
Sanjay Kumar Jaiswal
Microbiology Laboratory, Department of Genetics and Plant Breeding, Institute of Agricultural Sciences, Varanasi, India
LiveDNA: 91.3371
Akhil Anand
Microbiology Laboratory, Department of Genetics and Plant Breeding, Institute of Agricultural Sciences, Varanasi, India
Akhouri Vaishampayan
Microbiology Laboratory, Department of Genetics and Plant Breeding, Institute of Agricultural Sciences, Varanasi, India
Journal of Plant Sciences
Year: 2017 | Volume: 12 | Issue: 1 | Page No.: 30-38







ABSTRACT
Background and Objective: In agricultural eco-systems, microorganisms play a vital role in nutrient recycling through mobilization and solubilisation of mineral nutrient and biological nitrogen fixation. Symbiotic relation of soil microbes and legumes offer an alternative source to chemical fertilizer for plant growth and yield. The aim of the study was to assess the ability of soybean genotypes to nodulate with indigenous rhizobial inoculants. Materials and Methods: Field experiment was performed in four soybean cultivars with different rhizobial combinations in 2 experiments. In first experiment, soybean cultivar JS335 was inoculated with six rhizobia and two doses (30 and 60 kg) of N-fertilizer separately, while in second experiment evaluation of five isolates with three soybean cultivars were done in order to observe the effect of different inoculation and cultivar combination. The Data were analysed with one- and two-way analysis of variance (ANOVA) separately for first and second experiment, respectively. Results: The results showed significant difference (p<0.001) in symbiotic performance among the inoculants associated with soybean cultivars JS335. All test isolates were more effective and enhance 18.22-29.21% grain yield as compared to uninoculated control. Except isolate ALSR3, all test isolates enhanced 4.6-14.4% grain yield as compared to 60 kg ha–1 N-fertilizer. Soybean cultivar and inoculation combinations showed effective nodulation which were significantly different (p<0.001) from the uninoculated plots. The symbiotic parameters were much greater with cultivars VL47 and DS9712 in terms of nodule number, nodule dry weight and pod numbers plant. But non-significant difference was observed in grain yield. Conclusion: It was concluded that taken together, the symbiotic performance and grain yield data showed cultivar and inoculant specific effects and suggest that selection of rhizobial strains must be appropriate for the cultivars and newly released varieties.
PDF Abstract XML References Citation
Received: August 08, 2016;
Accepted: November 18, 2016;
Published: December 15, 2016
How to cite this article
Sanjay Kumar Jaiswal, Akhil Anand and Akhouri Vaishampayan, 2017. Response of Soybean (Glycine max L. Merr) Cultivars to Soil Microsymbionts Inoculation in Agriculture Field in India. Journal of Plant Sciences, 12: 30-38.
DOI: 10.3923/jps.2017.30.38
URL: https://scialert.net/abstract/?doi=jps.2017.30.38
DOI: 10.3923/jps.2017.30.38
URL: https://scialert.net/abstract/?doi=jps.2017.30.38
INTRODUCTION
Soybean has played a significant contribution to yellow revolution in India and as a food plant it forms an important part of the routine diet of the country1. India (10.21 Mt) occupies 5th position after USA (91.41 Mt), Brazil (56.96 Mt), Argentina (30.99 Mt) and China (14.5 Mt) in total production of soybean. However, the average yield of soybean in India is lower (1.1 t ha–1) as compared to the world average (1.8 t ha–1) and improving the current status of crop performance is a major challenge for India2.
In a low input agriculture like India, rhizobial inoculation gives a major benefit in the farming system, where recurrent soil erosion is occurring by the need to produce more and more food for feeding in increasing population and where chemical fertilizers are too expensive for the poor and marginal farmers. Biological nitrogen fixation is second only to photosynthesis as the most important biochemical process on earth. It can provide substantial amounts of N2 to plants and soil, which reduces the need for industrial fertilizers3.
Owing 40-45% protein in seeds soybean requires a high nitrogen supply. Symbiotic Nitrogen Fixation (SNF) resulting from mutual beneficial interaction between soybean and soil nodule bacteria (rhizobia) provides 60-70% of total nitrogen requirement of the plant. Population sizes of indigenous rhizobia compatible with the legume crop of choice are often very low in soils. Inoculation of legume seeds with rhizobia is perhaps the oldest agro-biotechnological application. Inoculants are generally produced regionally or locally, making use of selected bacterial strains and carrier materials available in that area.
Traditionally, isolation, testing and selection of single strains with desired properties (high N2 fixation efficiency with host plant) approach were applied for inoculants development. But still selection of strains is hampered even through the cracking of nodulation and N2 fixation genes in rhizobia4. Although rhizobial inoculants have been widely used for enhancing soil-based productivity, the economic potentiality of the inoculation technology is not yet fully realized because of inoculation failures. These occur when the bacterial population is too low or fails to grow or become established rapidly in the soil and rhizosphere5. There are several constraints to practical realization of this inoculation technology. The genetic instability of the inoculants in soils6, considerable reduction in numbers of inoculant strain7 and sensitivity to abiotic and biotic stresses constitute some of the constraints8.
Differences with respect to nitrogen fixation among various Rhizobium strains nodulating different host plants have been well stated9 and interactions of both rhizobial strain and host plant genotype have also been documented10-14. Thus, in SNF not only the microsymbiont (Rhizobium) but also the macrosymbiont (host plant) plays an important role and their co-selection under a given set of soil and environmental conditions may enhance the amount of fixing nitrogen and seed yield. Selection of host-cultivar compatible inoculants has been recognized as an important method for increasing nitrogen fixation in soybean15. It was observed that the SNF in legume depends upon co-selection of plant and bacterium genotypes16-18. Therefore, the present study was undertaken to identify soybean cultivars and brady rhizobial strains, which are superior in nodulation and plant development under field conditions.
MATERIALS AND METHODS
Study sites and sample collections: Soil and fresh root nodules were collected from soybean cultivated farmers, fields of different districts of Madhya Pradesh, Uttar Pradesh and Maharashtra provinces of India in August, 2010 and 2011. With the spade, a circle with radius of approximately 15 cm was described around the soybean plant and this section was cut out to a depth of at least 20 cm. The soil clump along with the plant was slowly lifted out with the help of spade. The soil was carefully removed from the root material, avoiding detachment of the secondary roots and nodules. The roots were placed into plastic bags. Soil samples were analyzed for their physical and chemical properties according to the procedure described by Jackson19. Collection site and characteristics of the soils used in this study are listed in Table 1.
Isolation of native rhizobia: Roots were washed thoroughly to remove soil. Nodules were washed with distilled water and surface sterilized with acidified mercuric chloride solution (0.1% w/v) for 1 minute followed by 70% alcohol for 1 min. Nodules were then thoroughly washed with sterile distilled water for 4-5 times. Each nodule was crushed separately with the help of a sterilized forceps and on the glass slide. A loopful of nodule exudates was streaked on Yeast mannitol agar medium plate with the help of inoculation needle and plates were kept in incubator at 28±1°C for 10 days.
Authentication of rhizobial isolates: All the rhizobial isolates were tested for nodule formation on soybean cv. JS 335 under laboratory condition as procedure described by Jaiswal et al.20. The rhizobial isolates, forming nodules in soybean were maintained on agar slants.
| Table 1: | Collection and soil types of soybean growing fields of three states of India used for isolation of root nodule bacteria |
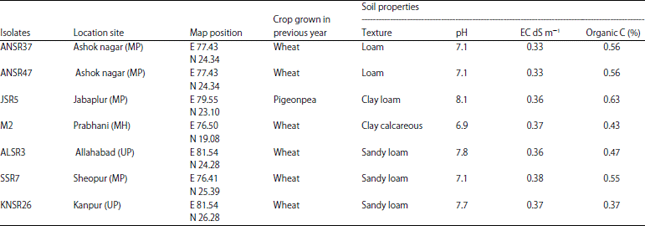 | |
| Parenthesis denotes state’s (MP: Madhya Pradesh, UP: Uttar Pradesh, MH: Maharashtra | |
Field design and planting: The 52 experiments were laid out in CRBD at Agricultural Research Farm, Banaras Hindu University, Varanasi, India, during Rabi season. In first experiment, treatments were consisted of seven rhizobial strains (ANSR37, ANSR47, JSR5, M2, SSR7, KNSR26 and ALSR3), two levels (30 and 60 kg ha–1) of nitrogen fertilizer and un-inoculated control with soybean cultivar JS335 in four replications. The second experiment was conducted with three soybean cultivars VL47, PK1042 and DS9712 in four replications with treatment comprised five rhizobial strains (ANSR37, ANSR47, JSR5, M2 and ALSR3) and un-inoculated control. The plot size was measured4 m×3 m (12 m2) with 15 cm plant-to-plant distance and row-to-row distance of 30 cm. The seeds were wrapped with sterilized charcoal containing single strain inoculums (3.7×108-7.3×108) with gum arabica as an adhesive before sowing. Data pertaining to nodule number, nodule fresh weight, nodule dry weight and plant dry weight were recorded after 45 days of sowing and pod number plant–1 along with grain yield data was recorded at the time of harvest.
Statistical analysis: The symbiotic data were analysed as one-way and two-way (ANOVA) using STATISTICA 8.0 software program21. Duncan’s Multiple Range Test (DMRT) was applied to separate the mean to know significant difference (p<0.05) between treatments21.
RESULTS
A total of 112 rhizobial isolates were isolated from collected nodules of soybean grown in farmers’ field of different districts in 3 states (Madhya Pradesh, Uttar Pradesh and Maharashtra) of India. The field soils were loamy, sandy loamy and clay loamy with pH varying from 6.9-8.1. Out of 112, 90 bacterial isolates were able to form nodule with the homologous plant in control conditions. Variation in duration for nodule appearance was observed with different rhizobial isolates (data not shown). Isolates ANSR37, ANSR47, JSR5, M2, SSR7, KNSR26 and ALSR3 were selected for field trial for selecting out the varietal and isolate combinations.
A one-way analysis of data collected from first experiment of field trial for the treatments different levels of nitrogen (30 and 60 kg ha–1 Urea), symbiotic response of isolates ANSR37, ANSR47, JSR5, M2, SSR7, KNSR26 and ALSR3 with soybean cultivar JS335 revealed significantly marked difference in nodule number, nodule fresh weight, nodule dry weight, plant dry weight and grain yield. In this result, symbiotic parameters of soybean cultivar JS335 with test isolates’ treatment were much higher as compared to nitrogen doses (Table 2). Significant differences (p<0.001) were observed in symbiotic parameters i.e. nodule number (9-37 nodules plant–1), nodule fresh weight (58-238 mg plant–1), nodule dry weight (25-101 mg plant–1) and plant dry weight (11.4-13.35 g) by the application of test isolates to soybean genotype JS335 relative to their uninoculated and nitrogen doses of urea (30 and 60 kg ha–1). The treatment of isolate ANSR 47 showed the highest symbiotic parameters in terms of nodule number, nodule fresh weight, nodule dry weight and plant dry weight. The highest grain yield was observed by the treatment of SSR 7 followed byANSR37 and M2 (Table 2). The average nodule number and dry weight of nodules at 45 days after planting were 1 and 1.25 mg plant–1 in control, but greatly increased by the test Rhizobium treatments (Table 2). The dry weight plant–1 accumulation was low in uninoculated plot and significantly increased (p<0.001) by all test inoculations. Grain yield of uninoculation was (12.09 q ha–1) was lower than N fertilizer.
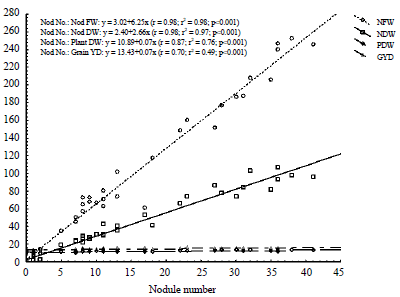 | |
| Fig. 1: | Correlation analysis of symbiotic parameters. Relationship of Nodule number (Nod No.) with nodule fresh weight (N FW), Nodule dry weight (N DW), Plant dry weight (PDW) and Grain yield (GYD) of soybean cultivar JS335 |
| Table 2: | Effect of different N-doses and inoculation response with soybean cultivar JS335 under field conditions |
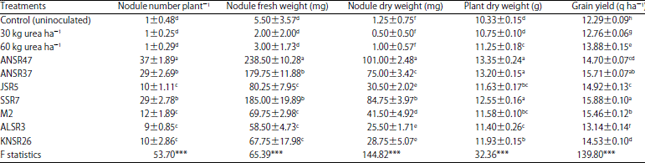 | |
| Values (Mean±SE) with dissimilar letters in a column are significantly different at p<0.001 (***) | |
However, all the test inoculants significantly increased grain yield from 18.22-29.21% in compare to uninoculated control and 4.6-14.4% more than N-fertilizer (Table 2).
Correlation and regression analysis showed a significant positive relationship of nodule number with nodule fresh weight (r = 0.98***), nodule dry weight (0.98***), plant dry weight (0.87***), grain yield (0.70***) (Fig. 1).
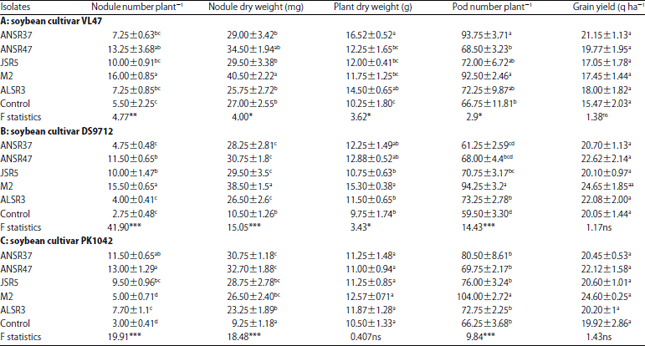
Symbiotic performance of soybean genotypes with different rhizobial isolates: The selected isolates showed induction of nodules on the test soybean cultivars. The two-way ANOVA showed that host cultivars and inoculation treatment as well as interaction had significant (p<0.05) effect on nodulation, nodule dry weight (Table 3). The one-way analysis was also done to see the effect of rhizobial isolates on soybean cultivar individually.
One-way analysis of rhizobial isolates and cultivar
Soybean cultivar VL47: The average nodule number plant–1 ranged from 6 (control)-16 (M2) with soybean cultivar VL47. Significantly higher (p<0.01) nodule number plant–1 was induced by rhizobial isolates M2 (16 nodules plant–1) and ANSR47 (14 nodules plant–1). Nodule number, nodule dry matter, plant dry weight (DW) and pod number plant–1 with soybean cultivar VL 47 varied significantly among the isolates (Table 4). Plant DW ranged from 11.75 g plant–1 with the isolate M2 to 16.52 g plant–1 with ANSR37 (Table 4). Nodule dry weight varied from 25.75-40.5 mg plant–1 with isolates ALSR3 and M2, respectively. The inoculation with isolate ANSR37 produced the highest pod number plant–1 as compared to others. There was no significant difference observed in grain yield in VL47 inoculated with test isolates but isolate ANSR37 inoculated to VL47 yielded ~37% more grain yield in compare to uninoculated control.
| Table 3: | A two-way (ANOVA) comparison of different soybean cultivars and microsymbionts for symbiotic relations in field conditions |
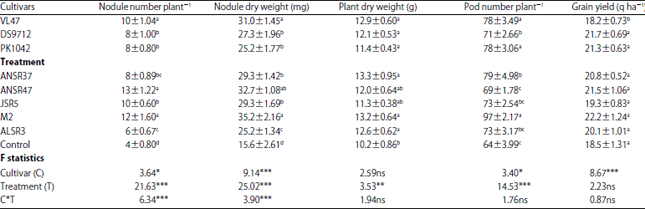 | |
| Values (Mean±SE) with dissimilar letters in a column are significantly different at p<0.05 (*), p<0.01 (**), p<0.001 (***) | |
| Table 4: | A one-way comparison in symbiotic performance of soybean cultivars with rhizobial isolates |
 | |
| Values (Mean±SE) with dissimilar letters in a column are significantly different at p<0.05 (*), p<0.01 (**), p<0.001 (***) | |
Soybean cultivar DS9712: The isolates were assessed for symbiotic parameter in terms of nodule number, nodule dry weight, Plant dry weight, pod number plant–1 and grain yieldwith soybean cultivar DS9712. Of these five isolates, ANSR47 (12 nodule plant–1), JSR5 (10 nodule plant–1) and M2 (12 nodule plant–1) produced much higher nodule number compared to ANSR37 and ALSR3 (4 nodule plant–1). Plant DW plant–1 was highest with isolate M2, followed by isolates ANSR47, ANSR37, ALSR3 (Table 4b). Isolate M2 had the highest pod number plant–1 also, followed by isolates ALSR3, JSR5 and ANSR47. Like VL47, there was no significant difference observed in seed yield in soybean cultivar DS9712 inoculated with test isolates (Table 4).
Soybean cultivar PK1042: With soybean cultivar PK1042, plant dry weight plant–1 and seed yield werenot significantly different with all test isolates. Isolate M2 produced the least nodule number plant–1 but the highest pod number plant–1 (104) while isolate ANSR47 formed the highest nodule (13 plant–1) but the least pod number (70 plant–1). Like VL47 and DS9712, no significant difference was observed in grain yield in soybean cultivar PK1042 inoculated with test isolates but isolate M2 was able to enhance 23.5% more yield in comparison to uninoculated control (Table 4).
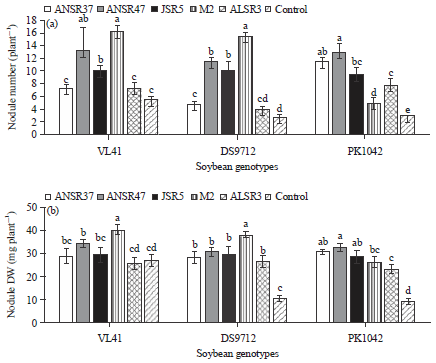 | |
| Fig. 2(a-b): | Soybean cultivars Rhizobium interaction, (a) Nodule number/plant and (b) Nodule dry weight (mg) |
Two-way ANOVA comparison of symbiotic performance in soybean cultivars and test isolates: A combined two-way ANOVA, comparison of the three soybean cultivars (VL47, DS9712 and PK1042) grown with five test isolates was done (Table 3).
The symbiotic response of soybean cultivars to indigenous rhizobia was assessed and showed a marked inoculation response on nodule number (Fig. 2a) and nodule dry weight plant–1 (Fig. 2b).
Soybean cultivar VL47 exhibited greater nodule number (10 nodule plant–1), nodule dry weight (31mg plant–1), plant dry weight (12.9 g plant–1) and pod number (78 plant–1) when compared to other cultivars (Table 3). There was no significant difference in plant dry weight observed.
Of the six treatments including control (5 isolates+1 uninoculation) tested with three soybean cultivars, isolate ANSR47 produced the highest nodule number (13 plant–1). The isolate M2 reported the highest nodule dry weight (35.2 mg plant–1) and pod number (97 plant–1) which indicated highest grain yield. The highest dry weight plant–1 was recorded for isolate ANSR37 (13.3 g plant–1).
The soybean cultivar×isolate interaction was significantly increased (p<0.001) for nodule number plant–1 and nodule dry weight plant–1 (Table 3). In general, inoculation increased nodule number plant–1, nodule dry weight plant–1, plant dry weight and pod number plant–1 in all test soybean cultivars when compared to their uninoculated control. The nodule number plant–1 was greater in interaction of isolate M2 with VL47 and DS9712 while isolate ANSR47 showed highest nodule plant–1 with soybean cultivar PK1042 (Fig. 2a). Nodule number for other isolates with soybean cultivars was also significantly different. As a result of difference in nodule number, nodule dry weight was also higher in soybean cultivars VL47 and DS9712 with M2 isolate (Fig. 2b).
Correlation analysis: Whether dealing with different soybean genotypes, nodule dry weight plant–1 was significantly correlated with pod number plant–1 (r = 0.27*) and dry weight plant–1 (r = 0.25*) (Fig. 3a). A significant correlation (r = 0.30**) was also observed between pod number and dry weight plant–1 (Fig. 3b). The high significant (r = 0.69***) correlation was observed between nodule number and nodule dry weight plant–1 while low significant correlation (r = 0.23*) was observed between pod number and nodule number plant–1 (Fig. 3c).
DISCUSSION
The data presented in 1st experiment reported that native rhizobial strains were symbiotically more effective on test soybean cultivar JS335 than the uninoculated and N-fertilizer doses. Results showed that inoculation with rhizobial isolates promoted large and significant increase (p<0.001) in all studied symbiotic parameters (nodule number, nodule dry weight, dry weightplant–1 and grain yield ha–1) (Table 2).
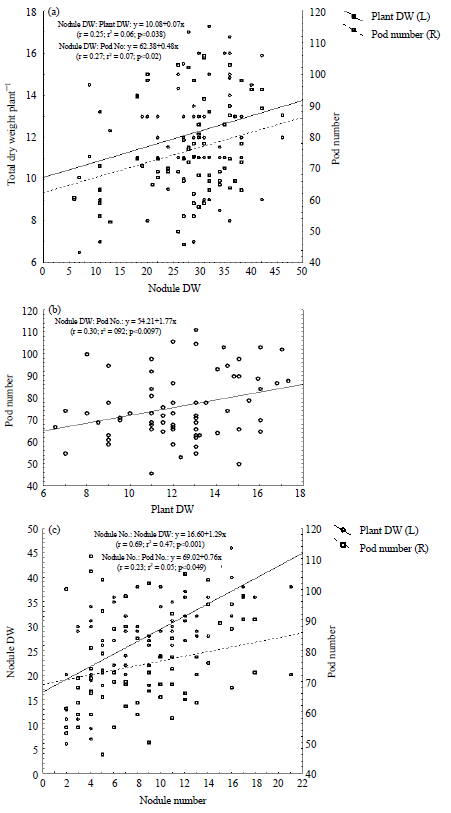 | |
| Fig. 3(a-c): | Correlation analysis of symbiotic parameters, (a) Relationship of nodule dry weight (nodule DW) with total dry weight plant–1 and pod number, (b) Pod number ([od No.) and plant dry weight (plant DW) and (c) Nodule number with nodule dry weight (nodule DW) and pod numbers (Pod No.) of test soybean cultivars |
In terms of grain yield all isolates were more effective and enhance 18.22-29.21% grain yield in compare to uninoculated control. Except isolate ALSR3, all test isolates enhanced 4.6-14.4% grain yield in compare to 60 kg ha–1 N-fertilizer. The results of isolate ALSR3 indicated that N derived from N2-fixation by this isolate promoted vegetative growth but did not increase grain yield in compare to 60 kg ha–1 N-fertilizer22. In earlier study at Embrapa Brazil it has also been study in soybean23.
It was reported here that the test soybean cultivars showed effective nodulation which are significantly different (p<0.001) from the uninoculated plots. Introduced inoculum played a major role for nodulation and grain yield in compared to control in studied location. The observed inoculation response for symbiotic parameters was much greater with cultivars VL47 and DS9712 in terms of nodule number, nodule dry weight and pod number. The presence of nodule in uninoculated plot was taken as an indication of the presence of cultivars compatible effective rhizobia in studied soil location but low in number. Previous reports had similar finding that variations in plant growth, nodulation and N2-fixation of different soybean genotypes due to affectivity of inoculated microsymbionts24,25. Though inoculation significantly promoted symbiotic performance of all test soybean cultivars, grain yield varied among the test cultivars but not significantly different. The non-significant difference of grain yield could suggest that the native soybean rhizobia was just as effective as the introduced test isolates or soil factors reduced the multiplication rate and symbiotic efficacy of the test isolates. Cultivar VL47 showed higher nodule number plant–1, nodule dry weight and plant dry weight but low grain yield in compare to other cultivars, indicated that fixed nitrogen promoted large amounts of vegetative growth rather than grain yield22.
The studied soil site was followed which could be a reason of low native population of rhizobia in soil26 caused low nodule number with the cultivars in uninoculated plots but they have been very effective.
A comparison of symbiotic performance and grain yield data suggest cultivar-specific effects and attributed to genetic difference among the genotypes.
Results showed that the cultivars varied with respect to nitrogen fixation, which was in agreement with the results of Caldwell and Vest27 who, reported the influence of host plant on symbiotic effectiveness nodulation and nitrogen fixation. The obtained results reveal significant effect (p<0.001) in legume host×Rhizobium interaction for all the three cultivars which is parallel to the result of host×Rhizobium interaction by Bello et al.10, Rennie and Kemp11, Skot12, Shivananda et al.28, Musiyiwa et al.29, Yates et al.30.
CONCLUSION
Taken together, the results suggest that soybean cultivar can respond remarkably to inoculation with indigenous competitive strains. It is indicated that a good compatibility of soybean cultivars with suitable rhizobial strains are needed which convey the information that selection of rhizobial strains must be appropriate for the newly released varieties to select out an efficient rhizobial strain mutually compatible with each individual cultivar. Furthermore, in studies, there is need to identify those effective soybean compatible rhizobia from Indian soils.
SIGNIFICANCE STATEMENT
This study discovers that importance of inoculants in Indian climatic conditions that can be beneficial for soybean production. This study will help the researcher to uncover the critical areas of molecular signal between cultivars dependent microsymbionts relations that many researchers were not able to explore.
REFERENCES
- Chauhan, G.S. and O.P. Joshi, 2005. Soybean (Glycine max): The 21st century crop. Indian J. Agric. Sci., 75: 461-469.
Direct Link - Carlsson, G. and K. Huss-Danell, 2003. Nitrogen fixation in perennial forage legumes in the field. Plant Soil, 253: 353-372.
CrossRefDirect Link - Franche, C., K. Lindstrom and C. Elmerich, 2009. Nitrogen-fixing bacteria associated with leguminous and non-leguminous plants. Plant Soil, 321: 35-59.
Direct Link - Brockwell, J., P.J. Bottomley and J.E. Thies, 1995. Manipulation of rhizobia microflora for improving legume productivity and soil fertility: A critical assessment. Plant Soil, 174: 143-180.
Direct Link - Bromfield, E.S.P., A.M.P. Behara, R.S. Singh and L.R. Barran, 1998. Genetic variation in local populations of Sinorhizobium meliloti. Soil Biol. Biochem., 30: 1707-1716.
CrossRefDirect Link - Hatzinger, P.B. and M. Alexander, 1994. Relationship between the number of bacteria added to soil or seeds and their abundance and distribution in the rhizosphere of alfalfa. Plant Soil, 158: 211-222.
Direct Link - Streeter, J.G., 1994. Failure of inoculant rhizobia to overcome the dominance of indigenous strains for nodule formation. Can. J. Microbiol., 40: 513-522.
CrossRefDirect Link - Bello, A.B., W.A. Ceron-Diaz, C.D. Nickell, E.O. El Sheriff and L.C. Davis, 1980. Influence of cultivar, between-row spacing, and plant population of fixation of soybean. Crop Sci., 20: 751-755.
CrossRefDirect Link - Rennie, R.J. and G.A. Kemp, 1983. N2-fixation in field beans quantified by 15N isotope dilution. II. Effect of cultivars of beans. Agron. J., 75: 645-649.
CrossRefDirect Link - Skot, L., 1983. Cultivar and Rhizobium strain effects on the symbiotic performance of pea (Pisum sativum). Physiol. Planta., 59: 585-589.
CrossRefDirect Link - Jaiswal, S.K. and B. Dhar, 2011. Identification and assessment of symbiotic effectiveness of phage-typed Rhizobium leguminosarum strains on lentil (Lens culinaris Medik) cultivars. Curr. Microbiol., 62: 1503-1509.
CrossRefPubMedDirect Link - Anand, A., S.K. Jaiswal, B. Dhar and A. Vaishampayan, 2012. Surviving and thriving in terms of symbiotic performance of antibiotic and phage-resistant mutants of Bradyrhizobium of soybean [Glycine max (L.) Merrill]. Curr. Microbiol., 65: 390-397.
CrossRefDirect Link - Israel, D.W., J.N. Mathis, W.M. Barbour and G.H. Elkan, 1986. Symbiotic effectiveness and host-strain interactions of Rhizobium fredii USDA 191 on different soybean cultivars. Applied Environ. Microbiol., 51: 898-903.
Direct Link - Appunu, C. and B. Dhar, 2006. Differential symbiotic response of phage-typed strains of Bradyrhizobium japonicum with soybean cultivars. J. Microbiol., 44: 363-368.
PubMedDirect Link - Jaiswal, S.K., A. Anand, B. Dhar and A. Vaishampayan, 2012. Genotypic characterization of phage-typed indigenous soybean bradyrhizobia and their host range symbiotic effectiveness. Microb. Ecol., 63: 116-126.
CrossRefDirect Link - Boddey, R.M., M. Fosu, W.K. Atakora, C.H. Miranda, L.H. Boddey, A.P. Guimaraes and B.D. Ahiabor, 2016. Cowpea (Vigna unguiculata) crops in Africa can respond to inoculation with rhizobium. Exp. Agric.
CrossRefDirect Link - Neves, M.C.P., A.D. Didonet, F.F. Duque and J. Dobereiner, 1985. Rhizobium strain effects on nitrogen transport and distribution in soybeans. J. Exp. Bot., 36: 1179-1192.
CrossRefDirect Link - Pule-Meulenberg, F., C. Gyogluu, J. Naab and F.D. Dakora, 2011. Symbiotic N nutrition, bradyrhizobial biodiversity and photosynthetic functioning of six inoculated promiscuous-nodulating soybean genotypes. J. Plant Physiol., 168: 540-548.
CrossRefDirect Link - Salvagiotti, F., K.G. Cassman, J.E. Specht, D.T. Walters, A. Weiss and A. Dobermann, 2008. Nitrogen uptake, fixation and response to fertilizer N in soybeans: A review. Field Crops Res., 108: 1-13.
CrossRefDirect Link - Raverkar, K.P., G.S.B. Ajay and D.L.N. Rao, 2005. Survival of soybean rhizobia in summer and proliferation during monsoon in Vertisols of central India. J. Indian Soc. Soil Sci., 53: 591-597.
Direct Link - Musiyiwa, K., S. Mpepereki and K.E. Giller, 2005. Symbiotic effectiveness and host ranges of indigenous rhizobia nodulating promiscuous soyabean varieties in Zimbabwean soils. Soil Biol. Biochem., 37: 1169-1176.
CrossRefDirect Link - Yates, R.J., J.G. Howieson, W.G. Reeve, L. Brau and J. Speijers et al., 2008. Host-strain mediated selection for an effective nitrogen-fixing symbiosis between Trifolium spp. and Rhizobium leguminosarum biovar trifolii. Soil Biol. Biochem., 40: 822-833.
CrossRefDirect Link