
K. Khucharoenphaisan
Department of Biology, Faculty of Science and Technology, Phranakhon Rajabhat University, Bangkok, 10220, Thailand
LiveDNA: 66.1915
K. Rodbangpong
Department of Biology, Faculty of Science and Technology, Phranakhon Rajabhat University, Bangkok, 10220, Thailand
P. Saengpaen
Department of Biology, Faculty of Science and Technology, Phranakhon Rajabhat University, Bangkok, 10220, Thailand
K. Sinma
Department of Soil Science, Faculty of Agriculture, Kasetsart University, Kamphaeng Saen Campus, Nakhon Pathom, 73140, Thailand
Journal of Plant Sciences
Year: 2016 | Volume: 11 | Issue: 1-3 | Page No.: 38-44







ABSTRACT
Actinomycetes have been promised as biocontrol and stimulating agents for use in agriculture without detrimental effects to the environmental due to their antifungal with secondary metabolites produced. The aim of this study was to screen soil actinomycetes according to its ability to produce various secondary metabolites against Phytophthora sp. that causing root rot disease of cassava and stimulating agent has also determined. Firstly, soil actinomycetes were isolated and tested for antagonistic activity toward the fungus by the dual culture technique. After that the selected isolate was determined on the stimulating agent as IAA production. Finally, extracellular anti-fungal metabolites produced by selected isolates were evaluated for anti-fungal potential toward the fungus with agar core technique. The result showed that 98 isolates from soil samples were screened on their anti-fungal activity. Among these, 38 isolates showed the inhibition activities against Phytophthora sp. in which was isolated from infected cassava. The culture supernatants without cell obtained from 16 isolates were affective against the fungus whereas 10 isolates produced affective thermostable compound. In total, the isolate LB35 was most promising on the basis of its interesting antimicrobial activity and could produce IAA with 50 μg mL–1. Based on its 16S rDNA sequence and phylogenetic tree analysis, isolate LB35 belong to the Streptomyces malaysiensis. The addition of isolate LB35 as fresh inoculums to the cassava field, the cassava showed more height comparing to control experiment. Moreover, S. malaysiensis has not to be phytopathogenic microorganism of cassava. This finding has been increased scope of agriculturally important actinomycetes applications.
PDF Abstract XML References Citation
Received: November 04, 2015;
Accepted: December 19, 2015;
Published: April 15, 2016
How to cite this article
K. Khucharoenphaisan, K. Rodbangpong, P. Saengpaen and K. Sinma, 2016. Exploration on Soil Actinomycetes Against Phytophthora sp. Causing Root Rot of Cassava and Plant Growth Promoting Activities. Journal of Plant Sciences, 11: 38-44.
DOI: 10.3923/jps.2016.38.44
URL: https://scialert.net/abstract/?doi=jps.2016.38.44
DOI: 10.3923/jps.2016.38.44
URL: https://scialert.net/abstract/?doi=jps.2016.38.44
INTRODUCTION
Actinomycetes play an important role in agriculture in term of biocontrol. Antagonism from these microorganisms against an extensive variety of phytopathogenic fungi has been reported (Doumbou et al., 2001; Khucharoenphaisan et al., 2013, 2014). Among species of those belonging to the genus Streptomyces and secreted a wide biologically active compounds including antibiotics, hydrolytic enzymes such as chitinase (Taechowisan et al., 2009; Prapagdee et al., 2008; Sinha et al., 2014). Moreover, enhanced soil fertility and also proved to possess biological agents against wide range of soil-borne plant pathogens such as Alternaria, Fusarium, Macrophomina, Phytophthora, Pythium, Rhizoctonia, Verticillium (Aghighi et al., 2004; Sinha et al., 2014). These actinomycetes may be an interested source to finding efficient biocontrol agents for plant health and promoting growth through secretion of plant growth regulators. Actinomycetes were found to produce a variety of metabolite including plant growth regulating agent such as auxin, gibbellin and Indole Acetic Acid (IAA) (Bloemberg and Lugtenberg, 2001). Various genera of Actinomycetes had ability to produce IAA such as Nocardiopsis jiangxieusis and Actinomadura glauciflara (Sharma et al., 2013; Khamna et al., 2009).
Cassava was raw material in various industrial processes including cassava starch production and monosodium glutamate industrial (Buensanteai and Athinuwat, 2012; Barros et al., 2014). In Thailand, it was considered as one of the main crops for ethanol production (Chaisinboon and Chontanawat, 2011). The production has increased drastically every year according to demand of consumer (Adeoti, 2010). The important factors affected to plant health are the microorganisms living in and out-side of the plant (Hirscha and Valdesc, 2010). However, the quality of cassava was affected by many factors such as fungal disease resulting to low levels of productivity (Fukuda, 1991). The root rot disease has been responsible for major losses in crop. The fungi caused root rot disease of cassava mainly Phytophthora spp. and Fusarium solani (Fukuda, 1991). To protect them from diseases and stimulate growth of the plant, the control strategies are the use of resistant variety and the application of fungicide. However, the control using chemical agent leads to environmental impact. The environmental friendly application was consider controlling the cassava disease and stimulate growth. The aim of this study was screening of 98 soil actinomycetes against Phytophthora sp. causing of cassava root rod disease. The culture broth of those with and without heat treatment was also investigated for testing activity. Moreover, the abilities of those isolates for IAA production were determined. Field experiments were done for the efficiency of selected actinomycetes strain against pathogenic fungi and stimulated growth of cassava.
MATERIALS AND METHODS
Soil sampling: Soil samples were collected from cassava fields of 10 different crop systems in tropical dry sandy in 3 provinces of Thailand, Lopburi, Nakhon Pathom and Kanjanaburi. The samples were kept at 4°C until use.
Isolation of soil actinomycetes and phytopathogenic fungus: Actinomyces were isolated from cassava field soil. Each sample was diluted to go on serial dilution and plate on humic acid vitamin agar (Hayakawa and Nonomura, 1987) supplemented with 50 mg L–1 of cyclohexamide and 20 mg L–1 of nalidixic acid. The isolation plates were incubated at 35±2°C for 7 days. The colonies had been transferred to International Streptomyces Project (ISP) medium No. 2 agar (Shirling and Gotlieb, 1966; Khucharoenphaisan and Sinma, 2011) plates for purity check.
Phytophthora sp. causing root rot of cassava was isolated from infected cassava. The isolated fungus was cultured on V8 agar at 35±2°C. Morphology under microscope showed mycelium with sporangium. It was identified as Phytophthora sp. using Internal Transcribed Spacer (ITS) region sequencing (Boonlue et al., 2003).
Screening of antifungal activity of actinomycetes: Ninety eight strains of soil actinomycetes were cultured on ISP medium No. 2 agar. The cultures of those actinomycetes were single streak on one side of PDA (potato dextrose agar) at distance of 1.0 cm from edge of plate and then incubated at 35±2°C for 7 days. The active mycelium of Phytophthora sp. was cut and transferred to the middle of cultured actinomycetes plate. The inhibition of mycelium growth was measured after 7-day cultivation. The inhibition was evaluated by radius of fungal colony compared to control culture. Three replications were done.
Test of culture filtrates: The effective soil actinomycetes were inoculated in ISP medium No. 2 broth and then incubated at 35±2°C for 7 days with shaking condition at 150 rpm. The cultured supernatants were filtrated using sterile cotton as supernatant. The supernatant with and without heat treatment at 100°C for 15 min were tested against phytopathogenic fungus of cassava activity using agar well method. The active mycelium of Phytophthora sp. was cut and transferred to the middle of cultured actinomycetes plate. The fungal plate was incubated at 35±2°C for 5 days. Thirty microliter of supernatant with and without heat treatment was put to each well and then inhibition of mycelium growth was measured. Three replications were done.
Identification of selected actinomycetes: The selected actinomycetes were tentative identified into genera based on morphological characteristics using crosshatch streak technique (Shirling and Gottlieb, 1966; Khucharoenphaisan et al., 2011). The morphological characteristics were observed as colony shape, color, sporulation, soluble pigment on ISP No. 2 and humic acid vitamin agar after 14-day cultivation at 35±2°C (Cuesta et al., 2012).
The 16S rDNA amplification of actinomycetes was prepared by PCR using universal primer 9F (5’-GAGTTTGATCC TGGCTCAG-3’) and 1541R (5’-AAGGAGGTGATCCAGCC-3’). The PCR products were purified and directly sequence using a Big Dye Terminator V3.1 cycle sequencing kit (Applied Biosystems) and the universal primers 9F (5’-GAGTTTGATCCTGGCTCAG-3’), 785F (5’-GGATTAGATACCCTGGTAGTC-3’), 802R (5’-TACCAGG GTATCTAATCC-3’) and 1541R (5’-AAGGAGGTGATCCAGCC-3’) (Khucharoenphaisan et al., 2012). The nucleotide sequences were compared with other bacteria using the Genetyx version 5.0. The phylogenetic tree was constructed by using the neighbor-joining method in MEGA version 4 software. The topology was evaluated by bootstrap analysis based on 1000 resamplings (Felsenstein, 1985).
Indole acetic acid (IAA) production: Selected soil actinomycetes was determined for its IAA production using colorimetric assay according to the methods of Goudjal et al. (2013) and Gangwar et al. (2012). The actinomycetes were grown on yeast malt extract broth containing 0.2% L-tryptophan and incubated at 35±2°C with shaking at 150 rpm for 5 days. Cultures were centrifuged 9000 rpm for 5 min. One milliliter of supernatant mixed with 2 mL of the Salkowski reagent. Absorbent was read at 530 nm using spectrophotometer. The level of IAA production was estimated by comparison with IAA standard.
Production of inoculums: The inoculum was prepared by placing 100 g of rice grain into plastic bag and autoclaving at 121°C for 15 min and then inoculated with S. malaysiensis or Phytophthora sp. and incubated at 35±2°C for 1 week after that mix with coconut dust in ratio 1:1. The sterile rice grain and coconut dust without inoculation were used as control using in the cassava fields experiment.
Fields experiment: The inoculum was dispersed in the soil after planting cassava for 2 month. In total, there were four treatment combinations: (T1) Control without microbial inoculation (T2) Inoculation with S. malaysiensis (T3) Inoculation with S. malaysiensis and Phytophthora sp. and (T4) Inoculation with Phytophthora sp. The efficiency of actinomyces was monitored by growth of the cassava. Twenty four cassavas per treatment were investigated.
Data analysis: The student t-test was used to determine significance fungal inhibition by supernatant and living cell of actinomycetes.
RESULTS AND DISCUSSION
Antifungal activity of actinomycetes: Among 98 isolates of actinomycetes from cassava field soils, 38 isolates showed antifungal activity against Phytophthora sp. as more than 60% (Fig. 1 and Table 1). All effective actinomyces were cultured in liquid medium and supernatants were collected for inhibition test.
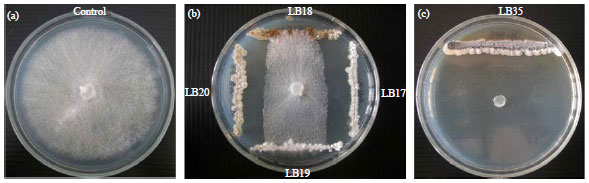 | |
| Fig. 1(a-c): | Colony of Phytophthora sp. isolates from infected cassava as a (a) Control and (b, c) Antifungal activity of actinomycetes stains after incubated on PDA at 35±2°C for 7 days |
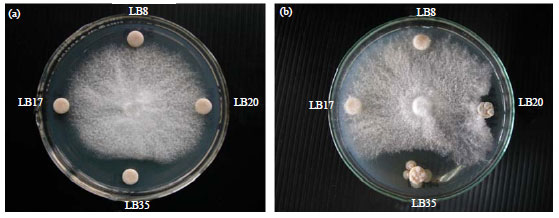 | |
| Fig. 2(a-b): | Anti-fungal activity of actinomycetes culture supernatant (a) With and (b) Without heat treatment at 100oC for 15 min against Phytophthora sp. |
| Table 1: | Anti Phytophthora sp. activities of actinomycetes supernatant with and without heat treated at 100oC for 10 min |
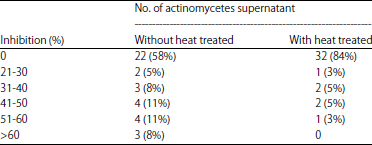 | |
The result showed that supernatants from 16 strains had significantly inhibited the Phytophthora sp. grown on agar plate. However, there are only 6 isolates producing effective supernatants with heat resistant at 100oC for 15 min (Fig. 2). This due to supernatant without heat treatment contained living cell of actinomycetes and it produced secondary metabolites or hydrolytic enzyme against the phytopathogenic fungus. Among these, isolate LB35 gave the highest inhibition of Phytophthora sp. in both supernatants with and without heat treatment. This indicated that isolate LB35 produced effective secondary metabolites with heat resistant such as bioactive compound to inhibit the phytopathogenic fungal growth. Some isolate of actinomyces had effective against the phytopathogenic fungal growth when use as cell or supernatant whereas supernatant with heat treatment were not. One of possible reason was the production of hydrolytic enzymes such as chitinase. Another one, the actinomycetes produced effective secondary metabolites to inhibit the phytopathogenic fungal growth but it not be heat resistant.
The finding agreed with the report of Suwan et al. (2012) that Streptomyces sp. isolated from soil that collected from Suthep-Pui National Park, Thailand have effective activity against phytopathogenic fungus. Moreover, Sinha et al. (2014) reported that Streptomyces plicatus was isolated from the soil could inhibit plant pathogenic fungi Phytophthora infestans and Sclerotium rolfsii to the tune of 80-100%. Elamvazhuthi and Subramanian (2013) also isolated Streptomyces from paddy soils and their antagonistic activity against Rhizoctonia solani, Helminthosporium oryzae, Curvularia lunata and Fusarium orysporum by using dual plate assay.
Production of indole-3-acetic acid (IAA): Among 6 isolates producing effective supernatant with heat resistant at 100°C for 15 min. Only strain LB35 could produce IAA with 50 μg mL–1. Generally, IAA were effected to plant growth stimulating and controlling the mechanism of cell division, cell elongation and cell differentiation resulted to decrease of root length while increase number of root hair for more uptake of nutrient (Peciorek and Friml, 2006; Dobbelaere et al., 1999; Bennett et al., 1998). Various genera of Actinomycetes had ability to produce IAA such as Nocardia jiangxiensis and Actinomadura glauciflara, Aquilaria crassawa and it can be induce by the medium containing tryptophane (Sharma et al., 2013). However, Streptomyces malaysiensis has not been reported.
Molecular identification: The 16S rDNA sequence was generated for strain LB35 (1416 nucleotides). The phylogenetic tree of species Streptomyces constructed by Neighbor-joining method from MEGA4 program. The tree is rooted by the nucleotide sequence of Bacillus subtilis AE4-3. Scale bar shown distance values under the tree means 0.02 substitutions per nucleotide position. Bootstrap analyses were performed with 1000 re-samplings and percent values (>50) are shown at the branching points. Comparison of this nucleotide sequence with members of actinomycetes clearly showed that this strain belong to the genus Streptomyces (Fig. 3). The close relationship to Streptomyces malaysiensis is supported by both of treeing algorithms and a high bootstrap value (Fig. 3).
Application of strain LB35 on cassava field: The cultures of S. malaysiensis LB35 and Phytophthora sp. was applied on the 2 month growth cassava in the field, except negative control. The effect of Phytophthora sp. and S. malaysiensis LB35 on cassava growth on the field were checked every 2 months. After 4 month of planting (Fig. 4), treatment 1 (T1) Phytophthora sp. and S. malaysiensis LB35 were not added to cassava field. The cassava had height of 131.29±8.27 cm (Table 2). Addition only S. malaysiensis LB35 to cassava field, the cassava showed the highest height of 143.92±10.42 cm as shown on treatment 2 (T2). After added Phytophthora sp. and S. malaysiensis LB35 to cassava field (treatment 3, T3), the growth of cassava in term of height was slow decrease at 136.21±6.98 cm. In treatment 4 (T4) which inoculated only Phytophthora sp. in the cassava field revealed the growth similar to control experiment (T1). This indicated that S. malaysiensis LB35 can be use to stimulate growth of cassava because it produced IAA and also produce biological agent to inhibit Phytophthora sp. phytopathogenic fungus that cause of root rot.
Streptomyces malaysiensis has not been reported to be phytopathogenic microorganisms. Furthermore, it can produce bioactive compound to inhibit phytopathogenic fungus. Li et al. (2008) reported that S. malaysiensis produced malayamycin against Stagonospora nodorum causing of stagonospora nodorum blotch of wheat.
| Table 2: | Growth of cassava after planting with Streptomyces malaysiensis LB35 and Phytophthora sp. on the field at 4 month |
 | |
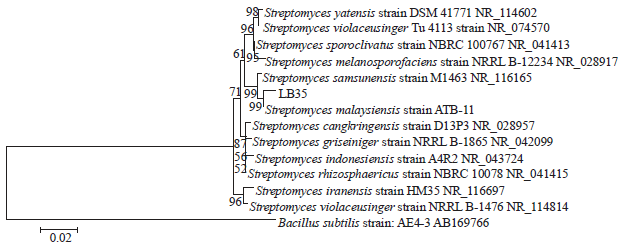 | |
| Fig. 3: | Phylogenetic tree of nucleotide sequence analysis of 16S rDNA of strain LB35 |
 | |
| Fig. 4(a-c): | Cassava growth in the fields (a) After 2-month and (b, c) 4-month planting |
Nonoh et al. (2010) isolated S. malaysiensis from national park soils in Kenya. It showed antagonistic activity to Fusarium oxysporum, Fusarium sp. and Colletotrichum kahawae. Streptomyces malaysiensis not only produced malayamycin, but also produced azalomycin F complex to inhibit phytopathogenic fungi such as Fusarium oxysporum, Rhizoctonia solani, Cladosporium cladosporioides, Fusarium chlamydosporum, Colletotrichum gloeosporioides (Cheng et al., 2010). Khucharoenphaisan et al. (2013) found that S. malaysiensis R58 produced thermostable bioactive compound on the culture broth against Colletotrichum gloeosporioides chi resulting to reduce chilli anthracnose.
CONCLUSION
The strain LB35 has high efficiency against phytopathogenic fungus Phytophthora sp. which isolated from infected cassava. This strain produced thermostable bioactive compound in the culture broth against Phytophthora sp. Moreover, it can produce IAA to stimulate growth of cassava in the field. Based on its 16S rDNA sequence, the actinomycetes strain LB35 is belonging to Streptomyces malaysiensis. Further study needs to be determined cassava production in the field. Moreover, it needed more potential bioactive antifungal compound and stimulating agents from S. malaysiensis, which are easily available in the soils.
ACKNOWLEDGMENTS
The authors thank to Dean of Faculty of Science and Technology for laboratory facilities, Phranakhon Rajabhat University. This research was supported by Higher Education Research Promotion (HERP). Under the office of the Higher Education Commission.
REFERENCES
- Adeoti, O., 2010. Water use impact of ethanol at a gasoline substitution ratio of 5% from cassava in Nigeria. Biomass Bioenergy, 34: 985-992.
CrossRefDirect Link - Aghighi, S., G.H.S. Bonjar, R. Rawashdeh, S. Batayneh and I. Saadoun, 2004. First report of antifungal spectra of activity of Iranian actinomycetes strains against Alternaria solani, Alternaria alternate, Fusarium solani, Phytophthora megasperma, Verticillium dahliae and Saccharomyces cerevisiae. Asian J. Plant Sci., 3: 463-471.
CrossRefDirect Link - Barros, J.A., E.V. Medeiros, K.A. Notaro, W.S. Moraes, J.M. Silva, T.C.E.S. Nascimento and K.A. Moreira, 2014. Different cover promote sandy soil suppressiveness to root rot disease of cassava caused by Fusarium solani. Afr. J. Microbiol. Res., 8: 967-973.
Direct Link - Bennett, M.J., A. Marchant, S.T. May and R. Swarup, 1998. Going the distance with auxin: Unravelling the molecular basis of auxin transport. Philos. Trans. R. Soc. London B: Biol. Sci., 353: 1511-1515.
CrossRefPubMedDirect Link - Bloemberg, G.V. and B.J.J. Lugtenberg, 2001. Molecular basis of plant growth promotion and biocontrol by rhizobacteria. Curr. Opin. Plant Biol., 4: 343-350.
CrossRefDirect Link - Boonlue, S., T. Aimi and T. Morinaga, 2003. Molecular characterization of a xylanase-producing thermophilic fungus isolated from Japanese soil. Curr. Microbiol., 47: 119-124.
CrossRefDirect Link - Buensanteai, N. and D. Athinuwat, 2012. The antagonistic activity of Trichoderma virens strain TvSUT10 against cassava stem rot in Thailand. Afr. J. Biotechnol., 11: 14996-15001.
Direct Link - Chaisinboon, O. and I. Chontanawat, 2011. Factors determining the competing use of Thailand's cassava for food and fuel. Energy Procedia, 9: 216-229.
CrossRefDirect Link - Cheng, J., S.H. Yang, S.A. Palaniyandi, J.S. Han, T. Yoon, T.J. Kim and J. Suh, 2010. Azalomycin F complex is an antifungal substance produced by Streptomyces malaysiensis MJM1968 isolated from agricultural soil. J. Korean Soc. Applied Biol. Chem., 53: 545-552.
CrossRefDirect Link - Cuesta, G., R. Garcia-de-la-Fuente, M. Abad and F. Fornes, 2012. Isolation and identification of actinomycetes from a compost-amended soil with potential as biocontrol agents. J. Environ. Manage., 95: S280-S284.
CrossRefPubMedDirect Link - Dobbelaere, S., A. Croonenborghs, A. Thys, A. Vande Broek and J. Vanderleyden, 1999. Phytostimulatory effect of Azospirillum brasilense wild type and mutant strains altered in IAA production on wheat. Plant Soil, 212: 153-162.
CrossRefDirect Link - Doumbou, C.L., M.K. Hamby-Salove, D.L. Crawford and C. Beaulieu, 2001. Actinomycetes, promising tools to control plant diseases and to promote plant growth. Phytoprotection, 82: 85-102.
CrossRefDirect Link - Elamvazhuthi, P. and M. Subramanian, 2013. Antagonistic activity of actinomycetes from jeypore paddy soils against selective phytopathogenic fungi. J. Mod. Biotechnol., 2: 66-72.
Direct Link - Felsenstein, J., 1985. Confidence limits on phylogenies: An approach using the bootstrap. Evolution, 39: 783-791.
CrossRefDirect Link - Gangwar, M., S. Rani and N. Sharma, 2012. Diversity of endophytic actinomyetes from wheat and its potential as plant growth promoting and biocontrol agents. J. Adv. Lab. Res. Biol., 3: 15-23.
Direct Link - Goudjal, Y., O. Toumatia, N. Sabaou, M. Barakate, F. Mathieu and A. Zitouni, 2013. Endophytic actinomycetes from spontaneous plants of Algerian Sahara: Indole-3-acetic acid production and tomato plants growth promoting activity. World J. Microbiol. Biotechnol., 29: 1821-1829.
CrossRefDirect Link - Hayakawa, M. and H. Nonomura, 1987. Humic acid-vitamin agar, a new medium for the selective isolation of soil actinomycetes. J. Ferment. Technol., 65: 501-509.
CrossRefDirect Link - Hirscha, A.M. and M. Valdesc, 2010. Micromonospora: An important microbe for biomedicine and potentially for biocontrol and biofuels. Soil Biol. Biochem., 42: 536-542.
CrossRefDirect Link - Khamna, S., A. Yokota and S. Lumyong, 2009. Actinomycetes isolated from medicinal plant rhizosphere soils: Diversity and screening of antifungal compounds, indole-3-acetic acid and siderophore production. World J. Microbiol. Biotechnol., 25: 649-655.
CrossRefDirect Link - Khucharoenphaisan, K., U. Puangpetch, K. Puttaraksa and K. Sinma, 2011. Grouping of actinomycetes isolated from termites using biochemical character. J. Biol. Sci., 11: 314-319.
CrossRefDirect Link - Khucharoenphaisan, K. and K. Sinma, 2011. Production and partial characterization of uric acid degrading enzyme from new source Saccharopolyspora sp. PNR11. Pak. J. Biol. Sci., 14: 226-231.
CrossRefDirect Link - Khucharoenphaisan, K., N. Sripairoj and K. Sinma, 2012. Isolation and identification of actinomycetes from termite's gut against human pathogen. Asian J. Anim. Vet. Adv., 7: 68-73.
CrossRefDirect Link - Khucharoenphaisan, K., K. Sinma and C. Lorrungruang, 2013. Efficiency of actinomycetes against phytopathogenic fungus of chilli anthracnose. J. Appl. Sci., 13: 472-478.
CrossRefDirect Link - Khucharoenphaisan, K., C. Suinongpai and K. Sinma, 2014. Isolation and screening of endophytic actinomycetes against phytopathogenic fungus of sugar cane red rot diseases. Proceedings of the Burapha University International Conference 2014: Global Warming and its Impacts, July 3-4, 2014, Pattaya, Thailand.
- Li, W., M. Csukai, A. Corran, P. Crowley, P.S. Solomon and R.P. Oliver, 2008. Malayamycin, a new streptomycete antifungal compound, specifically inhibits sporulation of Stagonospora nodorum (Berk) Castell and Germano, the cause of wheat glume blotch disease. Pest Manage. Sci., 64: 1294-1302.
CrossRefDirect Link - Nonoh, J.O., W. Lwande, D. Masiga, R. Herrmann and J.K. Presnail et al., 2010. Isolation and characterization of Streptomyces species with antifungal activity from selected national parks in Kenya. Afr. J. Microbiol. Res., 4: 856-864.
Direct Link - Prapagdee, B., C. Kuekulvong and S. Mongkolsuk, 2008. Antifungal potential of extracellular metabolites produced by Streptomyces hygroscopicus against phytopathogenic fungi. Int. J. Biol. Sci., 4: 330-337.
CrossRefPubMedDirect Link - Sharma, S.B., R.Z. Sayyed, M.H. Trivedi and T.A. Gobi, 2013. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. SpringerPlus, Vol. 2.
CrossRefDirect Link - Shirling, E.B. and D. Gottlieb, 1966. Methods for characterization of Streptomyces species. Int. J. Syst. Evol. Microbiol., 16: 313-340.
CrossRefDirect Link - Sinha, K., R. Hegde and A. Kush, 2014. Exploration on native actinomycetes strains and their potential against fungal plant pathogens. Int. J. Curr. Microbiol. Applied Sci., 3: 37-45.
Direct Link - Suwan, N., W. Boonying and S. Nalumpang, 2012. Antifungal activity of soil actinomycetes to control chilli anthracnose caused by Colletotrichum gloeosporioides. J. Agric. Technol., 8: 725-737.
Direct Link - Taechowisan, T., N. Chuaychot, S. Chanaphat, A. Wanbanjob and P. Tantiwachwutikul, 2009. Antagonistic Effects of Streptomyces sp. SRM1 on Colletotrichum musae. Biotechnology, 8: 86-92.
CrossRefDirect Link