
Chirom Oceania
Department of Biotechnology, Manipur University, Canchipur, Imphal, 795003, India
Thingujam Doni
Department of Biotechnology, Manipur University, Canchipur, Imphal, 795003, India
Leimapokpam Tikendra
Department of Biotechnology, Manipur University, Canchipur, Imphal, 795003, India
Potshangbam Nongdam
Department of Biotechnology, Manipur University, Canchipur, Imphal, 795003, India
Journal of Plant Sciences
Year: 2015 | Volume: 10 | Issue: 1 | Page No.: 15-24







ABSTRACT
Tomato is a highly popular crop for its rich nutritional values and constitutes an integral part of daily diets for substantial portion of human population of the world. Because of limitations in conventional breeding methods and increasing demand of this important crop for fast growing population, large scale propagation of tomato through plant tissue culture techniques becomes highly significant. Efficient in vitro generation protocols for tomato have been established successfully by employing seed and tissue culture derived shoot-tip explants. 6-benzyl amino purine (BAP) was highly effective in inducing seed germination and rapid leaf multiplication though presence of 0.4% activated charcocal (AC) had negative influence on culture development. The used of in vitro derived shoot-tips as explants removed the deleterious effect of AC resulting in the production of highest number of leaves in MS+3 mg L-1 BAP+0.5 mg L-1 NAA+AC. The Murashige and Skoog (MS) medium augmented with as 1-naphthalene acetic acid (NAA) was not favourable for seed germination but germination rate and root multiplication responses accelerated with the incorporation of AC in medium. Maximum number of roots differentiated from the germinating seeds in MS+3 mg L-1 NAA+1 mg L-1 BAP+AC. The MS medium supplemented with 1.5 mg L-1 NAA generated highest root formation when shoot-tip propagules were used for culture initiation. Preliminary investigation of synthetic seed development of tomato revealed the gel matrix produced from the complexation between 2% sodium alginate and 100 mM CaCl2 for 20-40 min as the most favourable for efficient encapsulation of in vitro derived shoot-tip propagules. The in vitro protocols developed from the present investigation can be utilized for propagation of tomato in larger scale for commercial purposes as well as local domestic consumption.
PDF Abstract XML References Citation
Received: March 26, 2015;
Accepted: May 13, 2015;
Published: May 27, 2015
How to cite this article
Chirom Oceania, Thingujam Doni, Leimapokpam Tikendra and Potshangbam Nongdam, 2015. Establishment of Efficient in vitro Culture and Plantlet Generation of Tomato (Lycopersicon esculentum Mill.) and Development of Synthetic Seeds. Journal of Plant Sciences, 10: 15-24.
DOI: 10.3923/jps.2015.15.24
URL: https://scialert.net/abstract/?doi=jps.2015.15.24
DOI: 10.3923/jps.2015.15.24
URL: https://scialert.net/abstract/?doi=jps.2015.15.24
INTRODUCTION
Tomato (Lycopersicon esculentum Mill.) belonging to the family Solanaceae is considered as the second most popular vegetable crop after potato in the world (Bhatia et al., 2005). The worldwide tomato production is 115 Mt from an area of 4.4 Mha but India produces 9 Mt from tomato cultivated area of 5.2 lakh hectars (Lumpkin, 2005; Himabindu et al., 2012). The tomato fruits have high nutritional values and are used extensively as one of the important ingredients in the preparation of wide range of vegetarian and non-vegetarian dishes. It is considered as an important commercial and dietary vegetable crop conferring health benefits because of rich contents of vitamins, minerals and antioxidant flavonoids like lycopene and zetaxanthin (Hossain et al., 2011; Singh et al., 2014). The conventional seed propagation method of new cultivar production and yield enhancement is less successful because of longer time requirement for cultivar development and several existing biotic and abiotic stress in normal field conditions. In vitro culture techniques can be employed to shorten time period for cultivar development and to enhance breeding efficiency. Improvement of tomato variety in terms of longer self-life, enhanced nutrient content and resistance to biotic and abiotic stress can be achieved through genetic manipulation using plant genetic engineering approach. An efficient in vitro regeneration protocol for tomato plants is a prerequisite for high production of transformed plants. Moreover, there is an immediate necessity to improve tomato production in order to meet the increasing demand of fast expanding human population and plant tissue culture techniques may offer an excellent opportunity of fulfilling the requirement by propagating tomato plants in large scale. There are reports of successful tomato regeneration in vitro using different explants (Bhatia et al., 2004; Mamidala and Nanna, 2011; Lima et al., 2004; Chandra et al., 2013). The synthetic seed technology which involves the encapsulation of small sized plant propagules with protective nutrient gel provides another promising option for mass propagation because of higher scale up capacity, possibility of automation of whole product process and direct delivery to field. Though synthetic seeds have been developed for potato (Patel et al., 2000), apple (Piccioni, 1997), Catharanthus roseus (Maqsood et al., 2012), banana (Hassanein et al., 2011), eggplant (Huda et al., 2007), black pepper (Nair and Gupta, 2007) and orchid (Mohanraj et al., 2009; Nagananda et al., 2011), reports on synthetic seed production in tomato are very scarce. The present study was conducted with an aim of establishing efficient and reproducible in vitro regeneration protocol for large scale propagation of tomato through seeds and shoot-tip culture. Preliminary investigation was also done to develop better quality synthetic seeds for tomato by encapsulating shoot-tip propagules with varied concentration of sodium alginate and calcium chloride at different time interval.
MATERIALS AND METHODS
Plant materials and explant surface sterilization: The present study was conducted at Department of Biotechnology, Manipur University, Manipur, India from February, 2014 to July, 2014. The fruits of commercially available tomato (Lycopersicon esculentum Mill.) were collected for the present investigation. The fruits were first washed thoroughly in running tape water with 20% teepol as wetting agent for 30-45 min followed by treatment with 70% ethanol for 1 min. The treated fruits were washed 3-4 times with double sterile distilled water before being subjected to 0.2% mercuric chloride treatment for 4-5 min. The fruits were finally rinsed 5-6 times with sterile distilled water to remove any mercuric chloride present on the surface. The fruits were finally bolt dried on to sterilized filter paper after which longitudinally cuts were made on to the fruits using sterile scalpel to remove the seeds for in vitro culture initiation.
Preparation of media, growth hormone combinations and incubation: The Murashige and Skoog (1962) medium was employed for in vitro culture and plantlet generation of tomato plants. Separate stock solutions were prepared for macro and micro-nutrients, vitamins, hormones and iron salts in a concentrated solution and stored at 25-30°C until use. Culture medium was prepared by adding the macro and micro nutrients, vitamins and other salts from the stock solutions one after another into a conical flask of desired size followed by volume make up with sterile distilled water as per the requirement. The pH was adjusted at 5.8 after adding 2% sucrose and the medium was boiled with 0.9% agar as gelling agent. Activated charcoal (0.4%) was incorporated into the medium which required sufficient swirling of culture vessels to completely dissolve the activated charcoal (AC). Definite quantities of medium were dispensed to culture tubes after growth regulators was added and culture vessels were tightly closed with appropriately sized cotton plugs. The medium was autoclaved finally at 121°C for 15 min and placed in an appropriate position either vertical or slanting to allow sufficient medium gelling. The medium was supplemented with indole-3-butyric acid (IBA), 1-naphthalene acetic acid (NAA) and 6-benzyl amino purine (BAP) in varied combinations and doses to study their influence on seed germination, shooting and rooting multiplication response and plantlet regeneration in vitro. The seeds obtained from surface sterilized fruits were inoculated onto the prepared medium with different combinations of growth hormones. The shoot-tip culture was also performed by using explants derived from in vitro raised seedlings to determine the most suitable phytohormone combinations and concentrations for maximum rooting and leaf formation. The cultures were maintained inside culture room in controlled condition with proper light illumination of 60 μmol m-2 sec-1 for 12 h a day using white fluorescent tubes and temperature at 25±2°C.
Hardening, data recording and synthetic seed production: The in vitro regenerated tomato plants with healthy leaves and well-developed roots were taken out from culture flasks and transferred to miniature pots containing autoclaved sand and soil (1:1) as potting mixture. The transplanted plants were kept in glass house for further acclimatization to nursery condition. The experiment was conducted twice with four replicates per hormonal combinations. The observation was made at regular intervals after every week and number of leaf and root development in each treatment was recorded. The data obtained from the present investigation was subjected to Analysis of Variance (ANOVA) and significant differences were determined by employing Duncan’s multiple range test at p = 0.05 (Duncan, 1955). The statistical data analysis was performed using SPSS software program (SPSS Inc., Chicago, USA).
Propagules for synthetic seed development were obtained by excising shoot tips of 3-5 mm length from in vitro raised tomato plants. Sodium alginate solutions of different concentrations (2, 3 and 4%) were prepared by adding alginate powder to calcium free liquid MS medium which were subsequently mixed with excised shoot-tip propagules. The CaCl2 solutions of 25, 50, 75 and 100 mM were separately prepared for gel complexation with sodium alginate of varied concentrations. The explants (micro shoots of in vitro regenerated tomato plants) were sucked with a pipette along with sodium alginate solution and dropped into calcium chloride solution. They were incubated for different time intervals of 10-40 min during which the ion exchange reaction occurred and sodium ions were replaced by calcium ions forming calcium alginate beads. The whole process must be done under aseptic conditions and size of alginate capsule depends on the inner diameter of pipette nozzle. The beads were recovered by decanting the CaCl2 solution and newly formed synseeds were washed several times with sterile water to remove traces of CaCl2. The uniform beads thus obtained were stored at 4°C in sterile condition.
RESULTS AND DISCUSSION
The excised seeds which were free from any surrounding fruit tissues were planted on to the MS medium supplemented with plant growth regulators of different combinations and concentrations. Germination of seeds started 3-4 days after inoculation as sprouting of seeds was observed in many hormonal combinations tested. Bhattarai et al. (2009) observed similar response of seed germination in tomato when germinating seeds sprouted to form seedlings under the influence of plant growth hormones. The germinated seeds differentiated into multiple shoots and leaves without root formation on MS+2.5 mg L-1 BAP (Fig. 1a).
 | |
| Fig. 1(a-f): | In vitro generation response of Lycopersicon esculentum Mill. and synthetic seed development, (a) Sprouting germinating seeds differentiating into shoots in MS+2.5 mg L-1BAP, (b) In vitro leaf multiplication in medium supplemented with 1.5 mg L-1 NAA+2.5 mg L-1 BAP+2.5 mg L-1 IBA, (c) High rate of root proliferation in 1.5 mg L-1 NAA enriched medium, (d) In vitro shoot tip growth and differentiation into multiple leaves in tomato culture, (e) Transplanted tomato plantlet in miniature pot containing autoclaved sand and soil (1:1) as potting mixture and (f) Firm and transparent round shaped tomato synthetic seeds developed by encapsulating shoot tip propagules with gel matrix derived from 2% sodium alginate with 100 mM CaCl2 solution |
Sun et al. (2006) also observed the failure of root production in tomato culture in absence of exogenous auxins in culture medium. Incorporation of 0.4% AC in same combination deteriorated the culture response towards formation of leaves and roots after seed germination. Seedling formation was not observed in MS+2.5 mg L-1 BAP+AC even after two subsequent passages and 10 weeks of culture. Pico et al. (2002) also reported germinated embryos failing to differentiate into functional plants despite successful formation of shoots in initial stage of culture. Addition of 0.5 mgL-1 NAA along with 3 mg L-1 BAP was found to be conducive for effective rooting and leaf multiplication with formation of 1.5±1.0 and 2.0±1.3 number of roots and leaves, respectively (Table 1). Presence of AC (0.4%) in above hormonal combination did not produce any seedling though the germinated seeds formed roots. This proved the detrimental effect of AC for in vitro growth and differentiation of culture if present along with higher concentration of BAP. The reducing effect of AC on culture development of tomato was not in line with previous findings in other plants where it always had promontory role in plantlet regeneration (Pan and van Staden, 1998; Nongdam and Chongtham, 2012).
| Table 1: | In vitro culture of Lycopersicon esculentum using seed explants for leaf and root generation |
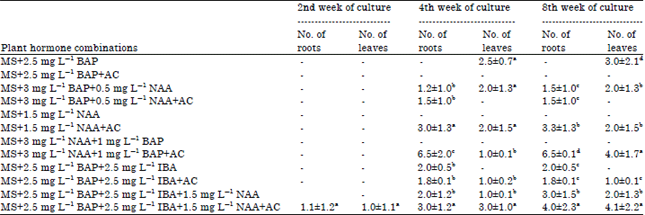 | |
Results are based on 4 replicates per treatment in two repeated experiments. Means followed by same letter are not significantly different at p = 0.05, hyphen (-) indicates the absence of in vitro root or leaf formation | |
MS medium augmented with 1.5 mg L-1 of NAA was found to be nonresponsive to seed germination as none of the seeds showed any sign of germination. However enrichment of medium with 0.4% AC and 1.5 mg L-1 of NAA significantly improved seed germination response and subsequent growth leading to complete seedling development with healthy leaf (2.0±1.5) and root (3.3±1.3) formation. The beneficial property of AC in promoting seed germination and culture growth had earlier been reported in different plants (Pacek-Bieniek et al., 2010; Benmahioul et al., 2012). The importance of AC for in vitro seed germination and seedling development might be due to its ability to adsorb phenolic exudates, aerate medium and provide near natural conditions for seed germination by absorbing the light (Ernst, 1975). The MS+3 mg L-1 NAA+1 mg L-1 BAP was also proved unsuitable for seed germination and plantlet regeneration in vitro as no growth response was observed in this hormonal combination. But addition of 0.4% AC in same phytohormone combination exhibited accelerated in vitro morphogenetic response producing maximum root formation (6.5±0.1) and better leaf production (4.0±1.7). The above observation indicated the non-responsiveness of tomato culture to higher concentration of NAA when AC was absent in medium. The promotory effect of NAA in rapid root induction under the influence of AC was earlier described in orchid by Nongdam and Chongtham (2011). MS medium supplemented with equal concentration of both BAP and IBA at 2.5 mg L-1 did not generate any functional plants though shoots and roots were differentiated from the germinating seeds. Inclusion of 0.4% AC in the same combination gave the required growth stimulus for in vitro production of plants with better leaf and root development. Presence of 1.5 mg L-1 of NAA along with 2.5 mg L-1 BAP+2.5 mg L-1 IBA elevated seed germination response and subsequent differentiation to complete seedlings with healthy leaves and roots. The in vitro shooting response was convincingly high with AC in the above growth regulator combination producing rapid leaf multiplication (Fig. 1b).
Shoot tips excised from seed derived tomato seedlings were used as explants for culture initiation on medium enriched with different combinations and doses of plant growth hormones. MS medium supplemented with 2.5 mg L-1 BAP+AC produced 5.5±2.5 number of leaves though root development was not observed (Table 2). This indicated the inability of BAP in inducing rooting in tomato in absence of exogenous auxins in medium. The finding is in contrast to the observation made by Kartha et al. (1976) which showed adventitious root formation in tomato culture on cytokinin enriched medium devoid of any auxins. However, root formation was exceedingly high (9.5±2.6) with better root differentiation in medium incorporated with 1.5 mg L-1 NAA (Fig. 1c).
| Table 2: | Shoot tip culture from in vitro raised tomato plants for leaf and root regeneration using different combinations and doses of phytohormones |
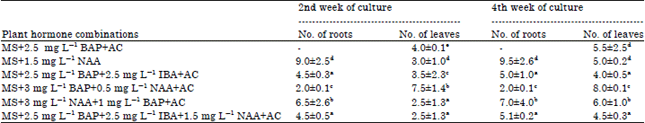 | |
Results are based on 4 replicates per treatment in two repeated experiments. Means followed by same letter are not significantly different at p = 0.05, hyphen (-) indicates the absence of in vitro root or leaf formation | |
The dramatic rise in culture growth especially root formation was quite astonishing as the germination response was totally absent when seed explant was used to initiate tomato culture. This might be due to NAA exhibiting unfavourable response to seed germination but its promotory influence on rapid rooting induction was expressed with the use shoot -tip explants. The stimulatory effect of NAA in rooting multiplication was previously described in tomato and Dendrobium orchid (Davis et al., 1994; Ajenifujah-Solebo et al., 2012; Nongdam and Tikendra, 2014). Medium enriched with 3 mg L-1 BAP+0.5 mg L-1NAA+AC induced rapid leaf proliferation producing healthy leaves (Fig. 1d). Though the above hormonal combination generated maximum leaves (8.0±0.1), it did not produce any leaves when seed explants were used. This indicated the inability of AC in providing a conducive culture environment for effective seed germination and subsequent shoot differentiation to leaves when BAP was present in higher concentration. The in vitro constraints of germinating seeds in developing into roots and leaves in the hormonal combination were removed when shoot-tips were used as explants. There have been similar reports of accelerated shoot multiplication and leaf development in tomato culture when BAP was supplemented in the MS medium (Uddin et al., 2004; Harish et al., 2010). The MS+3 mg L-1 NAA+1 mg L-1 BAP+AC and MS+2.5 mg L-1 BAP+2.5 mg L-1 IBA+1.5 mg L-1 NAA+AC displayed good shooting and rooting response though leaf and root formation were not significantly different from what had been observed with seed explants. The tomato plantlets having well established roots and leaves were removed from culture vessels and transplanted into small pots containing autoclaved sand and soil (1:1) as potting mixture (Fig. 1e). The transplanted plants were kept in glass house for further acclimatization to nursery condition.
The shoo-tip propagules obtained from in vitro raised tomato seedlings were encapsulated using different concentration of sodium alginate (2, 3 and 4%) and CaCl2 (25, 50, 75 and 100 mM) at interval of 10, 20, 30 and 40 min treatment duration. The 2% sodium alginate when complexed with 25 mM CaCl2, 50 mM CaCl2 and 75 mM CaCl2 did not produce favourable matrix for efficient encapsulation as soft and transparent gel was produced (Table 3). Treatment of 2% sodium alginate with 100 mM CaCl2 at treatment duration of 20, 30 and 40 min generated desirable firm and transparent encapsulating gel (Fig. 1f). Similar observation was made in synthetic seed development of other plants (Nagananda et al., 2011; Maqsood et al., 2012). However, Saiprasad and Polisetty (2003) observed complexation of 3% sodium alginate with 75 mM CaCl2 for 20-30 min as optimum combination for proper hardening of beads in different orchids. Also Mohanty et al. (2013) found 3% sodium alginate solution with 10 mM CaCl2 as favourable proper synthetic seed formation in Dendrobium orchids. The increased in sodium alginate concentration at 3% along with gradual raised in CaCl2 at 25, 50, 75 and 100 mM produced soft, firm and very less transparent beads which were not suitable for encapsulating plant propagules.
| Table 3: | Effect of different concentrations of sodium alginate and calcium chloride on nature of synthetic seeds developed at varied treatment duration |
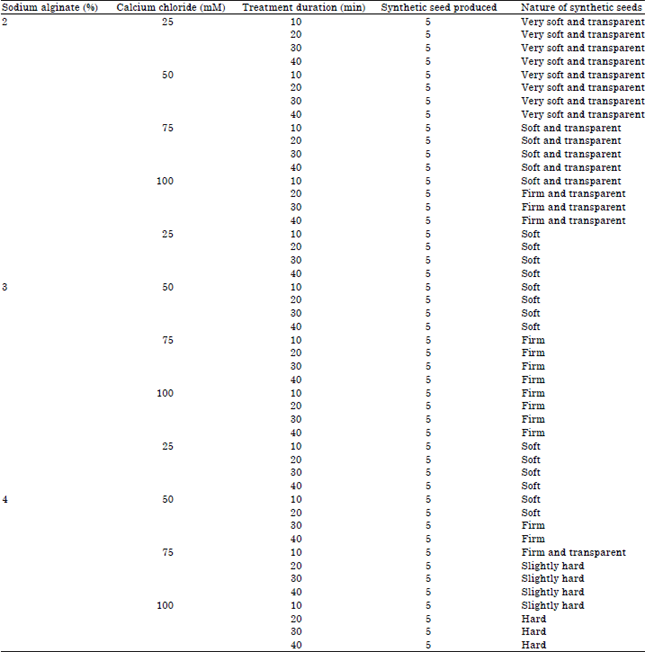 | |
The use of 4% sodium alginate with higher level of CaCl2 generated unwanted firm and very hard beads which were often short-lived. All forms of synthetic seeds obtained from the present work were stored at 4°C under sterile condition to check bead quality and durability. The very soft, hard and less transparent synseeds did not survive as they could not maintained their structure either due to high fragile nature of seed coat or over desiccation of hard covering. Those synseeds with firm and transparent encapsulating matrix can retain their shape and quality for longer duration of about 2-3 months at lower temperature of 4°C.
The present study revealed the differential in vitro response of tomato culture under varied hormonal conditions in medium. Leaf and root development were absent at 2 weeks of culture in most of phytohormones combinations tested even though seeds were germinated successfully. However the germinated seeds undergone further growth and morphogenetic differentiation leading to significant rise in leaf and root formation in 8 weeks of culture (Fig. 2). BAP was found to be most favourable for producing high seed germination and leaf multiplication though presence of AC hampered its influence on culture growth. The NAA was also found to be effective in inducing seed germination leading to rapid root proliferation. The MS medium supplemented with 3 mg L-1 NAA+1 mg L-1 BAP+AC produced maximum rooting for culture initiated from germinating seeds. The shoot-tip culture produced significantly higher leaf and root formation as compared to cultures initiated from seed explants (Fig. 3). The influence of BAP in promoting shooting response was high with maximum leaf formation recorded in MS+3 mg L-1 BAP+0.5 mg L-1 NAA+AC. The NAA also proved to be very effective in accelerating root multiplication with highest number of roots observed in medium augmented with 1.5 mg L-1 NAA.
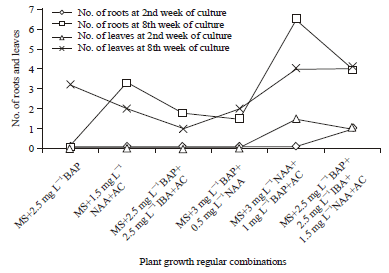 | |
| Fig. 2: | In vitro development of roots and leaves of tomato from shoot tip culture under different hormonal combinations and concentrations |
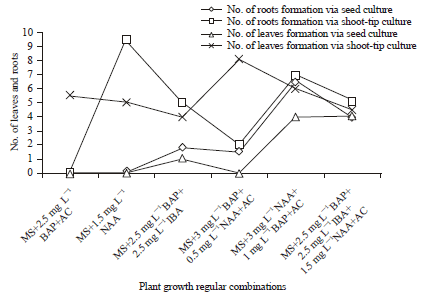 | |
| Fig. 3: | In vitro root and leaf development of tomato from seed and shoot tip culture under different growth regulator combinations and concentrations |
The preliminary investigation on synthetic seed development of tomato affirmed the complexation of 2% sodium alginate with 100 mM CaCl2 for 20-40 min duration as the most favourable condition for production of favourable transparent and firm gel beads.
REFERENCES
- Ajenifujah-Solebo, S.O.A., N.A. Isu, O. Olorode, I. Ingelbrecht and O.O. Abiade, 2012. Tissue culture regeneration of three Nigerian cultivars of tomatoes. Afr. J. Plant Sci., 6: 370-375.
CrossRefDirect Link - Benmahioul, B., M. Kiad-Harche and F. Daguin, 2012. Influence of activated charcoal on in vitro embryo germination and growth of plantlets of pistachio (Pistacia vera L.). Int. J. Sci. Nat., 3: 613-616.
Direct Link - Bhattarai, S.P., R.C. de la Pana, D.J. Midmore and K. Palchamy, 2009. In vitro culture of immature seed for rapid generation advancement in tomato. Euphytica, 167: 23-30.
CrossRefDirect Link - Bhatia, P., N. Ashwath and D.J. Midmore, 2005. Effects of genotype, explant orientation and wounding on shoot regeneration in tomato. In vitro Cell. Dev. Biol.-Plant, 41: 457-464.
CrossRefDirect Link - Bhatia, P., N. Ashwanth, T. Senaratna and D. Midmore, 2004. Tissue culture studies of tomato (Lycopersicon esculentum). Plant Cell. Tissue Organ Cult., 78: 1-21.
CrossRefDirect Link - Chandra, I., P. Singh, A. Bhattacharya, P. Singh, S. Javed and A. Singhamahapatra, 2013. In vitro callus induction, regeneration and micropropagation of Solanum lycopersicum. Int. J. Curr. Microbiol. Applied Sci., 2: 192-197.
Direct Link - Harish, M.C., S. Rajeevkumar and R. Sathishkumar, 2010. Efficient in vitro callus induction and regeneration of different tomato cultivars of India. Asian J. Biotechnol., 2: 178-184.
CrossRefDirect Link - Hassanein, A., I.A. Ibraheim, A.N. Galal and J. Salem, 2011. Conversion of banana synseed influenced by the bead type and seed coat. Am. J. Plant Sci., 2: 467-475.
CrossRefDirect Link - Himabindu, K.B., M.S. Priya, D.M. Reddy, P. Sudhakar and Y. Srinivasulu et al., 2012. Studies on the effect of various sterilants and culture conditions on in-vitro seed germination in tomato (Solanum lycopersicum). Int. J. Applied Biol. Pharmaceut. Technol., 3: 476-480.
Direct Link - Hossain, M.N., M. Fakruddin and M.N. Islam, 2011. Effect of chemical additives on the shelf life of tomato juice. Am. J. Food Technol., 6: 914-923.
CrossRefDirect Link - Huda, A.K.M.N., M. Rahman and M.A. Bari, 2007. Effect of carbon source in alginate bead on synthetic seed germination in eggplant (Solanum melongena L.). J. Plant Sci., 2: 538-544.
CrossRefDirect Link - Lima, J.E., R.F. Carvalho, A.T. Neto, A. Figueira and L.E. Peres, 2004. Micro-MsK: A tomato genotype with miniature size, short life cycle and improved in vitro shoot regeneration. Plant Sci., 167: 753-757.
CrossRefDirect Link - Maqsood, M., A. Mujib and Z.H. Siddiqui, 2012. Synthetic seed development and conversion to plantlet in Catharanthus roseus (L.) G. Don. Biotechnology, 11: 37-43.
CrossRefDirect Link - Mohanty, P., P. Nongkling, M.C. Das, S. Kumaria and P. Tandon, 2013. Short-term storage of alginate-encapsulated protocorm-like bodies of Dendrobium nobile Lindl.: An endangered medicinal orchid from North-East India. Biotech, 3: 235-239.
CrossRefDirect Link - Mohanraj, R., R. Ananthan and V.N. Bai, 2009. Production and storage of synthetic seeds in Coelogyne breviscapa lindl. Asian J. Biotechnol., 1: 124-128.
CrossRefDirect Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Nagananda, G.S., N. Satishchandra and S. Rajath, 2011. Regeneration of encapsulated protocorm like bodies of medicinally important vulnerable orchid Flickingeria nodosa (Dalz.) Seidenf. Int. J. Bot., 7: 310-313.
CrossRefDirect Link - Nair, R.R. and S.D. Gupta, 2007. In vitro plant regeneration from encapsulated somatic embryos of black pepper (Piper nigrum L.). J. Plant Sci., 2: 283-292.
CrossRefDirect Link - Nongdam, P. and L. Tikendra, 2014. Establishment of an efficient in vitro regeneration protocol for rapid and mass propagation of Dendrobium chrysotoxum Lindl. using seed culture. J. Sci. World.
CrossRefDirect Link - Nongdam, P. and N. Chongtham, 2012. In vitro seed germination and mass propagation of Cymbidium dayanum Reichb.: An important ornamental orchid of North-East India. Trends Hortic. Res., 2: 28-37.
CrossRefDirect Link - Nongdam, P. and N. Chongtham, 2011. In vitro rapid propagation of Cymbidium aloifolium (L.) Sw.: A medicinally important orchid via seed culture. J. Biol. Sci., 11: 254-260.
CrossRefDirect Link - Pacek-Bieniek, A., M. Dyduch-Sieminska and M. Rudas, 2010. Influence of activated charcoal on seed germination and seedling development by the asymbiotic method in Zygostates grandiflora (Lindl.) Mansf. (Orchidaceae). Folia Hortic., 22: 45-50.
CrossRefDirect Link - Pan, M.J. and J. van Staden, 1998. The use of charcoal in in vitro culture: A review. Plant Growth Regul., 26: 155-163.
CrossRefDirect Link - Patel, A.V., I. Pusch, G. Mix-Wagner and K.D. Vorlop, 2000. A novel encapsulation technique for the production of artificial seeds. Plant Cell Rep., 19: 868-874.
CrossRefDirect Link - Piccioni, E., 1997. Plantlets from encapsulated micropropagated buds of M.26 apple rootstock. Plant Cell Tissue Org. Cult., 47: 255-260.
CrossRefDirect Link - Pico, B., J. Herraiz, J.J. Ruiz and F. Nuez, 2002. Widening the genetic basis of virus resistance in tomato. Scientia Hortic., 94: 73-89.
CrossRefDirect Link - Mamidala, P. and R.S. Nanna, 2011. Effect of genotype, explant source and medium on in vitro regeneration of tomato. Int. J. Genet. Mol. Biol., 3: 45-50.
Direct Link - Saiprasad, G.V.S. and R. Polisetty, 2003. Propagation of three orchid genera using encapsulated protocorm-like bodies. In vitro Cell. Dev. Biol.-Plant, 39: 42-48.
CrossRefDirect Link - Sun, H.J., S. Uchii, S. Watanabe and H. Ezura, 2006. A highly efficient transformation protocol for Micro-Tom, a model cultivar for tomato functional genomics. Plant Cell Physiol. 47: 426-431.
CrossRefPubMedDirect Link - Kartha, K.K., O.L. Gamborg, J.P. Shyluk and F. Constabel, 1976. Morphogenetic investigations on in vitro leaf culture of tomato (Lycopersicon esculentum Mill. cv. Starfire) and high frequency plant regeneration. Zeitschrift fur Pflanzenphysiologie, 77: 292-301.
CrossRefDirect Link - Singh, T., N. Singh, A. Bahuguna, M. Nautiyal and V.K. Sharma, 2014. Performance of tomato (Solanum lycopersicum L.) hybrids for growth, yield and quality inside polyhouse under mid hill condition of Uttarakhand. Am. J. Drug Discov. Dev., 4: 202-209.
CrossRefDirect Link - Uddin, M.F., M.A. Taher, S.M.A. Islam and M.Z. Hossain, 2004. Effect of variety and plant growth regulators in MS medium on shoot induction from virus infected calli of tomato. J. Biol. Sci., 4: 521-526.
CrossRefDirect Link