecture (RSA) made up of structural features- like root length, spread, number and length of lateral roots, among others, exhibits great plasticity in response to environmental changes and would be critical to developing crops with more efficient roots. With the changing climate and rainfall abrasions, there is a gradual shift in the system of rice cultivation from a traditional transplanted anaerobic to an aerobic system. To achieve sustainable agricultural production, it needs to be well studied of anatomy and root system architecture6.
A structured root system is necessary for favourable plant growth. The reason for adopting many cultivation practices is to improve the root environment like soil pH and the drainage system7. Crop growth, yield and product quality depend on the roots to a large extent8. Absorption of water and nutrients is the principle function of the root and drought is the foremost abiotic stress that affects more than twenty percent of the total rice-producing area in Asia9. Atmospheric water stress has been linked to root system characteristics10. The development of a deep and extensive root system is an adaptive strategy of plants for drought avoidance11. Root characteristics such as thickness, depth of rooting and root length density have been associated with drought avoidance in rice12,13.Root angle distribution, which is regulated by the growth angle of the crown roots, is one of the root morphological traits used to determine the area of soil over which root capture water and nutrients14. For example, deep rooting is useful for extracting more water from the soil and minimizing drought stress under upland and rainfed lowland conditions15,16. In contrast, under waterlogged conditions, shallow rooting can help plants to absorb oxygen on the soil surface and helps avoid hypoxic conditions12. Maximum root depth of rice and root length density increase with time until the flowering stage and after that, it decreases rapidly until grain maturity17. The structural shape of the root varies with soil physics; particle size, soil water status and soil compaction18. Rice root formation also depends upon cultivation methods19. In upland rice which is sewn directly, their roots grow deeper than in transplanted practices used in flooded low land conditions.
Nutrient uptake by roots in the soil is accomplished by root interception, mass flow and diffusion20 where nutrient uptake occurs as a result of root surface contact with the nutrients in the soil. Root morphology (root length and radius) and architecture are the most important characteristics that influence plant resource acquisition21. Increasing root length density increases water uptake ability but gains in uptake afforded by root density are restricted to a critical root length density where all water in the soil is within approximately 1 cm of a root. Critical root length density is higher for nutrients with low movement in the soil, like phosphorus.
Rice having deep rooting potential may help plants to avoid drought-induced stress by extracting water from deep soil layers22. Lafitte et al.23 showed that deep-rooted genotypes have lower susceptibility to drought and improved productivity under aerobic field conditions while genotypes with shallow rooting systems are more susceptible to stress thereby have lower productivity. Kanbar et al.24 showed that maximum root depth, root shoot ratio and root dry weight conferred an advantage to grain yield under water stress. Root characteristics such as thickness, depth of rooting and root length density have been associated with drought avoidance in rice25. Deep rooting is useful for extracting more water from the soil and minimizing drought stress under upland and rainfed lowland conditions26.
To achieve a successful breeding programme, reliable and robust screening techniques is very important. A panel of 205 rice genotypes were screened through a hydroponic system27. Several root traits, root volume and root length were assessed with this technique. A total of 140 recombinant inbred lines were screened through a rhizotron system developed by Price et al.28 and several root traits evaluated for root growth of the longest visible root, several root past at different depths, root thickness and root mass at different depths after 7 weeks growth. To gain a better understanding of the complex root traits, 134 lines from the Bala x Azucena parental population were grown in agar-filled Perspex chambers to identify Quantitative Trait Loci (QTLs) for both Seminal Root Morphology (SRM) and gravitropic response29. In this experiment, a total of four and two main effect QTLs were detected for the SRM and gravitropic response respectively29.
The main objective of this research is to assess the rooting behaviour of a panel of Boro and Amon rice cultivars collected from the Bangladesh Rice Research Institute (BRRI) and identify deep and shallow-rooted cultivars to facilitate the breeding of drought resistance in rice.
MATERIALS AND METHODS
Study area: The hydroponic experiment was conducted on 14 December, 2018 in the laboratory of the Department of Biochemistry and Molecular Biology, University of Chittagong, Bangladesh. The rhizotron experiment was set up in the Biological Faculty glasshouse on 8 April, 2019, University of Chittagong, Bangladesh.
Experimental design of hydroponic screening of root growth of 13 accessions: All 13 accessions collected from Bangladesh Rice Research Institute (BRRI) were tested in the hydroponic set-up described in the Price and Tomos27. Three check varieties Azucena, Black Gora and IR 64 were included with these accessions, which were tested in hydroponics by Shrestha et al.30.
Each screen was used a randomised complete block design with four replications. Diluted domestic bleach (1% sodium hypochlorite (NaClO3) were used for surface sterilization of seeds and then washed with tap water three times. Seeds were then kept for germination at 30°C for three days. Germinated seedlings were placed in a black seedling tray which contains 84 wells with 55 mm deep and 40 mm and 12 mm diameter on the top and bottom respectively. Pieces of plastic mesh (15×5 mm) were used to hold germinated seedlings which were placed above the hole of the well. Each seedling tray was fitted in the 50 L plastic box 56 cm long, 36 cm wide and 29 cm deep. To prevent the entry of the light into the boxes, empty wells were then be covered with aluminium foil paper. The sides of the boxes were covered by black and white plastic paper. Aeration systems were set up in the box. About 50 L of half-strength Yoshida’s Nutrient solution (pH 5.5) were used to fill up the box for the first three weeks and full strength for the final four weeks31. pH was adjusted daily and the nutrient solution was replaced on weekly basis. The experiment was conducted in a controlled environment, the humidity was maintained at 70% and the temperature was 25-28°C throughout the experiment. The supplementary lights 120 μM m–2 sec–1 (PAR) hrs a day were supplied. The maximum root and shoot length of each plant was recorded weekly for five weeks.
Rhizotron screening: From 13 accessions were tested in hydroponic screening, 6 were selected for the rhizotron screening. Among six accessions four were Boro cultivars (BR 3, BR14, BR 19, BRRI 60) and two were Amon cultivars (BR 22 and BR 32). Two check varieties Azucena and IR 64 were included with these accessions.
A set of 32 glass-fronted rhizotrons was made as described by Price et al.4. Two sheets of 4 mm thick glass 120 cm long and 30 cm wide were used to make up each rhizotron. To hold the two sheets 15 mm apart two wooden strips and a coca-cola bottle cap were used. Duct tape was used to seal the base and two sides of the glass rhizotron. After that wooden strips were removed. Then the rhizotron was filled with 7 kg of soil which gave a total weight of 13 kg of each rhizotron. The rhizotron was grouped in a stack and leaned at an angle of 15 from vertical to encourage the root growth down the lower surface of the glass rhizotron. Polystyrene insulation board were used in front of the stacks to keep out light and maintain temperature.
Each chamber of rhizotron was saturated with full strength Yoshida’s nutrient solution pH5.5 before sowing the seeds. Seeds were sown directly on rhizotron, after seedlings germination, the seedlings were thinned to one plant. Drip irrigation systems were connected to each rhizotron described as Price et al.4. Each rhizotron received last irrigation on the 45 DAS and watering was stopped until harvested on day 52.
Traits measured from rhizotron: Shoot growth was monitored as shoot length while the length of the longest visible root length was recorded on weekly basis until 52 DAS. On day 21 and 42 the angle of spread of the root system was measured with a protractor described as Price et al.4. On the evening of day 42, a digital image was taken of each rhizotron using 12 Megapixel cameras. At the end of experiment day 52, plant shoot was removed in one day, oven-dried at 80 for three days and dry weight was recorded. Over the week, rhizotron was opened, short sections of three of the thickest root are removed from each root system from the base of the root, gently washed with water, oven-dried and weighed.
Image analysis: The number of the main axes of nine angle classes representing each of the 100 subdivision of the protractor was counted. Roots in the division 0-100 and 170-1800 are counted together in the 0-100 class to give the number of roots in the most horizontal of the nine angle classes. A weighted average of the angle was calculated by multiplying the number of roots in each class by the halfway angle of each class (e.g., 5 for the 0-10° class and 35 for the 30-40° class), summing across all classes and dividing that by the total number of axes.
Statistical analysis: Minitab version 17 was used to analyse the data. For both Experiment (Hydroponic and rhizotron screening) data were checked for block effect by analysis of variance (ANOVA) and corrected by taking the residual for further analysis if required. In each screen, each trait one way ANOVA was used to assess the existence of genotypic difference. Genotype means were used to observe the correlation between traits and screens.
RESULTS AND DISCUSSION
Hydroponic screening: The mean root length and shoot length increased linearly with time. Plants had a mean root length of 8.0 cm at day 26 which increased up to 9.0 cm at day 37 and up to 15 cm at day 52. Shoot growth showed a greater increase from 17 cm at days 26-21 cm at day 37 and increased up to 30 cm at day 52. All shoot and root length data collected in hydroponics screening were statistically analyzed for cultivars difference by one way ANOVA.
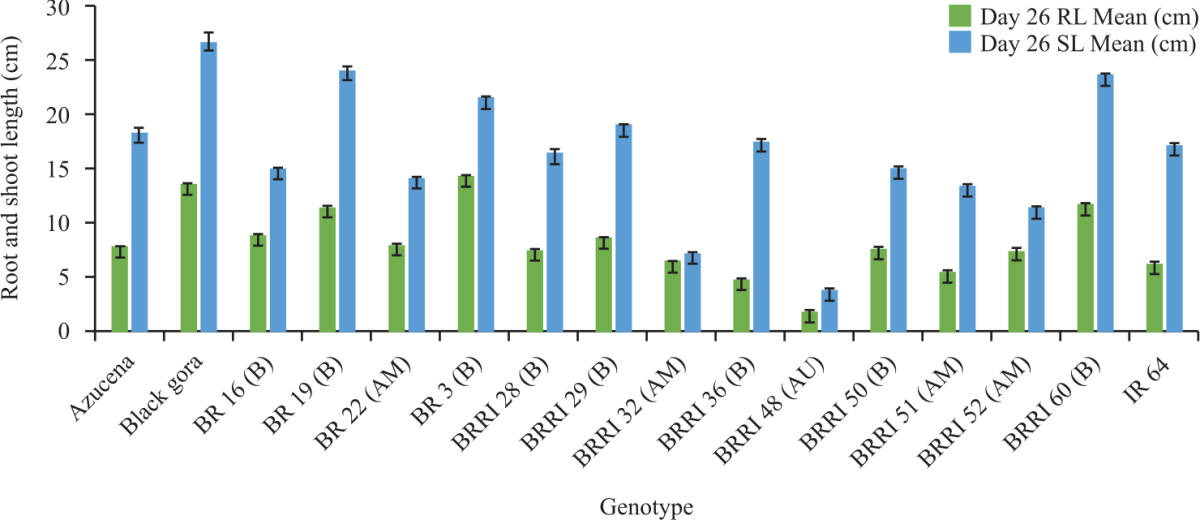
A total of 16 cultivars tested in the hydroponics showed a great difference in root length and shoot length throughout the experiment. The maximum root length and shoot length of all genotypes at days 26 is shown in Fig. 1.
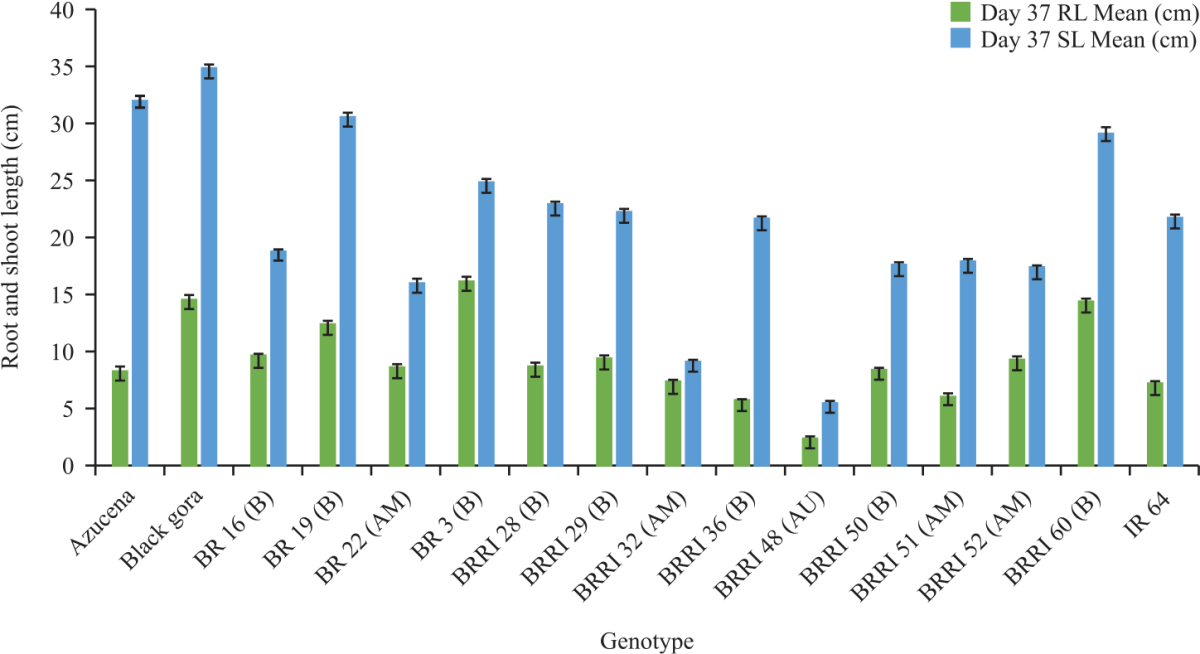
In the root length, significant variation was observed at all days of measurement and the proportion of variation explained by the genotypes was increased from day 26-52 where it was highest at day 37. That means the highest discrimination of root length was observed on day 37. Azucena, Black Gora, BR 3, BR 19, BR 16, BRRI 60 had the longest root on that day while IR64, BRRI 51, BRRI 36, had the shortest (Fig. 2).
The maximum root length of days 37 ranges from 14-16 cm and Black Gora and BR3 were the highest and BRRI 36 and IR 64 were the lowest (Fig. 2) and at that time 58.27% variation explained by genotype (F = 5.27, p = 0.001).
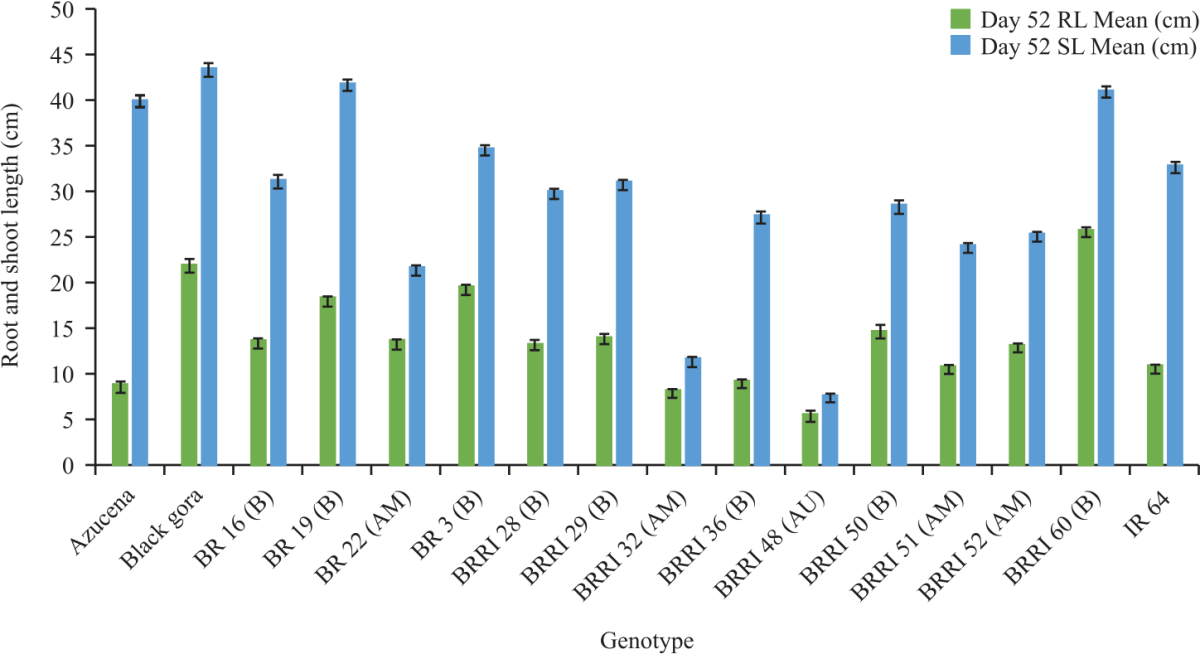
The data of root length and shoot length were recorded at 52 days of hydroponic experiment which showed significant result (p = 0.001). Black Gora, BRRI 60 and BR 3 had the longest root length and IR 64, BRRI 51 and BRRI 36 had the shortest root length (Fig. 3).
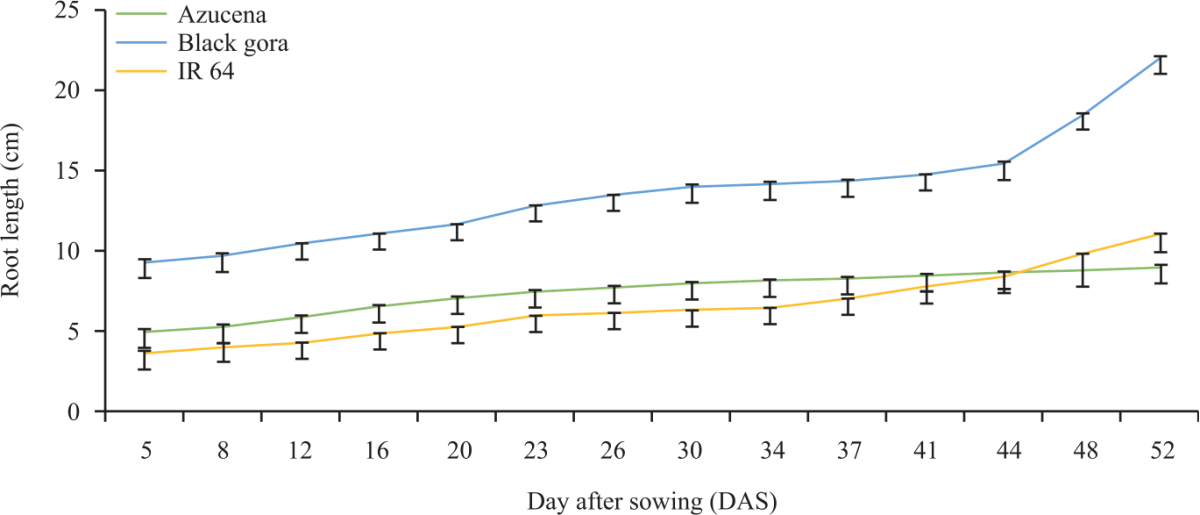
The maximum visible root and shoot length of the check varieties (Fig. 4 and 5) shows that Black Gora and Azucena had long root and shoot length and IR 64 had shallow rooted which follow the same pattern as previously detected in Al-Shugeairy et al.32.
Rhizotron screening: The mean values of all traits measured on the rhizotrons are presented in Table 1, which also summarises the one way ANOVA. All traits were highly significantly different between cultivars with cultivars generally explaining the majority of the variation.
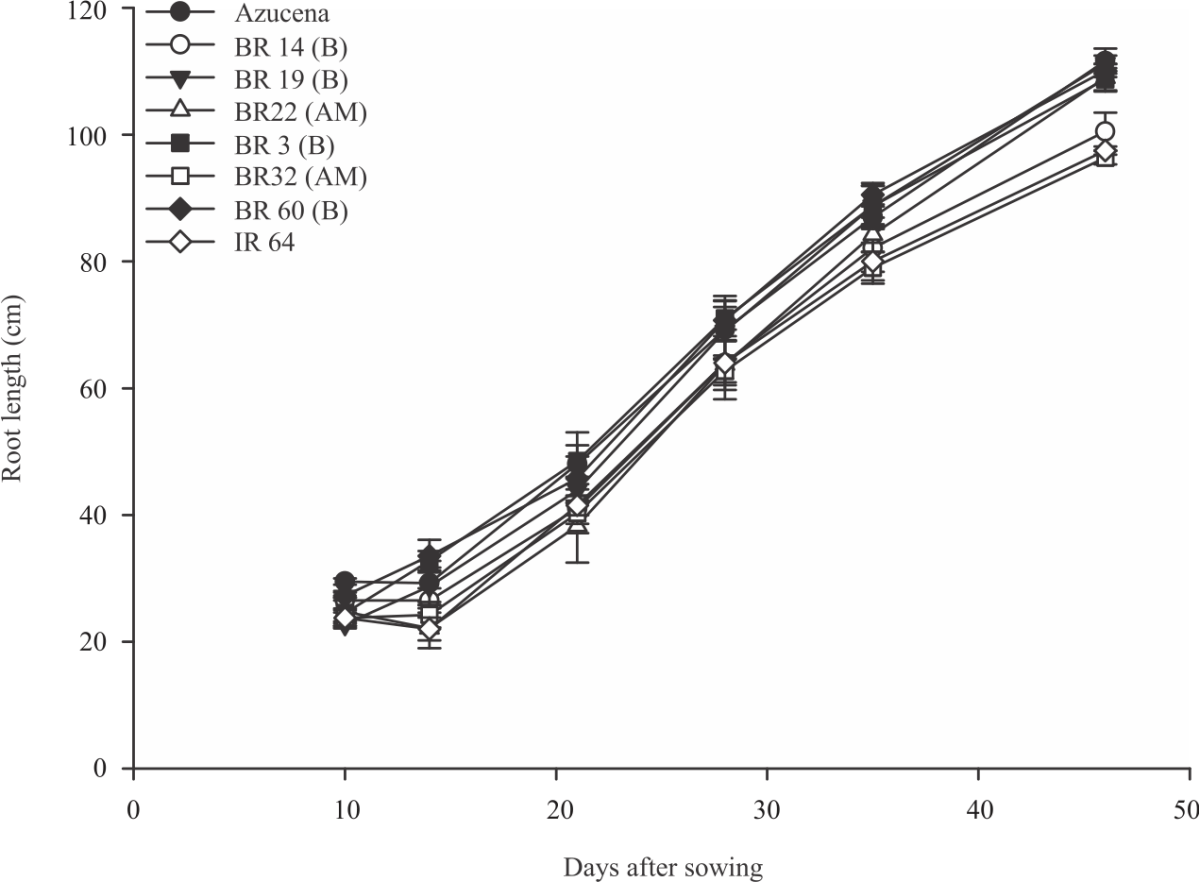
The maximum visible root length measured in the rhizotron increased 109 cm on day 33-118 cm on day 45 (Fig. 6). The mean shoot length increased from 22-74 cm from day 14-45 respectively (Fig. 7). Among the accessions Azucena, BR14, BR32 had the highest shoot length where BRRI 60, BR 3 and IR 64 showed the shortest shoot length (Fig. 7 and 8).
 |
| Fig. 1: | 2-D graphical representation of combined root length (green) and shoot length (blue) at 26 days of all hydroponic genotypes Error bar represents standard errors |
 |
| Fig. 2: | 2-D graphical representation of combined root length (green) and shoot length (blue) at day 37 of all hydroponic genotypes Error bar represents standard errors |
 |
| Fig. 3: | 2-D graphical representation of combined root length (green) and shoot length (blue) at day 52 of all hydroponic genotypes Error bar represents standard errors |
The highest variation of the shoot length showed at day 42 where it explained 91.83% variation by accessions. The proportion variation explained by genotypes for the maximum visible root length showed an increasing pattern from day 24-45 respectively. Among the landraces genotype Azucena, BR 19, BR 3, BR 22 and BRRI 60 had the longest visible roots which were higher at all stages than the mean root length. IR 64, BR 14 and BR 32 had the shortest root systems (Fig. 8).
Comparing the genotypes with maximum visible root length and shoot length at day 46, the harvesting time BR 19, BR 22, BR 60, BR 3 and Azucena showed the highest root length where Azucena, BR 14 and BR 32 showed the highest shoot length. BR 32 and BR 14 showed almost similar shoot length at that time. IR 64 and BR 32 was the lowest on both traits (Fig. 8).
The root angle spread at day 46 was highly statistically different between genotype (R2 = 69.79%). BR 14 and BR 60 had a wide distributed root angle of approximately 139°. BR32 had the lower root angle which was measured at 118° at that time (Fig. 9).
 |
| Fig. 4: | 2-D line graph showing the increase in maximum root length from day 5-52 after sowing (DAS) of check cultivars Error bar represents standard errors |
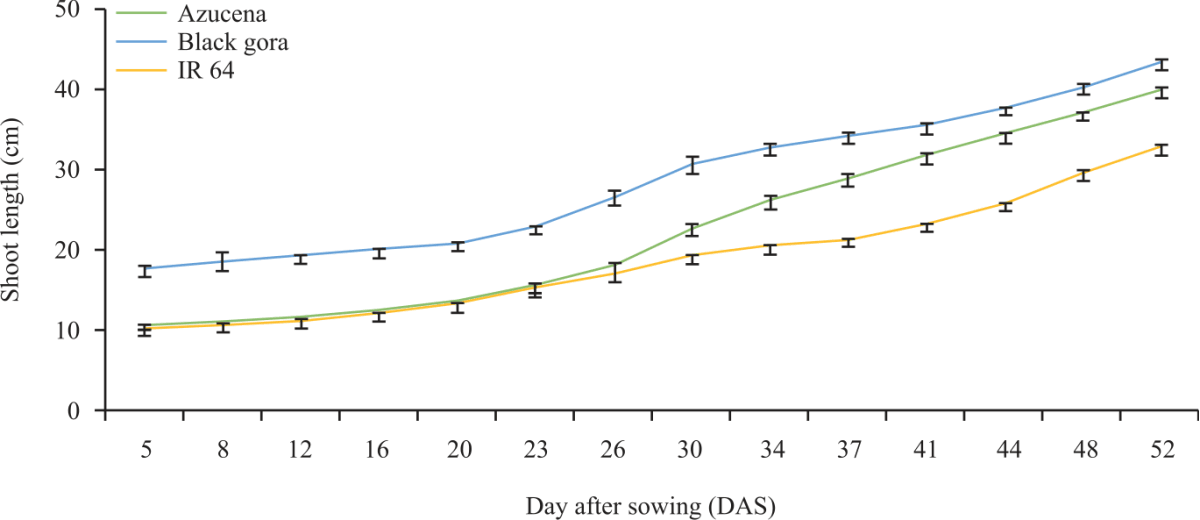 |
| Fig. 5: | 2-D line graph showing the increase in maximum shoot length from day 5-52 after sowing (DAS) of check cultivars Error bar represents standard errors |
 |
| Fig. 6: | Increase in maximum visible root length from 10-42 days after sowing for five cultivars Error bar represents standard errors |
| Table 1: Means of all rhizotron shoot and root data with F, P and R2 (adj) value of one way ANOVA | ||||
| Trait | Mean | F | P | R2 (adj) |
| Maximum visible root length day 10 (cm) | 20.56 | 2.55 | 0.041 | 25.95 |
| Maximum visible root length day 14 (cm) | 27.39 | 3.12 | 0.017 | 32.37 |
| Maximum visible root length day 21 (cm) | 41.39 | 1.11 | 0.386 | 2.52 |
| Maximum visible root length day 28 (cm) | 66.81 | 0.81 | 0.589 | 0 |
| Maximum visible root length day 35 (cm) | 85.04 | 2.35 | 0.05 | 23.36 |
| Maximum visible root length day 42 (cm) | 98.93 | 15.38 | 0.00 | 76.45 |
| Maximum visible root length day 46 (cm) | 105.64 | 13.2 | 0.00 | 73.37 |
| Shoot length day 10 (cm) | 25.34 | 2.55 | 0.041 | 25.95 |
| Shoot length day 14 (cm) | 30.26 | 5.93 | 0.00 | 52.69 |
| Shoot length day 21 (cm) | 39.98 | 9.39 | 0.00 | 65.45 |
| Shoot length day 28 (cm) | 52.18 | 12.64 | 0.00 | 72.45 |
| Shoot length day 35 (cm) | 63.43 | 31.89 | 0.00 | 87.46 |
| Shoot length day 42 (cm) | 73.89 | 50.79 | 0.00 | 91.83 |
| Shoot length day 46 (cm) | 77.51 | 32.34 | 0.00 | 87.62 |
| Root angle at day 46 | 135.78 | 11.23 | 0.00 | 69.79 |
| Shoot dry weight (g) | 5.69 | 182.2 | 0.00 | 97.61 |
| Root mass (g) | 8.50 | 54.27 | 0.00 | 92.32 |
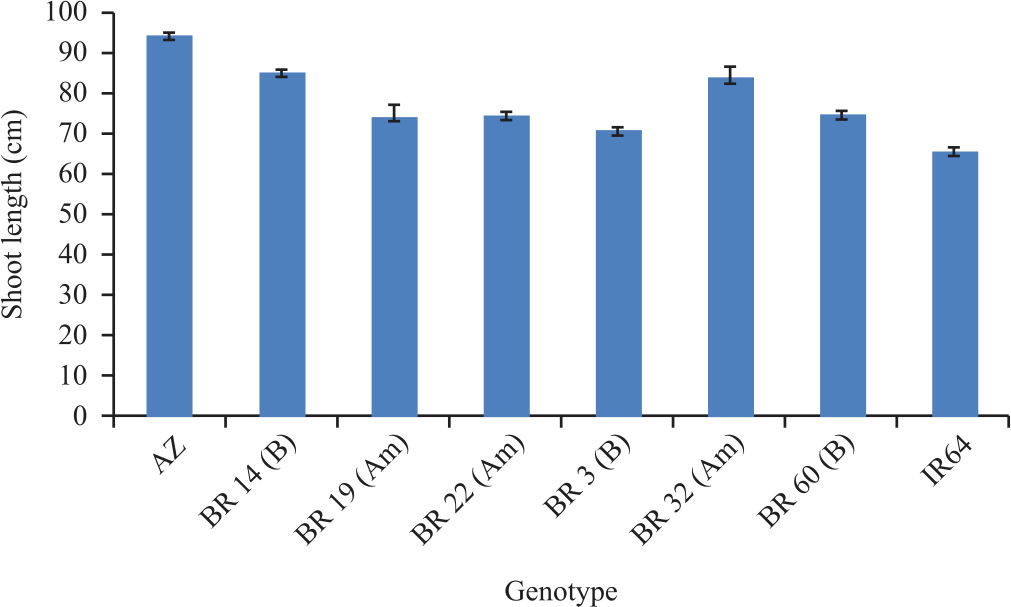 |
| Fig. 7: | Maximum shoot length at 42 DAS for the cultivars Error bar represents standard errors |
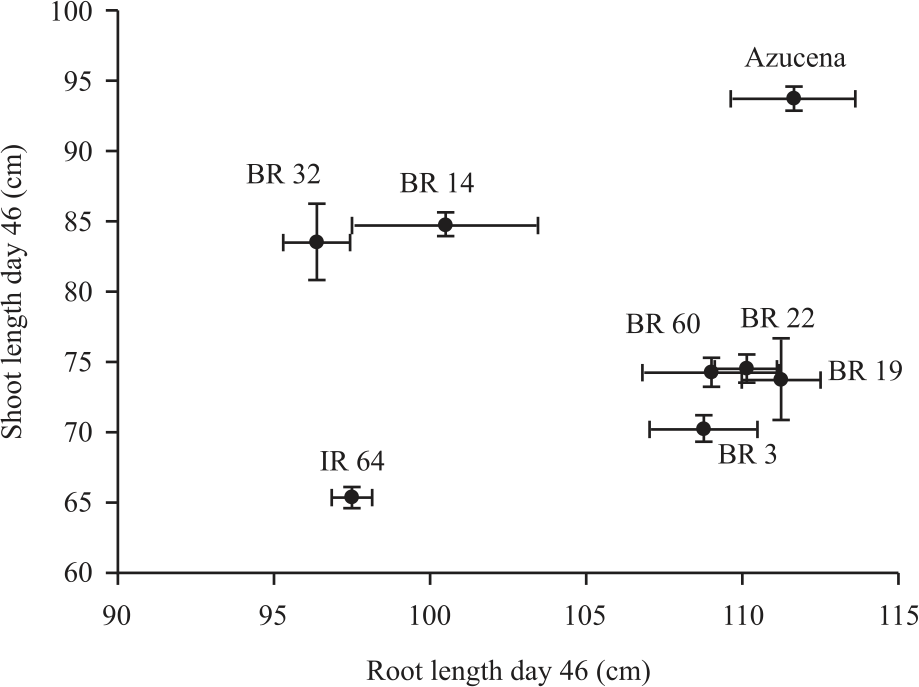 |
| Fig. 8: | Root and shoot length at 46 DAS of all rhizotron genotypes Error bars represent standard errors |
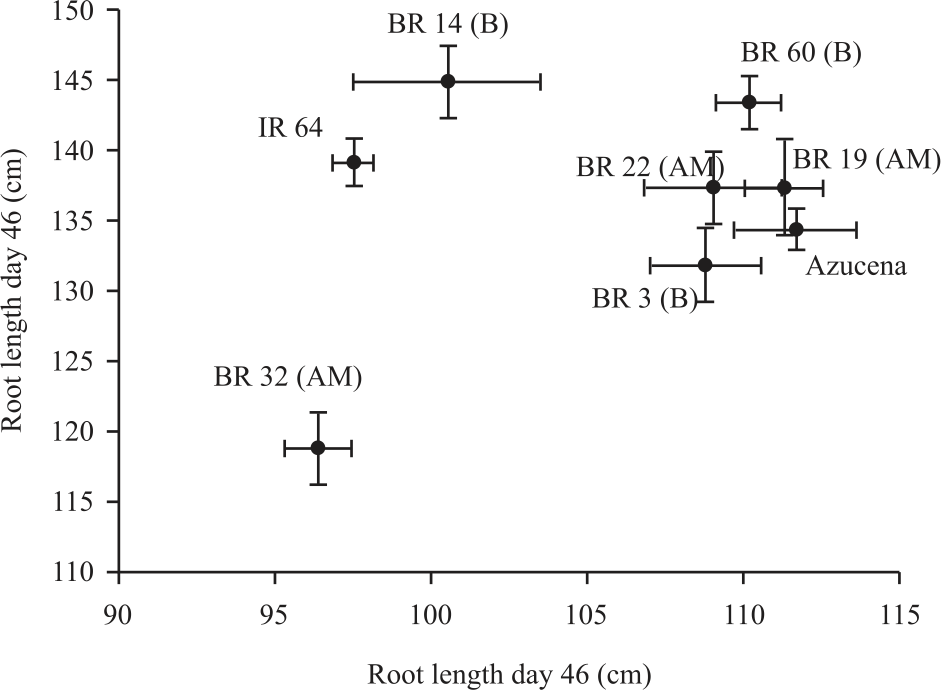 |
| Fig. 9: | Root length day 46 and root angle day 46 of all rhizotron cultivars Error bars represent standard errors |
For the root, dry weight BR 19 and BR 3 had the highest root dry weight which was higher than that of BR 32 (Fig. 10). The root mass of the different genotype represented in Fig.10. Root mass showed great discrimination among the cultivars. Analysis of variance (ANOVA) revealed 92.32% variation explained by the accessions. Among the measured traits plant biomass showed the proportion of variation explained by the cultivars was 97.6%.
The maximum visible root length of the check varieties (Fig. 11) shows that Azucena long with vertical axes (Fig. 9) and IR 64 had shallow rooted which follows the same pattern as previously detected in Al-Shugeairy et al.32. The maximum visible shoot length from 21-45 Days After Sowing (DAS) of the check varieties (Fig. 12) Azucena and IR64 follows the same trends as described in Al-Shugeairy et al.32.
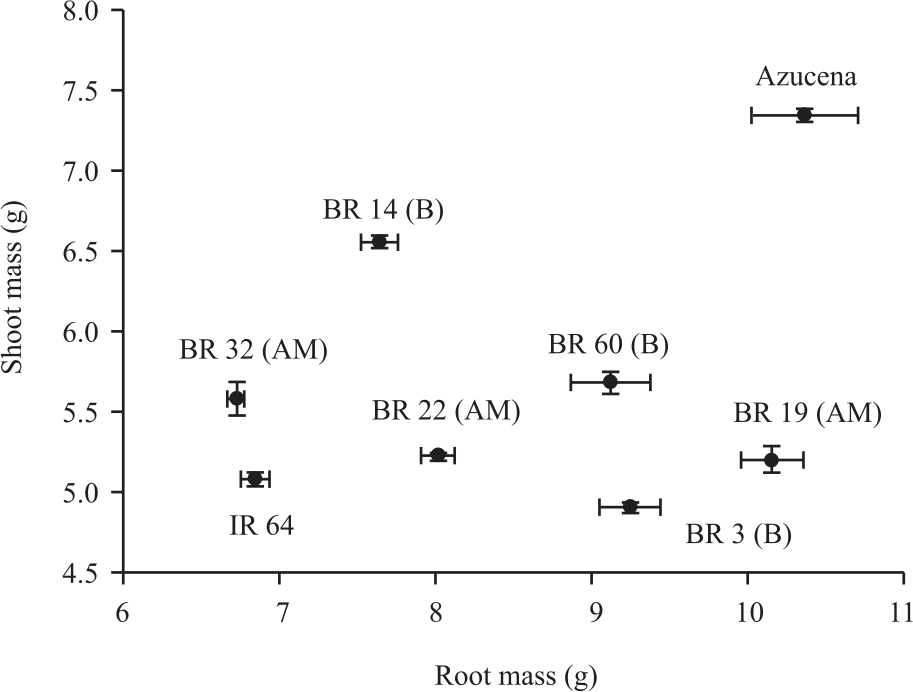 |
| Fig. 10: | Total root mass plotted against total plant dry weight at 46 DAS for eight cultivars along with check varieties Azucena (deep, vertical roots) and IR64 (Shallow, more horizontal roots Bar represents standard error |
Correlation between rhizotron traits: The data collected from rhizotron root and shoot traits were closely related to each other. Many traits showed a strong correlation, the most significant correlations are described below.
Plant biomass had a very strong correlation with many of the root traits. It strongly correlated with maximum visible root length at all days, which was strongest at day10 (r = 0.55, p = 0.001).
Maximum visible root length correlated with other roots and shoot traits. At all days maximum visible root length correlated with shoot length at all days, the strongest correlation observed in maximum visible root length at day 21 and shoot length day 10 (r = 0.502, p = 0.003). Maximum visible root length did not correlate with root angle.
Comparing the maximum root length data in hydroponic screening, significant variation was observed within the accessions. But according to Shrestha et al.30 hydroponic is probably not the most reliable method for testing rooting depth30. The author suggests that rhizotrons are better though this rhizotron method requires much more effort. The genotypic variation observed in the hydroponic medium reflects the variation that also occurs in the soil. Shrestha et al.30 reported that a very good agreement between root growth in a liquid medium and root growth in the soil pot. Evidence suggests that the correlation between herbicide score and root length in the hydroponic medium reflects that agreement33.
Considering the rhizotron screening data, high heritability and significant trait variation suggest a clear and distinguishable difference within the accessions. The rhizotron data suggested that BR 19, BR 22 and BR 60 has the highest maximum visible root length among the cultivars tested. They had the highest root mass which indicated that the biggest rooting system (Fig. 10). Comparing the root and shoot mass, BR 14 had the highest shoot mass with low root mass revealing this as a cultivar with a remarkably low relative root mass. BR 19 and BR 3 had the highest root dry weight with low shoot mass. The highest root mass indicates well-developed root systems in addition to the shoot mass. The evidence suggested that in different planting practice Boro rice cultivars showed the best rooting behaviour like the number of roots, root length density than the other varieties34. Another study also suggested that Boro rice cultivars showed better yield in strongly saline soils to all parameters studies35. It is noted that cultivars identified deep-rooted here in this current study BR 3, BR 19, BR 60 is also from the Boro rice group.
Comparing the root length of the accessions tested in the rhizotron, they can be ranked as BR 19>BR 22>BR 60>BR 3. In the hydroponic screening system, comparing the root length data, ranked can be BR 60>BR 3>BR 19>BR 22 among the 16 accessions tested. Though in hydroponic screening systems and rhizotron screening system the ranked is a bit different, the rooting behaviour of these four cultivars is similar in these two methods.
The check varieties had similar rooting behaviour in the previously reported by Al-Shugeairy et al.30 and Shrestha et al.31. The report suggested that Azucena had a long root with vertical axes whereas IR 64 had a shallow-rooted lateral branching system. The traits information like root and shoot length, root and shoot biomass and root angle collected from the current study would be very valuable for future study. The accessions identified as deep and shallow-rooted, field trial under drought and yield determination for these cultivars would be very important. At the same time, drought screening of these cultivars would be valuable to identify drought resistance cultivars to elucidate the natural genetic diversity within rice to breed better rice cultivars. The two methods hydroponic and rhizotron system, used in this study has the advantage that root and shoot growth can be continually assessed. The rank of the tested cultivars, based on these two experiments is a bit different due to the two systems, one is liquid media (hydroponic) and another is soil-based systems. In a hydroponic system though the root growth is monitored, however, there was an issue of maintaining the pH of the media on regular basis and the root of the accessions intertwine with each other. Like hydroponic, in the rhizotron method root growth and root angle can be continually measured, but due to 2D construction, the root growth is restricted in an all-natural direction as well as a very labour-intensive screening method.
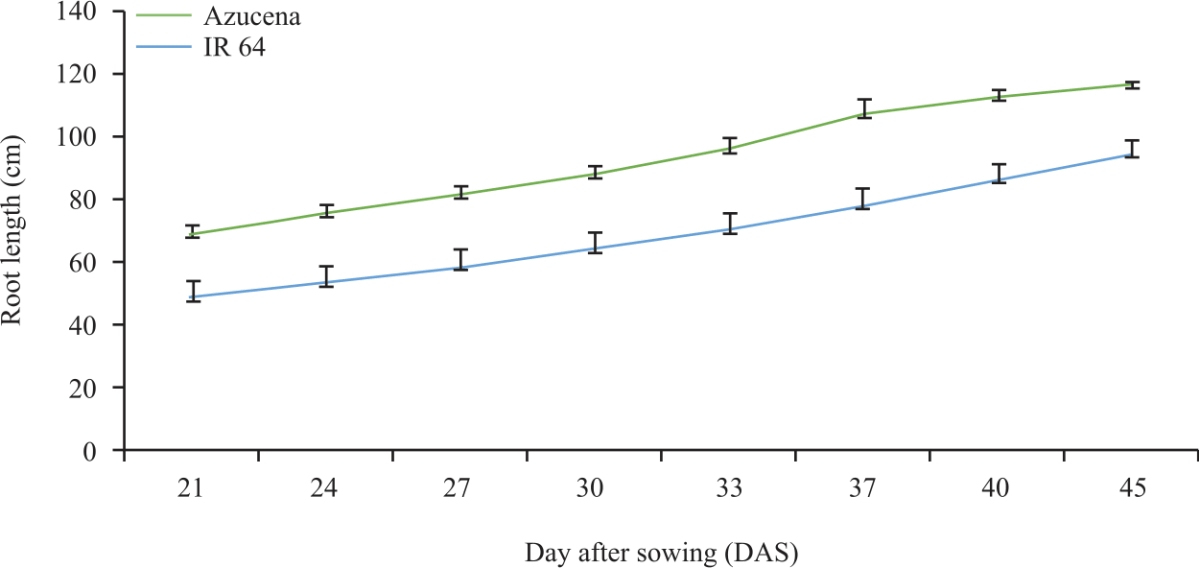 |
| Fig. 11: | Increase in maximum visible root length from 21-45 days after sowing for two check varieties (Azucena and IR64) Error bar represents standard errors |
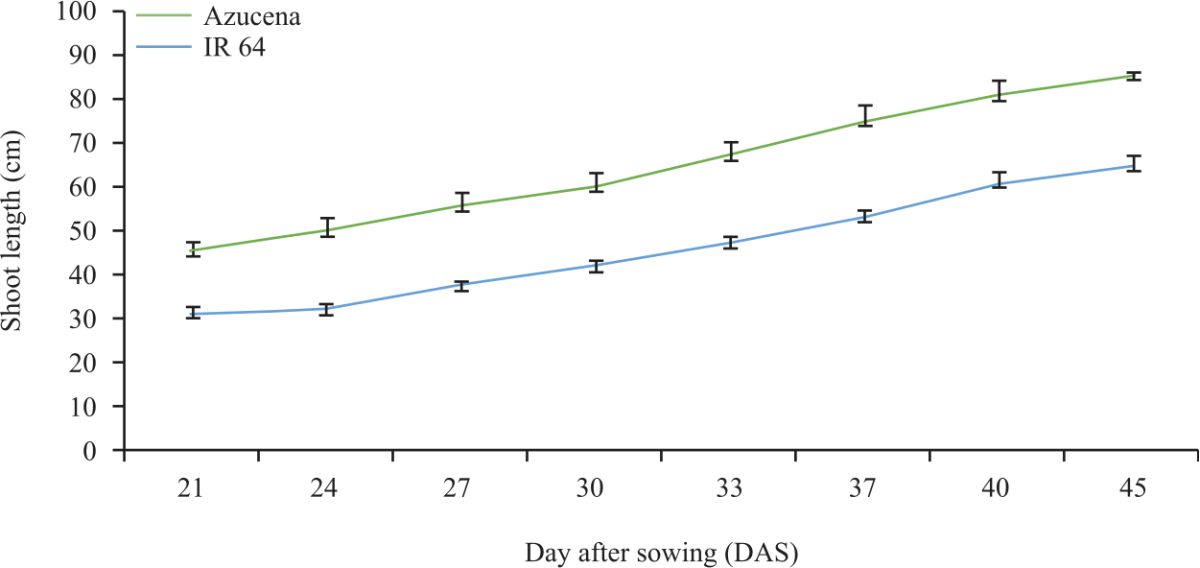 |
| Fig. 12: | Increase in maximum visible shoot length from 21-45 days after sowing for two check varieties (Azucena and IR64) Error bar represents standard errors |
CONCLUSION
The hydroponic and rhizotron data suggested that BR 19 (Boro), BR 22 (Amon), BR 60 (Boro) and BR 3 (Boro) has the deepest rooting systems than the other accessions tested in these screening methods. The rhizotron data also suggested that BR19 and BR 3, both are Boro cultivars had surprisingly deep and shallow rooting systems with very low shoot mass among the cultivars. From the study, for example, BR 19, BR 22 and BR 3 is a very promising cultivar that needs to be assessed under field conditions. If these traits prove to be valuable under field conditions these cultivars have the scope for use in rice breeding programmes.
SIGNIFICANCE STATEMENT
Comparing the assessment of the screening methods, four accessions were considered as deeply rooted genotypes which may contribute in a greater way to the development of drought resistance variety in rice. The trait data which was collected from the study could be very helpful for the crop researcher to interpret the other drought data. It is, therefore, necessary to screen these cultivars at the field level under drought condition. The development of drought-tolerant cultivars and potential sources of tolerance will bring a direct impact on rice-eating countries and help to cope up with food insecurity.
REFERENCES
- Caicedo, A.L., S.H. Williamson, R.D. Hernandez, A. Boyko and A. Fledel-Alon et al., 2007. Genome-wide patterns of nucleotide polymorphism in domesticated rice. PLoS Genet., 3: 1745-1756.
CrossRefDirect Link - Dawe, D, 2000. The contribution of rice research to poverty alleviation. Stud. Plant Sci., 7: 3-12.
CrossRefDirect Link - Ahmadi, N., A. Audebert, M.J. Bennett, A. Bishopp and A.C. de Oliveira et al., 2014. The roots of future rice harvests. Rice, Vol. 7.
CrossRef - Serraj, R., L. Krishnamurthy, J. Kashiwagi, J. Kumar, S. Chandra and J.H. Crouch, 2004. Variation in root traits of chickpea (Cicer arietinum L.) grown under terminal drought. Field Crops Res., 88: 115-127.
CrossRefDirect Link - Lobet, G., A. Paez-Garcia, H. Schneider, A. Junker, J.A. Atkinson and S. Tracy, 2019. Demystifying roots: A need for clarification and extended concepts in root phenotyping. Plant Sci., 282: 11-13.
CrossRefDirect Link - Ye, H., M. Roorkiwal, B. Valliyodan, L. Zhou, P. Chen, R.K. Varshney and H.T. Nguyen, 2018. Genetic diversity of root system architecture in response to drought stress in grain legumes. J. Exp. Bot., 69: 3267-3277.
CrossRefDirect Link - Yu, J., G. Pressoir, W.H. Briggs, I.V. Bi and M. Yamasaki et al., 2006. A unified mixed-model method for association mapping that accounts for multiple levels of relatedness. Nat. Genet., 38: 203-208.
CrossRefDirect Link - Pandey, S. and H. Bhandari, 2009. Drought: Economic Costs and Research Implications. In: Drought Frontiers in Rice: Crop Improvement for Increased Rainfed Production, Serraj, R., J. Bennett and B. Hardy (Eds.)., World Scientific Publishing, New Jersey, pp: 3-17.
CrossRefDirect Link - Mottaleb, K.A., R.M. Rejesus, M.V.R. Murty, S. Mohanty and T. Li, 2017. Benefits of the development and dissemination of climate-smart rice: Ex ante impact assessment of drought-tolerant rice in South Asia. Mitigation Adapt. Strategies Global Change, 22: 879-901.
CrossRefDirect Link - Kato, Y., A. Kamoshita, J. Yamagishi, H. Imoto and J. Abe, 2007. Growth of rice (Oryza sativa L.) cultivars under upland conditions with different levels of water supply 3. Root system development, soil moisture changeand plant water status. Plant Prod. Sci., 10: 3-13.
CrossRefDirect Link - Eapen, D., J. Martínez-Guadarrama, O. Hernández-Bruno, L. Flores, J. Nieto-Sotelo and G.I. Cassab, 2017. Synergy between root hydrotropic response and root biomass in maize (Zea mays L.) enhances drought avoidance. Plant Sci., 265: 87-99.
CrossRefDirect Link - Hund, A., N. Ruta and M. Liedgens, 2009. Rooting depth and water use efficiency of tropical maize inbred lines, differing in drought tolerance. Plant Soil, 318: 311-325.
CrossRefDirect Link - Uga, Y., K. Sugimoto, S. Ogawa, J. Rane and M. Ishitani et al., 2013. Control of root system architecture by deeper rooting 1 increases rice yield under drought conditions. Nat. Genet., 45: 1097-1102.
CrossRefDirect Link - Suwanmontri, P., A. Kamoshita and S. Fukai, 2021. Recent changes in rice production in rainfed lowland and irrigated ecosystems in Thailand. Plant Prod. Sci., 24: 15-28.
CrossRefDirect Link - Uga, Y., K. Okuno and M. Yano, 2011. Dro1, a major QTL involved in deep rooting of rice under upland field conditions. J. Exp. Bot., 62: 2485-2494.
Direct Link - Zhao, K., C.W. Tung, G.C. Eizenga, M.H. Wright and M.L. Ali et al., 2011. Genome-wide association mapping reveals a rich genetic architecture of complex traits in Oryza sativa. Nat. Commun., Vol. 2.
CrossRefDirect Link - Gowda, V.R.P., A. Henry, A. Yamauchi, H.E. Shashidhar and R. Serraj, 2011. Root biology and genetic improvement for drought avoidance in rice. Field Crops Res., 122: 1-13.
CrossRefDirect Link - Bouman, B.A.M., E. Humphreys, T.P. Tuong and R. Barker, 2007. Rice and water. Adv. Agron., 92: 187-237.
CrossRefDirect Link - Edwards, J., C. Johnson, C. Santos-Medellín, E. Lurie and N.K. Podishetty et al., 2015. Structure, variation, and assembly of the root-associated microbiomes of rice. Proc. Nat. Acad. Sci., 112: E911-E920.
CrossRefDirect Link - Ranathunge, K., L. Kotula, E. Steudle and R. Lafitte, 2004. Water permeability and reflection coefficient of the outer part of young rice roots are differently affected by closure of water channels (aquaporins) or blockage of apoplastic pores. J. Exp. Bot., 55: 433-447.
CrossRefDirect Link - Wang, H.Z., K.Z. Yang, J.J. Zou, L.L. Zhu and Z.D. Xie et al., 2015. Transcriptional regulation of PIN genes by four lips and MYB88 during Arabidopsis root gravitropism. Nat. Commun., Vol. 6.
CrossRefDirect Link - Ramalingam, P., A. Kamoshita, V. Deshmukh, S. Yaginuma and Y. Uga, 2017. Association between root growth angle and root length density of a near-isogenic line of IR64 rice with deeper rooting 1 under different levels of soil compaction. Plant Prod. Sci., 20: 162-175.
CrossRefDirect Link - Kanbar, A., M. Toorchi and H. Shashidhar, 2009. Relationship between root and yield morphological characters in rainfed low land rice (Oryza sativa L.). Cereal Res. Commun., 37: 261-268.
CrossRefDirect Link - Han, J.H., N.H. Shin, J.H. Moon, J.H. Chin and S.C. Yoo, 2016. A simple phenotyping method for deep-rooting rice grown in pots. J. Plant Biotechnol., 43: 444-449.
CrossRefDirect Link - MacMillan, K., K. Emrich, H.P. Piepho, C.E. Mullins and A.H. Price, 2006. Assessing the importance of genotype × environment interaction for root traits in rice using a mapping population II: Conventional QTL analysis. Theor. Appl. Genet., 113: 953-964.
CrossRefDirect Link - Chen, C., G.J. Norton and A.H. Price, 2020. Genome-wide association mapping for salt tolerance of rice seedlings grown in hydroponic and soil systems using the bengal and assam aus panel. Front. Plant Sci., Vol. 11.
CrossRefDirect Link - Price, A.H., K.A. Steele, B.J. Moore and R.G.W. Jones, 2002. Upland rice grown in soil-filled chambers and exposed to contrasting water-deficit regimes: II. Mapping quantitative trait loci for root morphology and distribution. Field Crops Res., 76: 25-43.
CrossRefDirect Link - Norton, G.J. and A.H. Price, 2009. Mapping of quantitative trait loci for seminal root morphology and gravitropic response in rice. Euphytica, 166: 229-237.
CrossRefDirect Link - Shrestha, R., Z. Al‐Shugeairy, F. Al‐Ogaidi, M. Munasinghe, M. Radermacher, J. Vandenhirtz and A.H. Price, 2014. Comparing simple root phenotyping methods on a core set of rice genotypes. Plant Biol., 16: 632-642.
CrossRefDirect Link - Elec, V., C.A. Quimio, R. Mendoza, A.G.C. Sajise, S.E.J. Beebout, G.B. Gregorio and R.K. Singh, 2013. Maintaining elevated Fe2+ concentration in solution culture for the development of a rapid and repeatable screening technique for iron toxicity tolerance in rice (Oryza sativa L.). Plant Soil, 372: 253-264.
CrossRefDirect Link - Al-Shugeairy, Z., M.S. Islam, R. Shrestha, F. Al-Ogaidi, G.J. Norton and A.H. Price, 2014. High throughput screening of rooting depth in rice using buried herbicide. Ann. Appl. Biol., 165: 96-107.
CrossRefDirect Link - Munasinghe, M. and A.H. Price, 2016. Genetic and root phenotype diversity in Sri Lankan rice landraces may be related to drought resistance. Rice, Vol. 9.
CrossRef - Uddin, M.R., L.J. Wade, J.Y. Pyon and M.A. Mazid, 2009. Rooting behavior of rice cultivars under different planting methods. J. Crop Sci. Biotechnol., 12: 17-23.
CrossRefDirect Link - Islam, S.S. and M.Z. Hossain, 2020. Morphological and yield performance of rice varieties grown in moderately and strongly saline soils of Khulna, Bangladesh. Global Sci. J., 8: 582-596.
Direct Link