
N.K. Njeru
School of Biological Sciences, University of Nairobi, Nairobi, Kenya
J.W. Muthomi
Department of Plant Science and Crop Protection, University of Nairobi, Nairobi, Kenya
C.K. Mutegi
International Institute of Tropical Agriculture, Nairobi, Kenya
J.M. Wagacha
School of Biological Sciences, University of Nairobi, Nairobi, Kenya
Journal of Plant Sciences
Year: 2016 | Volume: 11 | Issue: 1-3 | Page No.: 12-21







ABSTRACT
Fusarium head blight (FHB) is an economically important disease of wheat, which causes reduction in grain yield both quantitatively through reduced seed weight and qualitatively by contaminating grains with mycotoxins. The effect of cropping systems on accumulation of FHB inocula in crop residues and soil was assessed at hard dough stage of wheat during the 2013 cropping season in three agro-ecological zones in Narok County, Kenya. A semi structured questionnaire was used to obtain information on wheat production practices. Fusarium spp. were isolated from crop residues and top soil, while incidence and severity of FHB were assessed at mid-anthesis. Majority of the wheat farmers were small scale producers who rotated wheat with maize, grew the two crops in adjacent fields, grew wheat in consecutive years, left wheat residues as standing hay for livestock and practiced simple land preparation methods. Prevalence of FHB was 100%, while mean incidence and severity were 20.7 and 28.4%, respectively. The most frequently isolated Fusarium spp. were F. chlamydosporum and F. graminearum in crop residues and F. oxysporum and F. proliferatum in soil. The incidence of Fusarium spp. in soil and crop residues was highly correlated to FHB incidence but not to severity of the disease. Wheat production practices affect the survival of Fusarium spp. in soil and crop residues between cropping seasons. It is therefore, recommended that after harvesting, wheat straw and maize stover should be removed from the field or be incorporated into the soil through tillage to allow faster decomposition. Inclusion of maize as a rotation crop in wheat production should also be avoided.
PDF Abstract XML References Citation
Received: March 11, 2016;
Accepted: March 30, 2016;
Published: April 15, 2016
How to cite this article
N.K. Njeru, J.W. Muthomi, C.K. Mutegi and J.M. Wagacha, 2016. Effect of Cropping Systems on Accumulation of Fusarium Head
Blight of Wheat Inocula in Crop Residues and Soils. Journal of Plant Sciences, 11: 12-21.
DOI: 10.3923/jps.2016.12.21
URL: https://scialert.net/abstract/?doi=jps.2016.12.21
DOI: 10.3923/jps.2016.12.21
URL: https://scialert.net/abstract/?doi=jps.2016.12.21
INTRODUCTION
Wheat (Triticum aestivum L.) is the most widely cultivated cereal grain worldwide providing approximately 20% of global calories to humans (IRA., 2013). Trade in wheat and wheat products in the world are greater than for all other commercial crops combined (Curtis, 2002). In Kenya, wheat production output is lower than consumption with the deficit being met through imports. Narok County is one of Kenya’s leading wheat growing regions with over 100,000 ha under wheat production (Price Water House Coopers, 2005). With highland areas that lie 3000 m a.s.l. and rainfall that ranges between 1200-1800 mm per annum (Price Water House Coopers, 2005), Narok is highly suited for wheat farming.
Infection of wheat by various fungi including Fusarium, Puccinia, Alternaria and Epicoccum species causes reduction in grain yield and quality (McMullen et al., 2008; Matny et al., 2012). Infection of wheat ears by Fusarium spp. causes Fusarium head blight (FHB), which is one of the economically important diseases of wheat and other small grain cereals in humid and semi-humid tropics (Abedi-Tizaki and Sabbagh, 2012; Landschoot et al., 2013). Prevalence of FHB in the field varies from trace to virtually 100% and losses may vary correspondingly (Muthomi et al., 2012).
Fusarium head blight is caused by a complex of Fusarium spp. although, F. graminearum, F. poae, F. avenaceaum and F. culmorum are the most important worldwide (Parry et al., 1995). Fusarium graminearum predominates in hotter regions, while F. culmorum and F. poae are the predominant species in cooler regions (Parry et al., 1995; Holt, 2014). The disease causes reduction in grain yield both quantitatively through reduced seed weight and qualitatively by contaminating grains with mycotoxins. Infected wheat heads produce shriveled grains, which are shrunken and chalky white (Lipps, 2002). The extent of grain yield reduction depends on the Fusarium spp. involved (Henriksen and Elen, 2005; Wagacha et al., 2011).
Fusarium head blight is a result of the complex interaction between weather conditions and agricultural practices including crop rotation, tillage method, fungicide application and host resistance (Landschoot et al., 2013). Favorable environmental conditions such as frequent rainfall and high relative humidity enhance production of primary inocula on crop residues (Fernando et al., 2000; Landschoot et al., 2013). Extended periods of greater than 90% relative humidity and 15-30°C temperature enhance infection. Continued wet conditions after flowering may increase potential of the spores to be blown by wind or rain-splashed to nearby ears (Keller, 2011).
Rotation or mixed cropping of wheat with another cereal crop promotes accumulation of FHB inocula with the cereals acting as alternative hosts for Fusarium spp. (Silvestro et al., 2013). The amount of crop residues at planting is determined by the previous crop and the type of tillage done (Dill-Macky and Jones, 2000). Tillage buries residues below the soil surface and allows for their decomposition thereby reducing the amount of inocula available for spread (Kleczewski, 2014). This implies that minimal or no tillage contributes to FHB epidemics because it leaves crop residues on the soil surface. The level of resistance of a variety also affects the incidence of FHB of wheat (Landschoot et al., 2013).
All Fusarium spp. that infect cereals survive as saprophytes on crop residues and contribute inocula for infection of wheat plants in the field, in which the residue is present and more distant fields (Parry et al., 1995; Keller, 2011). Infected crop residues left in the field also play a role in survival and spread of Fusarium spp. by increasing the population of the pathogens in soils (Keller, 2011; Silvestro et al., 2013). This study evaluated the effect of cropping systems on accumulation of Fusarium head blight of wheat inocula in soil and crop debris in Narok County, Kenya.
MATERIALS AND METHODS
Field survey and sampling: A field survey covering 51 wheat farms were carried out in Narok County between June and October, 2013. The survey was carried out at mid-anthesis (GS 65-69) in three Agro-Ecological Zones (AEZ), 15 wheat farms in Lower Highland 3 (LH3), 18 in Lower Highland 2 (LH2) and 18 in Upper Highland 3 (UH3). This was done by selecting the 5th wheat growing farm in a transect. A semi structured questionnaire was used to obtain information on wheat production practices. Incidence of FHB was determined as the number of blighted heads over the total number of heads within 5, 1 m2 randomly selected quadrants in each farm. Severity of FHB was measured on a scale of 0-9 as the average proportion of the bleached spikelets (1 = no symptoms, 2 = <5%, 3 = 5-15%, 4 = 16-25%, 5 = 25-44%, 6 = 46-65%, 7 = 66-85%, 8 = 86-95%, 9 = 96-100% (Miedaner et al., 1996). Approximately 200 g composite sample of top soil and 250 g of crop residues were sampled from each farm. Soil was sampled from the top 5 cm in a zigzag pattern in each farm and then mixed to make a composite sample from which 1 g sub-sample was taken for fungal isolation. The soil samples were air dried and stored at room temperature (23±2°C).
Isolation and identification of Fusarium spp. from crop residues and soil: Fungal pathogens were isolated on low strength Potato Dextrose Agar (PDA, 17 g PDA in 1000 mL of distilled water) amended with minerals and antibiotics (Muthomi et al., 2002). Fifty milligrams of penicillin, tetracycline, streptomycin and pentachloronitrobenzene (PCNB) each were added after the media was cooled to approximately 45°C. The crop residues were cut into small pieces (≈0.5 cm long) and surface sterilized in 1.3% sodium hypochlorite for 3 min and rinsed 3 times in sterile distilled water. Five pieces of crop residues were plated in each petri plate and replicated 3 times. The plates were incubated for 5 days at 25°C under 12 h day light and 12 h darkness cycles. The number of residues per plate infected with Fusarium spp. and other fungal pathogens were counted.
Soil samples were passed through 2 mm mesh sieve to remove crop residues and 1 g was suspended in 9 mL sterile distilled water. The suspension was serially diluted to 10–3 and 200 μL aliquot of the 10–3 dilution was plated on low strength PDA. The plates were incubated for 5-10 days after which colonies of Fusarium spp. and other fungi were counted and the number of Colony Forming Units per gram (CFU g–1) of soil calculated as follows:
![]()
The relative isolation frequency of each Fusarium spp. and other fungal pathogens were calculated as follows (Gonzalez et al., 1999):
![]()
Fusarium colonies were sub-cultured on PDA and Synthetic Nutrient Agar (SNA; Nirenberg, 1981): (KH2PO4 1.0 g, KNO3 1.0 g, MgSO4 0.5 g, KCl 0.5 g, glucose 0.2 g and agar 20 g). Fusarium cultures on SNA were incubated for 10-14 days under near UV-light to facilitate sporulation, while those on PDA were incubated at 25°C for 5-10 days. Cultures on PDA were used for identification of Fusarium spp. based on cultural characterics, while cultures on SNA were used for microscopic identification. Features for microscopic identification were morphology of microconidia and macroconidia, type of conidiophores and chlamydospores. Fusarium spp. were identified to species level using manuals by Nelson et al. (1983) and Leslie and Summerell (2006). Other fungal colonies were sub-cultured on PDA, incubated at 25°C for 10 days and identified to genus level based on cultural and morphological characteristics using manual by Barnett and Hunter (1998).
Data analysis: Data were subjected to analysis of variance (ANOVA) using the PROC ANOVA procedure of GENSTAT version 15 and the mean differences compared using Fisher’s protected LSD at 5% probability level. The same software was used to carry out correlation analysis for incidence of Fusarium spp. in crop residues and soil and FHB incidence and severity. Data that were not normally distributed were transformed using arcsine before analysis.
RESULTS
Farming practices in Narok County, Kenya: Small scale farms were classified as farms which were less than 8 ha, medium scale farms ranged from 8-20 ha while farms greater than 20 ha were considered large scale (Fig. 1a).
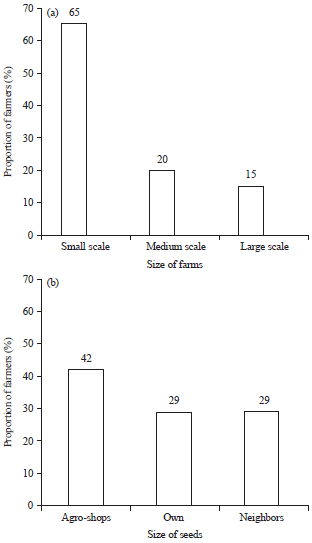 | |
| Fig. 1(a-b): | Percentage of farmers growing wheat in different (a) Farm sizes and (b) Sources of wheat planting seeds in Narok County |
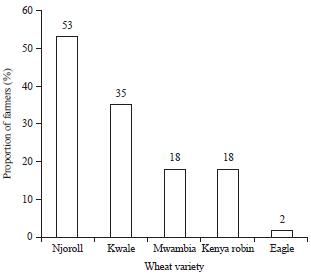 | |
| Fig. 2: | Percentage of farmers growing different wheat varieties during the 2013 cropping season in Narok County |
Majority (58%) of the wheat farmers did not have a specific place, where they sourced planting seeds. The farmers mainly used their own seeds saved from the previous crop and from neighbors and then mixed the two for planting. However, 42% of the farmers bought planting seeds from agro-shops (Fig. 1b). Five varieties of wheat-Kwale, Njoro II, Mwamba, Kenya Robin II and eagle were grown (Fig. 2). Variety Kwale was grown by 53% of farmers across all the agro-ecological zones, while the least popular variety was eagle, which was grown by only 2% of the farmers.
Majority of the farmers practiced simple land preparation practices-ploughing and harrowing, while the rest practiced chisel ploughing and/or harrowing (Fig. 3a), however, all the farmers used tractor plough. Some farmers (43%) applied the herbicide Round-up® after ploughing. Majority (65%) of the farmers applied di-ammonium phosphate (DAP) fertilizer at planting and a small proportion (11%) of farmers who did not apply DAP used farmyard manure to plant other crops like maize and potatoes (Fig. 3b).
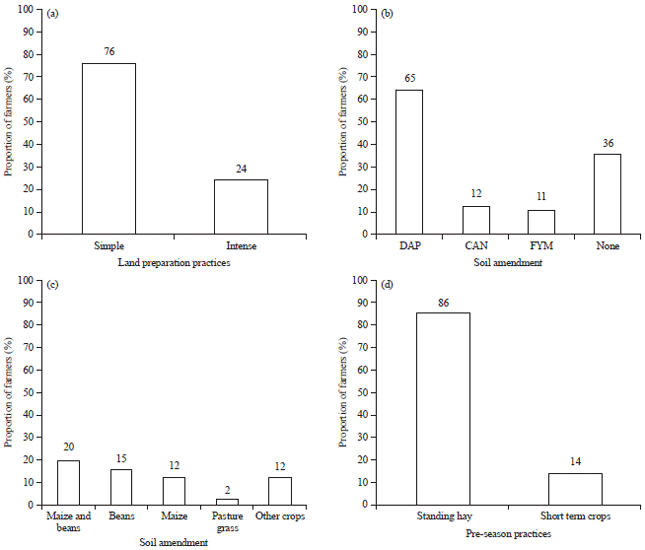 | |
| Fig. 3(a-d): | Proportion of farmers practicing various methods of (a) Land preparation, (b) Applying various soil amendments, (c) Other food crops commonly grown in fields, where wheat was cultivated and (d) Handling of wheat residues after harvesting and pre-season practices, DAP: Di-ammonium phosphate, CAN: Calcium ammonium nitrate, FYM: Farm yard manure and other crops-peas, potatoes |
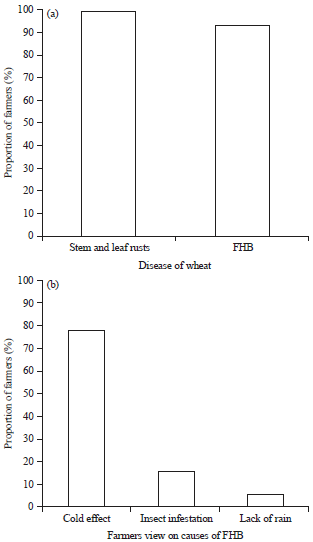 | |
| Fig. 4(a-b): | Prevalence of (a) Wheat diseases in Narok County and (b) Farmers’ views on causes of FHB |
Additionally, 12% of wheat farmers who used DAP at planting also applied Calcium Ammonium Nitrate (CAN) foliar fertilizer on wheat before heading stage.
Wheat was mainly grown as a mono crop but 2% of the farmers intercropped wheat with the pasture grass, Boma rhodes. Other crops commonly grown between wheat cropping seasons or in rotation were maize, beans, peas and potatoes (Fig. 3c). The proportion of wheat farmers that practiced crop rotation was 57% with 98% of these farmers leaving wheat straw and maize stover as standing fodder for livestock and a few (2%) burning the crop residues before land preparation. More than 50% of the farmers grew wheat and maize in adjacent fields. After harvesting, wheat straw was left as standing hay for livestock until the next wheat cropping season (Fig. 3d) and livestock grazed freely mainly in the farms where wheat or maize were grown.
| Table 1: | Prevalence, incidence and severity of FHB of wheat in different agro-ecological zones in Narok County, Kenya |
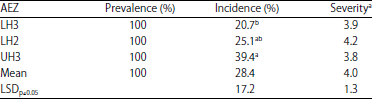 | |
Severity score based on scale by Miedaner et al. (1996). 1: No symptoms, 2: <5%, 3: 5-15%, 4: 16-25%, 5: 25-44%, 6: 46-65%, 7: 66-85%, 8: 86-95%, 9: 96-100%. Incidence means followed by different letters are significantly different (p≤0.05) | |
Majority (86%) of the farmers grew one wheat crop annually and the rest (14%) planted short term crops between wheat cropping seasons.
Major diseases of wheat in Narok county: The most common diseases affecting wheat were stem rust (Puccinia graminis), leaf rust (Puccinia triticina) and Fusarium head blight (Fig. 4a). Stem rust and leaf rust were more prevalent, with the farmers having greater knowledge on stem rust than leaf rust. Although most farmers (94%) could recognize FHB, majority (63%) of the farmers thought that FHB was frost damage or "cold effect", 37% considered it to be another symptom of rust, 16% considered it to be insect infestation, while 6% thought it was early maturity due to lack of enough rains (Fig. 4b). The farmers used Folicur® to manage both rust and FHB while some used insecticides such as Bulldock®.
Prevalence, incidence and severity of FHB of wheat in Narok County: Fusarium head blight was observed in all the farms sampled but the disease incidence and severity varied among farms and agro-ecological zones (Table 1). Incidence of FHB varied between 1 and 100%, while severity varied from 1-95%. The highest FHB incidence and severity were observed in AEZ UH3, while the lowest incidence and severity were observed in AEZ LH3. There was no significant (p≥0.05) difference in FHB severity among the three AEZ, but incidence significantly (p≤0.05) varied among the AEZ.
Incidence of Fusarium spp. in crop residues and soil sampled from wheat fields
Incidence of Fusarium spp. in crop residues: The fungi isolated in high frequencies from crop residues were Fusarium spp., Pythium spp. and Trichoderma spp. while Alternaria spp., Colletotrichum spp., Curvularia spp. and Cladosporium spp. were isolated in low frequencies (<0.1%) (Table 2). Fusarium spp. and Pythium spp. were isolated in the highest frequency in the three agro-ecological zones with a mean incidence of 54.2 and 23.4%, respectively.
| Table 2: | Isolation frequency (%) of fungal pathogens from crop residues sampled from wheat fields in different agro-ecological zones of Narok County |
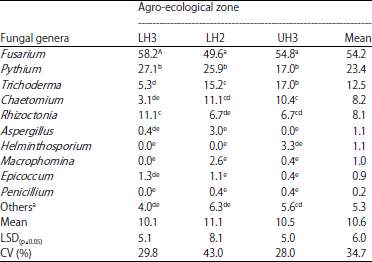 | |
Means followed by different letters within columns are significantly different (p≤0.05), LH3: Lower highland 3, LH2: Lower highland 2: UH3: Upper highland 3, aFungi isolated in low frequency and unidentified fungal species | |
| Table 3: | Incidence (%) of Fusarium spp. in crop residues sampled from wheat fields in different agro-ecological zones in Narok County |
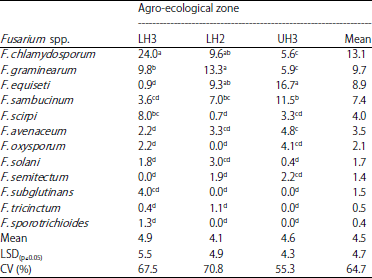 | |
Means followed by different letters within columns are significantly different (p≤0.05), LH3: Lower highland 3, LH2: Lower highland 2, UH3: Upper highland 3 | |
The incidence of fungal pathogens in crop residues varied significantly (p≤0.05) among the three agro-ecological zones; and 17% of the crop residue samples had multiple fungal infections. The incidence of Fusarium spp. in crop residues varied significantly (p≤0.05) between LH3 and UH3. Twelve Fusarium spp. were isolated from crop residues with F. chlamydosporum, F. graminearum, F. equiseti and F. sambucinum being the major species (Table 3). Fusarium chlamydosporum was the most prevalent in LH3, F. graminearum in LH2 and F. equiseti in UH3.
Incidence of Fusarium spp. in soil: The incidence of fungal pathogens in soil significantly (p≤0.05) varied among the three AEZ (Table 4). Penicillium spp. were the most prevalent in agro-ecological zone LH3, Aspergillus spp. in LH2 and Fusarium spp. in UH3.
| Table 4: | Population (CFU/g soil×1000) of fungal pathogens in soil sampled from wheat fields in different agro-ecological zones of Narok County |
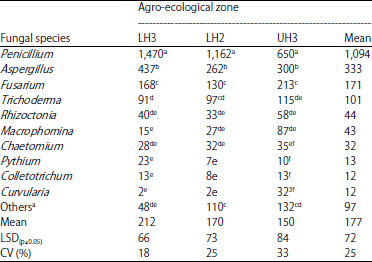 | |
Means followed by different letters within columns are significantly different (p≤0.05), LH3: Lower highland 3, LH2: Lower highland 2, UH3: Upper highland 3, aUnidentified fungal species | |
| Table 5: | Population (CFU/g soil) of Fusarium spp. in soil sampled from wheat fields in different agro-ecological zones in Narok County |
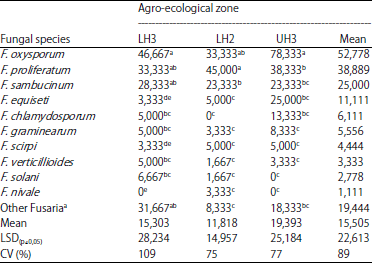 | |
Means followed by different letters within columns are significantly different (p≤0.05), LH3: Lower highland 3, LH2: Lower highland 2, UH3: Upper highland 3, aUnidentified Fusarium species | |
The population of Fusarium spp. in the soils was up to 168×103 CFU g–1 soil. The major Fusarium spp. isolated from soil were F. oxysporum, F. proliferatum and F. sambucinum (Table 5) with Fusarium oxysporum being the most prevalent (up to 78,000 CFU g–1 soil) in LH3 and UH3, while F. proliferatum was the most prevalent in LH2.
Correlation among the incidence of Fusarium spp. in crop residues and soil and FHB incidence and severity: The incidence of Fusarium spp. in crop residues and soil was highly correlated to the incidence of FHB at hard dough stage (Table 6). However, there was no significant correlation between the incidence of Fusarium spp. in crop residues and FHB severity.
| Table 6: | Correlation among incidence of Fusarium spp. in crop residues and soil and FHB incidence and severity |
 | |
ns: Not significant, *Significant, **Highly significant, aIncidence of Fusarium spp. in crop residues, bIncidence of Fusarium spp. in soil | |
DISCUSSION
Most of the wheat farmers in Narok County were small scale producers who used farm saved seeds and rarely used pesticides to control pests and diseases of wheat. Some of the small scale wheat farmers lacked knowledge on key disease management practices like crop rotation, land preparation methods, residue management and knowledge of common wheat diseases and their management. Planting seeds infected with FHB pathogens could result in low germination and low seedling vigor, predisposing the wheat seedlings to the disease (Kleczewski, 2013). Therefore, infected planting seeds could have contributed to the high prevalence of FHB in Narok County, where most farmers planted seeds saved from previous seasons.
The most widely grown wheat variety, Kwale is high yielding and moderately susceptible to FHB (Muthomi et al., 2007). Wheat varieties differ in their susceptibility to FHB and therefore, the variety grown could determine the levels of FHB and mycotoxin contamination (Dill-Macky and Jones, 2000; Nopsa, 2010). Previous studies have demonstrated that all wheat varieties grown in Kenya are susceptible to FHB (Muthomi et al., 2002; Ndungu, 2006; Muthomi et al., 2007). This implies that a FHB epidemic could pose a serious threat to wheat production in the country.
Most farmers left wheat residues as standing hay for livestock consumption until the next wheat cropping season. Livestock spread infected wheat residues across the fields as they grazed freely. All Fusarium spp. that cause FHB are capable of surviving as saprophytes on crop residues (Parry et al., 1995). Wheat and maize residues act as sources of primary inocula for FHB and also as a pathway for introduction of mycotoxins to the animal feed chain (FSA., 2007; Wagacha et al., 2010; Keller, 2011).
The findings of this study concur with the report by Dill-Macky and Jones (2000) observed a positive relationship between previous crop residues, tillage and the incidence of FHB. However, tillage has little effect on FHB inocula following crop rotation or mixed cropping with legumes (Dill-Macky and Jones, 2000) and other short term crops. The high proportion of farmers who rotated wheat with maize and those who did not practice crop rotation could have contributed to the high prevalence of FHB.
Majority (>50%) of the farmers grew wheat and maize in adjacent fields, 32% rotated wheat with maize and 43% grew wheat in consecutive years. Maize and wheat in adjacent fields could provide alternative hosts for Fusarium spp. Growing wheat and maize in adjacent farms also contributes to spread of FHB inocula and secondary infection (Wagacha et al., 2010). Del Ponte et al. (2003) observed that under favorable conditions, severe FHB symptoms can develop in wheat fields, where there is no significant inoculum coming from within the field. However, only Fusarium spores produced at wheat flowering stage become primary inocula (Wilcoxson et al., 1992; Inch et al., 2005).
Maize and wheat are the most susceptible cereal host crops to Fusarium infection (FSA., 2007). Rotation of wheat with maize and intensive cereal production increases the reservoir of FHB inocula in the soil because Fusarium spp., inocula overwinter in the partly decomposed cereal residues (Dill-Macky and Jones, 2000; Pereyra and Dill-Macky, 2004). Guo et al. (2010) and Landschoot et al. (2013) showed that conventional tillage made significant reduction in inocula levels of Fusarium spp. However, repeated wheat-maize rotation results in higher incidence of FHB when coupled with favorable weather conditions than tillage methods and wheat variety. This could explain the high prevalence of FHB observed in Narok where 32% of farmers rotated wheat with maize and 43% grew wheat in consecutive years.
Most farmers could not recognize FHB and thought it was caused by lack of rains, insect infestation, while others thought it was frost damage. Lack of recognition of the disease by majority of the farmers could be attributed to its symptom manifestation particularly the bleaching, which could be caused by other factors such as drought (Snijders and Perkowski, 1990; Matny et al., 2012). Lack of proper diagnosis of the disease implies that the farmers did not apply any specific measures for FHB management and control. Stem rust and leaf rust were also common diseases of wheat. Rusts are more serious if plants are attacked at milk stage or earlier (CABI., 2005). Stem rust, caused by Puccinia graminis badly shrivels kernels, many of them being so light and chaffy that they are blown out with chaff during threshing (CABI., 2005). However, stem rust does not overwinter and therefore does not usually cause significant yield losses. A recent study showed significant losses by rusts are rare in the world, although the disease can occur at significant levels in particular fields or throughout a particular growing season (Marsalis and Goldberg, 2013).
Most (54%) of the sampled crop residues were infected with Fusarium spp. Crop residues act as substrate for Fusarium spp., which survive as mycelia, conidia and ascospores (McMullen et al., 1997; Dill-Macky and Jones, 2000). The survival structures provide the primary inocula during the subsequent cropping season. Extended periods of greater than 90% relative humidity coupled with temperature between 15 and 30°C facilitate infection of wheat ears by Fusarium spp. (De Wolf et al., 2003). Cross infection of wheat ears occurs by wind blowing or water splashed when conducive environmental conditions continue after flowering (Cowger and Sutton, 2005; Maldonado-Ramirez et al., 2005).
Multiple Fusarium spp. were isolated from crop residues with F. chlamydosporum, F. graminearum, F. equiseti and F. sambucinum being the most common. These findings concur with the findings by Muthomi et al. (2007) and Wagacha et al. (2010). The higher incidence of F. chlamydosporum than F. graminerum the most virulent Fusarium species could be attributed to the fact that the latter species does not produce chlamydosopores. Fusarium chlamydosporum and F. equiseti produce chlamydospores that may over winter in crop residues, while F. graminearium survive in crop residues as ascospores, conidia or mycelia (Dill-Macky and Jones, 2000; Guo et al., 2010). The high incidence of F. graminearum and F. equiseti imply their causative nature of the observed FHB in Narok County. Fusarium graminearum causes blighting of infected wheat ears and is highly virulent and the main cause of FHB of wheat worldwide (Brennan et al., 2005; Leslie and Summerell, 2006).
Fusarium spp. were also isolated from most of the soil samples but in lower incidence than from crop residues. Fusarium oxysporum and F. proliferatum were the predominant species in soil. These results concur with the findings by Latiffah et al. (2007), Lukall (2012) and Silvestro et al. (2013) and contrast the findings by Muthomi et al. (2012) and Jeschke et al. (1987). Fusarium proliferatum has a wide range of hosts and produces high levels of fumonisins in maize (Leslie and Summerell, 2006) a crop which in this study was commonly grown as a rotation crop with wheat by the farmers. Fusarium oxysporum is a cosmopolitan soil saprophyte (Leslie and Summerell, 2006) that causes many plant diseases, including vascular wilts, damping off, crown rots and root rots (Summerell and Rugg, 1992).
Fusarium spp., persist as mycelia, chlamydospores and conidia in soil (McMullen and Stack, 1983; Keller, 2011). This implies that soil is a potential source of FHB inocula as well as inocula for other crop diseases. There were significant differences in the incidence of Fusarium spp. in soil among the three agro-ecological zones. The differences could be attributed to differences in soil types (Larkin et al., 1993) and cropping systems. The presence of Fusarium spp. in agricultural soils could impact on crop health and consequently on the amount and quality of human food and animal feeds (Keller, 2011).
CONCLUSION
This study showed that wheat production practices by farmers in Narok County contribute to build-up of FHB primary inocula and consequently to high prevalence of FHB. The incidence of Fusarium spp. in crop residues and soil were highly correlated to the incidence of FHB but not to severity of FHB. The high incidence and diversity of Fusarium spp. in crop residues and soil implies that crop residues and soil play an important role in the survival of FHB inocula between cropping seasons and consequently to FHB. Inclusion of maize as a rotation crop in wheat production and leaving crop residues on the soil surface after harvesting should be avoided.
ACKNOWLEDGMENTS
The Regional Universities Forum for Capacity Building in Agriculture (RUFORUM) is gratefully acknowledged for funding this study (RU/2012/GRG-69). We also acknowledge Extension Officers, Ministry of Agriculture in Narok County for logistical support during sample collection.
REFERENCES
- Brennan, J.M., D. Egan, B.M. Cooke and F.M. Doohan, 2005. Effect of temperature on head blight of wheat caused by Fusarium culmorum and F. graminearum. Plant Pathol., 54: 156-160.
CrossRefDirect Link - De Wolf, E.D., L.V. Madden and P.E. Lipps, 2003. Risk assessment models for wheat Fusarium head blight epidemics based on within-season weather data. Phytopathology, 93: 428-435.
CrossRefDirect Link - Dill-Macky, R. and R.K. Jones, 2000. The effect of previous crop residues and tillage on Fusarium head blight of wheat. Plant Dis., 84: 71-76.
CrossRefDirect Link - Fernando, W.G.D., J.D. Miller, W.L. Seaman, K. Seifert and T.C. Paulitz, 2000. Daily and seasonal dynamics of airborne spores of Fusarium graminearum and other Fusarium species sampled over wheat plots. Can. J. Bot., 78: 497-505.
CrossRefDirect Link - Gonzalez, H.H.L., E.J. Martinez, A. Pacin and S.L. Resnik, 1999. Relationship between Fusarium graminearum and Alternaria alternata contamination and deoxynivalenol occurrence on Argentinian durum wheat. Mycopathologia, 144: 97-102.
Direct Link - Guo, X.W., W.G.D. Fernando, P. Bullock and H. Sapirstein, 2010. Quantifying cropping practices in relation to inoculum levels of Fusarium graminearum on crop stubble. Plant Pathol., 59: 1107-1113.
CrossRefDirect Link - Henriksen, B. and O. Elen, 2005. Natural Fusarium grain infection level in wheat, barley and oat after early application of fungicides and herbicides. J. Phytopathol., 135: 214-220.
CrossRefDirect Link - Inch, S., W.G.D. Fernando and J. Gilbert, 2005. Seasonal and daily variation in the airborne concentration of Gibberella zeae (Schw.) Petch spores in Manitoba. Canadian J. Plant Pathol., 27: 357-363.
CrossRefDirect Link - Jeschke, N., P.E. Nelson and W.F.O. Marasas, 1987. Toxicity to ducklings of Fusarium moniliforme isolated from corn intended for use in poultry feed. Poult. Sci., 66: 1619-1623.
CrossRefDirect Link - Landschoot, S., K. Audenaert, W. Waegeman, B. de Baets and G. Haesaert, 2013. Influence of maize-wheat rotation systems on Fusarium head blight infection and deoxynivalenol content in wheat under low versus high disease pressure. Crop Protect., 52: 14-21.
CrossRefDirect Link - Larkin, R.P., D.I. Hopkins and F.N. Martin, 1993. Ecology of Fusarium oxysporum F. sp. niveum in soils suppressive and conducive to Fusarium wilt of watermelon. Phytopathology, 83: 1105-1116.
CrossRefDirect Link - Latiffah, Z., M.M. Zariman and S. Baharuddin, 2007. Diversity of Fusarium species in cultivated soils in Penang. Malaysian J. Microbiol., 3: 27-30.
Direct Link - Maldonado-Ramirez, S.L., D.G. Schmale III, E.J. Shields and G.C. Bergstrom, 2005. The relative abundance of viable spores of Gibberella zeae in the planetary boundary layer suggests the role of long-distance transport in regional epidemics of Fusarium head blight. Agric. For. Meteorol., 132: 20-27.
CrossRefDirect Link - McMullen, M.P., R. Jones and D. Gallenberg, 1997. Scab of wheat and barley: A re-emerging disease of devastating impact. Plant Dis., 81: 1340-1348.
CrossRefDirect Link - McMullen, M.P. and R.W. Stack, 1983. Fusarium species associated with grassland soils. Can. J. Bot., 61: 2530-2538.
CrossRefDirect Link - Miedaner, T., G. Gang and H.H. Geiger, 1996. Quantitative-genetic basis of aggressiveness of 42 isolates of Fusarium culmorum for winter rye head blight. Plant Dis., 80: 500-504.
Direct Link - Muthomi, J.W., S.L. Musyimi, J.M. Wagacha and R.D. Narla, 2012. Occurrence of Fusarium species and associated T2-toxin in Kenyan wheat. Agric. Sci., 3: 24-34.
CrossRefDirect Link - Muthomi, J.W., E.C. Oerke, H.W. Dehne and E.W. Mutitu, 2002. Susceptibility of Kenyan wheat varieties to head blight, fungal invasion and deoxynivalenol accumulation inoculated with Fusarium graminearum. J. Phytopathol., 150: 30-36.
CrossRefDirect Link - Nelson, P.E., T.A. Toussoun and W.F.O. Marasas, 1983. Fusarium Species: An Illustrated Manual for Identification. 1st Edn., Pennsylvania State University Press, University Park, University Park, PA., USA., ISBN-13: 978-0271003498, Pages: 226.
Direct Link - Nirenberg, H.I., 1981. A simplified method for identifying Fusarium spp. occurring on wheat. Can. J. Bot., 59: 1599-1609.
CrossRefDirect Link - Parry, D.W., P. Jenkinson and L. McLeod, 1995. Fusarium ear blight (Scab) in small grain cereals-A review. Plant Pathol., 44: 207-238.
CrossRefDirect Link - Pereyra, S.A., R. Dill-Macky and A.L. Sims, 2004. Survival and inoculum production of Gibberella zeae in wheat residue. Plant Dis., 88: 724-730.
CrossRefDirect Link - Silvestro, L.B., S.A. Stenglein, H. Forjan, A.M. Arambarri, L. Manso and M.V. Moreno, 2013. Occurrence and distribution of soil Fusarium species under wheat crop in zero tillage. Spanish J. Agric. Res., 11: 72-79.
Direct Link - Snijders, C.H.A. and J. Perkowski, 1990. Effects of head blight caused by Fusarium culmorum on toxin content and weight of wheat kernels. Phytopathology, 80: 566-570.
Direct Link - Abedi-Tizaki, M. and S.K. Sabbagh, 2012. Morphological and molecular identification of Fusarium head blight isolates from wheat in North of Iran. Aust. J. Crop Sci., 6: 1356-1361.
Direct Link - Wagacha, J.M., U. Steiner, H.W. Dehne, S. Zuehlke, M. Spiteller, J. Muthomi and E.C. Oerke, 2010. Diversity in mycotoxins and fungal species infecting wheat in Nakuru District, Kenya. J. Phytopathol., 158: 527-535.
CrossRefDirect Link - Wilcoxson, R.D., R.H. Busch and E.A. Ozmon, 1992. Fusarium head blight resistance in spring wheat cultivars. Plant Dis., 76: 658-661.
Direct Link - Summerell, B.A. and C.A. Rugg, 1992. Vascular wilt of Helichrysum species caused by Fusarium oxysporum. Aust. Plant Pathol., 21: 18-19.
CrossRefDirect Link