
Olubukola Grace Abraham
Department of Botany, Obafemi Awolowo University, Ile-Ife, Osun State, Nigeria
LiveDNA: 234.18205
Chinyere Constance Nwokeocha
Natural History Museum, Obafemi Awolowo University, Ile-Ife, Osun State, Nigeria
Richard Olutayo Akinwale
Department of Crop Production and Protection, Obafemi Awolowo University, Ile-Ife, Osun State, Nigeria
Julius Olaoye Faluyi
Department of Botany, Obafemi Awolowo University, Ile-Ife, Osun State, Nigeria
Journal of Plant Sciences
Year: 2017 | Volume: 12 | Issue: 2 | Page No.: 52-58







ABSTRACT
Background and Objective: The continual research into increasing yield in rice is very important in ensuring food security. There has been stagnation in yield/hectare of rice since the introduction of IR8 in the 1980s, therefore there is the need for breeders to go back to the wild relatives and landraces of Oryza sativa, which have proven to be veritable sources of genes for germplasm improvement. This study was aimed at highlighting vegetative characters that are directly associated to yield which can help the breeder during selection process. Materials and Methods: Thirty-seven accessions selected from four local varieties of landraces were used in this study. These were planted on the field in rows of fourteen plants, each row representing an accession. Data were collected in the accessions on vegetative traits and yield characters. The data were subjected to one-way analysis of variance (ANOVA) to assess diversity among accessions. Pearson correlation coefficients was used to determine relationships among characters and principal components analysis was used to identify major attributes that are responsible for the variations observed in the accessions. Results: Results showed that the number of days to booting and maturity highly correlated with the total number of spikelets produced as well as the leaf area index (LAI) of both the penultimate and flag leaves. The two groups of yield patterns observed were intermediate or late maturing, high tillering, dense panicle with low TGW (1000-grain weight) on one side and early maturing, low tillering, less dense or sparse panicle with high TGW on the other. Conclusion: The length of the ligule was shown to be the significant distinguishing morphological character between these two categories.
PDF Abstract XML References Citation
Received: September 05, 2017;
Accepted: September 22, 2017;
Published: September 28, 2017
How to cite this article
Olubukola Grace Abraham, Chinyere Constance Nwokeocha, Richard Olutayo Akinwale and Julius Olaoye Faluyi, 2017. Screening for Yield Related Characters in Some Landrace Accessions of Oryza sativa Linn. in Nigeria. Journal of Plant Sciences, 12: 52-58.
DOI: 10.3923/jps.2017.52.58
URL: https://scialert.net/abstract/?doi=jps.2017.52.58
DOI: 10.3923/jps.2017.52.58
URL: https://scialert.net/abstract/?doi=jps.2017.52.58
INTRODUCTION
The Oryza genus belong to the sub-family Oryzae and family Poaceae, possessing about 25 species occurring in the tropical and subtropical regions of Africa, Asia, Australia and South America1. While it is widely accepted that Asia is the center of origin for the Oryza species, some researchers have considered it to be Africa since it is the continent with, by far, the largest number of indigenous species2. The most common of the members of this family is Oryza sativa Linn. (also called Asian rice), a common staple cereal crop which had fed millions of people over the years. The recent population boom has, however, increased the need for maximum production of rice without necessarily increasing the agricultural inputs or cultivation area3-5. The econometric International Model for Policy Analysis of Agricultural Commodities and Trade (IMPACT ) was used to predict that the production of rice has to increase by 43% before 20306.
The genetic richness of the rice germplasm is being threatened by the extensive promotion of improved varieties by seed companies and its indiscriminate use by farmers, leading to constant erosion of the indigenous varieties or landraces7. Zeven8 defined landraces as ‘varieties which have the capacity to tolerate biotic and abiotic stress, giving them high yielding stability (i.e., can give high yield under varying environmental conditions) and a moderate yield level when cultivated with low input agricultural system. In maize, landraces and wild varieties have been widely used in breeding for higher yields, drought tolerance, disease resistance among others over the years. However, in rice, breeding for higher yield have stagnated at 10 t h–1 since the introduction of IR8 in the 1980s, which is a semi-dwarf tropical indica inbred variety1. The semi-dwarf stature and photoperiod insensitivity which were characters that were peculiar to this hybrid (IR8) have unfortunately yielded little success for improvements, since maximum yield is mostly determined by the genetic richness of a variety and the environment9,10.
Monteith11 carried out a study on some cereal crops and determined that the amount of insolation directly affect yield. However, Long et al.9, was able to prove that the genetic makeup of a plant is of more importance that the plant’s available photosynthetic surfaces since sink size i.e., yield is a product of the efficiency by which a plant can utilize and store photosynthates. This is essentially due to the fact that increase in leaf area will lead to increase in self-shading i.e., the upper canopy of leaves cast shade on the lower canopy thereby limiting the amount of insolation available to them. Therefore, Long et al.9, designed an ideal rice plant which will be efficient in energy usage as one possessing thin leaves (which are better adapted to photosynthetic efficiency) with lower canopy of lax leaves and top canopy of erect leaves, so that the upper leaves can intercept minimal amount of energy and the remaining energy can be used by the lower canopy.
Furthermore, Montieth11 inferred that increase in maturity period of cereal plants can increase yield since the plants will have more time to build vegetative biomass which can, in turn, help in the final grain filling hence increase in yield. This was corroborated by Efisue et al.12, who determined that plants with moderate maturity period and high leaf area index produce more spikelets/plant.
Yoshida13 had however pointed out that highlighting yield components would yield a meaningful blueprint or template for selecting and breeding for the target yield, since yield is a complex character which is a function of interaction of the plant genes and the environment9,10.
The objective of this study therefore was to identify reliable secondary traits that can be used for indirect selection for improved grain yield of landraces of rice. This study will help both the rice researchers and breeders to identify promising high-yielding rice plants from landraces for improving known varieties or ultimate development of new lines.
MATERIALS AND METHODS
A field experiment was conducted in the Department of Botany, Obafemi Awolowo University between 2013 and 2016. Thirty-seven accessions were selected from four landrace (collected from different agro-ecologies, Table 1) population and proper characterization was made on them through 2 planting seasons between 2014 and 2015. The selected accessions were seeded in nurseries and transplanted to the field when they were two weeks old, using a plot size of 18.7×10.5 m2. Each accession was planted in 14 replicates and data was collected on the best five at maturity. The vegetative characters considered were: Days to booting (DB), days to maturity (DM), culm length (CL), number of tillers (NT), average ligule length (AvLi), flag leaf area index (FLAI), penultimate leaf area index (PLAI). The following reproductive parameters were also documented: Panicle length (PL), number of filled spikelets/panicle (NFS), number of unfilled spikelets/panicle (NUS), Percentage of filled spikelets/panicle (PFS), Panicle density/panicle (PD) and 1000-grain weight/plant (TGW). Panicle density was calculated using the formula below:
| Table 1: | Agro-ecological zone and location of landraces used in this study |
 | |
![]()
Statistical analysis: Data were subjected to analysis using Statistical Analysis Software (SAS), version 9.0 (SAS Institute, Cary, North Carolina, USA). One-way Analysis of Variance (ANOVA) was used to determine the significance of the variance within accessions. The TGW was measured/plant, not in 5 replicates, hence not included in the ANOVA procedure). Pearson correlation coefficients was used to determine the strength and significance of the relationships among the vegetative and yield characters. Principal components analysis was used to determine the major morphological characters that contribute to the variations observed in the population under study. The correlation coefficients were used to construct a cluster diagram using Paleontological Statistics Software (PAST) (Version 2.17).
Probability level used was 0.01<p<0.05.
RESULTS
The DB and DM showed that the accessions ranged from early to late maturing, CL indicated the accessions ranged from dwarf to tall individuals and NT showed they ranged from low to high tillering plants (Table 2). Also, the PF ranged from very low (about 43%) to high (100%) and panicle density ranged from sparse to dense. Results from ANOVA showed that all characters considered varied significantly among accessions (Table 2).
Table 3, PL was shown to have highly significant correlation with CL, PLAI and FLAI. NFS had highly significant correlation with DB, DM, NT, PLAI, FLAI and AvLi. NUS had highly significant correlation with DB, DM, NT and AvLi. TNS had highly significant correlation with DB, DM, NT, PLAI, FLAI and AvLi. PFS had negative but highly significant correlation with DB and DM. PD showed highly significant correlation with DB, DM, NT and AvLi. TGW had negative but highly significant correlation with DM, DM, CL AvLi and PFS.
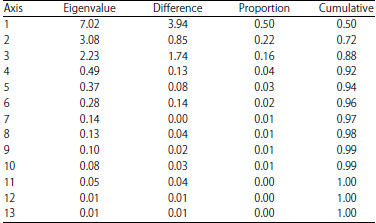
The principal components analysis (Table 4) showed the factor loading of characters on 13 axes with the first three axes being the most important. The first principal component, PC1 (Table 5) is of primary importance having eigen proportion of 0.50. The traits associated with this axis were DB (0.35), DM (0.34), NT (0.31), AvLi (0.34), TNS (0.34), PD (0.32) and TGW (-0.34). Also, attributes like PLAI (0.50), FLAI (0.49), PL (0.47) and CL (0.43) are of secondary importance since they loaded on the second principal components axis with an eigen proportion of 0.22. The percentage of filled spikelets (0.58) and NFS (0.43) loaded on the PC3 indicating (eigen proportion = 0.16) that they are of tertiary importance.
The cluster analysis as shown in Fig. 1 and Appendix 1 divided the accessions under study into two broad groups:
| • | Early maturing, low tillering, low panicle density and high 1000-grain weight which comprised of the Ijesa-Isu and Ikole accessions, and |
| • | Intermediate/Late maturing, high tillering, high panicle density and low 1000-grain weight, comprising the Awgu and Ifewara accessions |
DISCUSSION
Results in this study showed that DB and DM had positive and highly significant correlation with TNS and PD. The magnitude of this relationship as revealed by r2 which is ≈0.45 indicated that the length of maturity period is of importance to number of grains produced/plant. This means that the number of spikelets produced/panicle is a direct effect of the length of the period involved in building vegetative biomass and panicle production, supporting the findings of Montieth11 and Long et al.9. Peng et al.14 and Kropff et al.15, have also observed in separate studies that there is a positive relationship between growth duration and the period of time from panicle initiation to full heading. As such, early maturing plants have shorter period of time for panicle production compared to the late maturing ones, hence, less number of spikelets/panicle. This in effect is an indication that yield in terms of number of grains produced is a function of maturity period in rice i.e. the higher the maturity period, the higher the number of grains produced. The above elucidation is also true for NT, indicating that individuals that produces more tillers are more likely to have high grain yield/panicle.
| Table 2: | Mean, range and f-values of the characters accessed in the accessions study |
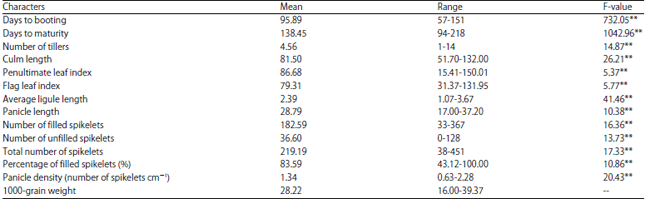 | |
Panicle density: Number of spikelets per unit cm of the panicle, 1000-grain weight was not measured in replicates and thus was not included in the ANOVA procedure ** = Highly significant i.e., p<0.01, * = Significant i.e., 0.01< p<0.05 | |
| Table 3: | Pearson correlation coefficients of the variables accessed in the study |
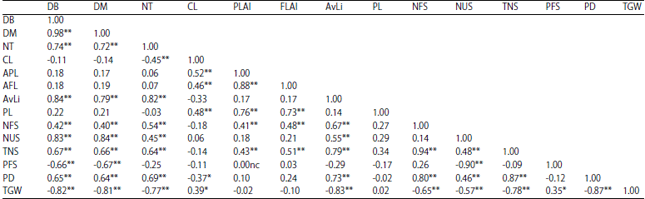 | |
DB = Days to booting, DM = Days to maturity, NT = Number of tillers, CL = Culm length, PLAI = Penultimate leaf area index, FLAI = Flag leaf area index, AvLi = Average ligule length, PL=Panicle length, NFS = Number of filled spikelets, NUS = Number of unfilled spikelets, TNS = Total number of spikelets, PFS = Percentage of filled spikelets, PD = Panicle density and TGW = 1000-grain weight, ** = Highly significant i.e., p<0.01, * = Significant i.e., 0.01< p<0.05; nc = No correlation | |
| Table 4: | Eigen values, eigen differences and proportions of variation contribution of thirteen principal components axes |
 | |
This is in support of the findings of Rao et al.16, that the gene that signals the development of tiller meristems is also responsible for determining panicle architecture i.e., panicle branching and number of spikelets produced/panicle.
The high positive correlation that exists between days to maturity and NUS and its (days to maturity) high negative correlation with PFS point to the fact that grain filling may be low for late maturing plants than the early maturing ones. Long et al.9, pointed out that this is a function of individual plant and its genetic makeup.
| Table 5: | Factor loading of thirty-seven accessions of landraces of rice for fourteen characters on three principal components axes |
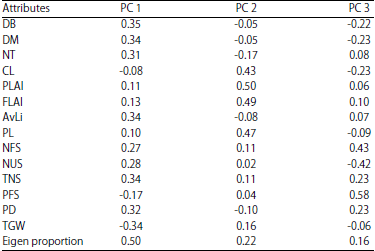 | |
DB = Days to booting, DM = Days to maturity, NT = Number of tillers, CL = Culm length, PLAI = Penultimate leaf area index, FLAI = Flag leaf area index, AvLi = Average ligule length, PL = Panicle length, NFS = Number of filled spikelets, NUS = Number of unfilled spikelets, TNS = Total number of spikelets, PFS = Percentage of filled spikelets, PD = Panicle density and TGW = 1000-grain weight | |
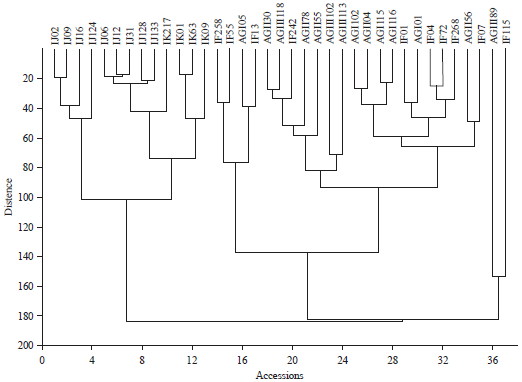 | |
| Fig. 1: | Clustering of the accessions based on the similarities of their quantitative vegetative and reproductive characters |
The PL had highly significant correlation with CL, PLAI and FLAI. This is an indication that these vegetative characters determine how long a panicle will be. However, since the only yield parameter that had highly significant correlation with CL is the PL, we agree with the findings of Ameen et al.17, who posited that there is a weak relationship between paddy yield and culm height. The insignificant correlation of PL with other yield characters i.e., NFS, NUS, TNS, PFS and TGW also suggest that the length of the panicle is neither a function of how much spikelets it will bear nor the weight of the grains. Yang and Zhang18 are also of the opinion that a number of high yielding cultivars which have been released after various experiments fall short in grain yield due to poor grain filling capacity.
The AvLi had highly significant correlation with TNS and PD. However, AvLi has negative but highly significant correlation with TGW (r2 = 0.69). In other words, plants with longer ligules are likely to produce more spikelets or grains but low 1000-grain weight. The above trend of observation for AvLi was also observed in the NT although the correlation coefficients were lower compared with that of the AvLi. It is also worthy to note at this point that though 1000-grain weight may be low in the individuals with high NT, grain weight/plant may prove to be a more reliable method to determine the trade-offs between weight and number of grains in the rice plant as observed by Afiukwa et al.19.
Results from the principal components analysis further strengthens the hypothesis that the ligule length is an important morphological character when considering yield in rice. This stems from the fact that it is the only morphological character that is not directly associated with the reproductive phase of the rice plant life cycle. Hence, it can be used as a selection criterion where the luxury of time is unavailable for reproductive stage selection. This strengthens the position of Sharma et al.20, that selecting for ideotypes of rice plants with desirable character composition helps in achieving better breeding procedures through early elimination of undesirable defects in cultivars.
It is noteworthy to mention here that, since the ligule is an outgrowth of the leaf sheath, forming a kind of support system for internodes at the node, one can say that this is a way by which a plant helps the culm to bear the weight of the panicle it eventually produces. Interestingly, this character can be determined in a rice plant at seedling stage and can be used as a selection criterion for grain yield as indicated by pearson’s correlation coefficients as well as principal components analysis in this study. The efficiency of seedling stage characterization has been demonstrated in rice for drought tolerance21,22, salinity23 among others. While the number of tillers may also be considered as an indicator, the fact that this character is usually not determined until the onset of booting in the rice plant is be its disadvantage against the ligule length as selection criteria.
| APPENDIX 1: | Mean values of the quantitative vegetative and reproductive characters |
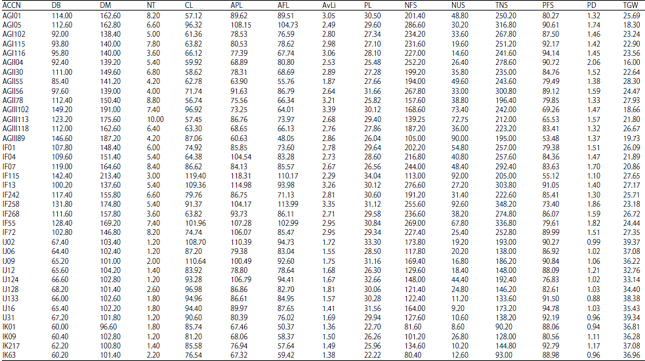 | |
Keys: DB = Days to booting, DM = Days to maturity, NT = Number of tillers, CL = Culm length, APL = Area penultimate leaf, AFL = Area flag leaf, AvLi = Average ligule length, PL = Panicle length, NFS = Number of filled spikelets, NUS = Number of unfilled spikelets, TNS = Total number of spikelets, PFS = Percentage of filled spikelets, PD = Panicle density and TGW = 1000-grain weight | |
CONCLUSION
This study concluded that the period of time from seeding to maturity and the tillering ability of a rice plant are important factors that determine grain yield in terms of number and weight. Also, the ligule length (as observed in this study) is a good indicator to distinguish between the two established yield patterns in the selected landraces. This will be very useful for breeders to select plants from landraces for the improvement of yield in this all important crop.
SIGNIFICANCE STATEMENTS
It was shown in this study that the length of the ligule is of high and important significance in yield determination either for weight or number of grains. This study has therefore provided a solution to the rice breeder’s problem of identifying a high yielding variety in a population of yet-to-be-characterized landrace by using the length of the ligule.
REFERENCES
- Li, G., J. Zhang, C. Yang, Y. Song and C. Zheng et al., 2014. Optimal yield-related attributes of irrigated rice for high yield potential based on path analysis and stability analysis. Crop J., 2: 235-243.
CrossRefDirect Link - Das, P., B. Mukherjee, C.K. Santra, S. Gupta and T. Dasgupta, 2014. Agro-botanical characterization of some released F1 hybrids in rice (Oryza sativa L.). J. Sci. Res. Publ., 4: 1-8.
Direct Link - Cassman, K.G., 1999. Ecological intensification of cereal production systems: Yield potential, soil quality and precision agriculture. Proc. Nat. Acad. Sci., 96: 5952-5959.
CrossRefDirect Link - Zeven, A.C., 1998. Landraces: A review of definitions and classifications. Euphytica, 104: 127-139.
CrossRefDirect Link - Long, S.P., X.G. Zhu, S.L. Naidu and D.R. Ort, 2006. Can improvement in photosynthesis increase crop yields? Plant Cell Environ., 29: 315-330.
CrossRefDirect Link - Sarwar, G., M. Harun-Ur-Rashid, S. Parveen and M. Hossain, 2015. Correlation and path coefficient analysis for agro-morphological important traits in aman rice genotypes (Oryza sativa L.). Adv. Biores., 6: 40-47.
Direct Link - Monteith, J.L., 1977. Climate and the efficiency of crop production in Britain. Proc. R. Soc., 281: 277-294.
Direct Link - Efisue, A.A., B.C. Umunna and J.A. Orluchukwu, 2014. Effects of yield components on yield potential of some lowland rice (Oyza sativa L.) in coastal region of Southern Nigeria. J. Plant Breeding Crop Sci., 6: 119-127.
CrossRefDirect Link - Rao, N.N., K. Prasad and U. Vijayraghavan, 2008. The making of a bushy grass with a branched flowering stem: Key rice plant architecture traits regulated by RFL the rice LFY homolog. Plant Signal Behav., 3: 981-983.
PubMedDirect Link - Ameen, A., Z. Aslam, Q.U. Zaman, S.I. Zamir, I. Khan and M.J. Subhani, 2014. Performance of different cultivars in direct seeded rice (Oryza sativa L.) with various seeding densities. Am. J. Plant Sci., 5: 3119-3128.
CrossRefDirect Link - Yang, J. and J. Zhang, 2010. Grain-filling problem in 'super' rice. J. Exp. Bot., 61: 1-5.
CrossRefDirect Link - Afiukwa, C.A., J.O. Faluyi, C.J. Atkinson, B.E. Ubi, D.O. Igwe and R.O. Akinwale, 2016. Screening of some rice varieties and landraces cultivated in Nigeria for drought tolerance based on phenotypic traits and their association with SSR polymorphism. African J. Agric. Res., 11: 2599-2615.
CrossRefDirect Link - Sharma, D., G.S. Sanghera, P. Sahu, P. Sahu and M. Parikh et al., 2013. Tailoring rice plants for sustainable yield through ideotype breeding and physiological interventions. Afr. J. Agric. Res., 8: 5004-5019.
Direct Link - Swain, P., M. Anumalla, S. Prusty, B.C. Marndi and G.J.N. Rao, 2014. Characterization of some Indian native land race rice accessions for drought tolerance at seedling stage. Aust. J. Crop Sci., 8: 324-331.
Direct Link - Rahman M.S., H. Miyake and Y. Takeoka, 2001. Effect of sodium chloride salinity on seed germination and early seedling growth of rice (Oryza sativa L.). Pak. J. Biol. Sci., 4: 351-355.
CrossRefDirect Link