
Deborah Vicuna Requesens
Arkansas Biosciences Institute, Arkansas State University, P.O. Box 639, AR 72467, USA
Renee P. Malone
Arkansas Biosciences Institute, Arkansas State University, P.O. Box 639, AR 72467, USA
Philip J. Dix
Department of Biology, National University of Ireland Maynooth, Maynooth, Co. Kildare, Ireland
Journal of Plant Sciences
Year: 2014 | Volume: 9 | Issue: 2 | Page No.: 58-66







ABSTRACT
Heterologous expression of peroxidase genes has been shown to influence morphology and stress responses of several crop plants but little is known about the effect in woody species. In this study, a barley cell-wall peroxidase gene (prx8), peviously shown to influence growth and stress tolerance in tobacco, was introduced into the genome of apple (Malus domestica cv. Greensleeves) via Agrobacterium-mediated transformation and the presence of the transgene confirmed by PCR and Southern blot analysis. The transgenic plants had up to 4-fold higher levels of peroxidase activity compared to wild type plants and exhibited faster growth and increased xylem production. Leaf discs were incubated at high temperatures (44°C) and electrolyte leakage measurements indicated enhanced tolerance against temperature stress. This effect was confirmed when whole plants were subjected to heat stress after 6 and 12 months growth in soil. These results indicate a link between peroxidase activity levels and resistance to thermal stress, as well as biochemical and physiological changes, in this woody plant.
PDF Abstract XML References Citation
Received: February 05, 2014;
Accepted: May 14, 2014;
Published: May 28, 2014
How to cite this article
Deborah Vicuna Requesens, Renee P. Malone and Philip J. Dix, 2014. Expression of a Barley Peroxidase in Transgenic Apple (Malus domestica L.) Results in Altered Growth, Xylem Formation and Tolerance to Heat Stress. Journal of Plant Sciences, 9: 58-66.
DOI: 10.3923/jps.2014.58.66
URL: https://scialert.net/abstract/?doi=jps.2014.58.66
DOI: 10.3923/jps.2014.58.66
URL: https://scialert.net/abstract/?doi=jps.2014.58.66
INTRODUCTION
Class III peroxidases (EC 1.11.1.7) are involved in a wide range of biochemical and physiological processes including lignification, suberisation, auxin oxidation and the cross-linking of molecules in the cell wall (Passardi et al., 2004). Transgenic tobacco plants with altered peroxidase expression presented modified levels of lignin compared to wild type plants (Lagrimini et al., 1997; Quiroga et al., 2000; Heggie et al., 2005). Peroxidases have also been linked to defense mechanisms against a variety of abiotic stresses (Fang and Kao, 2000; Amaya et al., 1999). Elevated peroxidase activities in transgenic plants have resulted in increases in tolerance to several stresses including high temperatures, osmotic (Amaya et al., 1999) and oxidative (Kawaoka et al., 2003) stress. In a recent report (Vicuna et al., 2011) a defence-related cell wall peroxidase of barley, prx8, proved particularly effective in enhancing tolerance to several different abiotic stresses, when over-expressed in transgenic tobacco.
In view of the well-known effects of elevated peroxidase activities on metabolism of phenolics and lignification, as well as on stress tolerance, it was of interest to explore the consequences of over-expression of a cell wall peroxidase in a woody crop. For the current investigation with apple, we chose to express the barley peroxidase gene (prx8) previously explored in tobacco (Vicuna et al., 2011). This gene is induced in response to powdery mildew fungus attack in barley (Thordal-Christensen et al., 1992) and its over-expression has been found to increase lignin deposition in transgenic tobacco plants (Burbridge, unpublished results). Here we report that over-expression of this barley peroxidase gene in Malus domestica cv. ‘Greensleeves’ plants results in profound changes in plant growth and xylem formation, as well as greatly enhanced tolerance to heat stress.
MATERIALS AND METHODS
Location: The experimental work was performed in the Plant Cell Biology Laboratory, National University of Ireland Maynooth, between 1st September 2002 and 31st March 2005.
Plant material: Malus domestica L., cv. Greensleeves shoot cultures were grown under 16/8 h light/dark photoperiod regime at 22°C. In vitro grown plants were maintained on Murashige and Skoog (MS) medium (Murashige and Skoog, 1962) supplemented with 1 mg L-1 benzylaminopurine (BAP) and 0.1 mg L-1 Indole Butyric Acid (IBA). For rooting, shoots were transferred to hormone-free MS medium until roots were visible and then planted in John Innes multi-purpose compost and grown in growth rooms under the same temperature and day length conditions as the shoot cultures.
Vector construction: A transformation vector containing the barley prx8 gene was employed for transformations (Vicuna et al., 2011). The entire prx8 sequence was amplified from vector pBKL4-301 (Kristensen et al., 1997), KpnI or SacI restriction sites were added on either end of the gene and it was ligated into the binary vector pCAMBIA3201 to create pCAMBIA3201/prx8 which was then transformed into Agrobacterium tumefaciens strain C851 as described by James and Dandekar (1991). The prx8 gene was under the control of the cauliflower mosaic virus (CaMV) 35S promoter and terminator and the vector included a phosphinothricin resistance gene (bar) expressed from the same promoter.
Genetic transformation: The procedure for stable transformation was as described by James and Dandekar (1991). Briefly, leaf discs (0.5 cm diameter) from in vitro grown apple plants were co-cultivated with A. tumefaciens strain C851 containing the vector described above for 20 min then blotted and placed on co-cultivation medium at 25°C in darkness for 3 days. The explants were then placed on selection medium containing 5 mg L-1 Bialaphos. Resistant shoots were excised and placed on apple rooting medium (James and Dandekar, 1991) containing 5 mg L-1 Bialaphos.
DNA isolation, PCR detection and Southern analysis: Total genomic DNA was isolated from apple leaves according to Frey (1999). Forty microgram of DNA was digested with BamH1 and resolved on a 0.8% agarose gel, then blotted onto a nylon membrane (Hybond N, Amersham). The blot was probed for the prx8 gene, following a non-radioactive method (McCabe et al., 1997). The prx8 probe was DIG-labelled according to the manufacturer’s instructions (Invitrogen, Carlsbad, CA, USA). Hybridization was carried out for 16 h at 42°C. The blot was then washed in 2X SSC, containing 0.1% SDS, at room temperature and 0.1X SSC, containing 0.1% SDS, at 65°C. It was then blocked and incubated in anti-DIG-AP (Sigma Chemical Co., St. Louis, MO, USA) and finally incubated in 1, 2-dioxetane chemiluminescent substrate (CDPstar, Sigma Chemical Co.).
PCR analyses were carried out on 200-300 ng genomic DNA using 5’-ATGGCTTCTTCATAC-3' and 5’-TCACGAGTTCACCCTGGAGCAGCT-3’ as primers. An initial step of 94°C for 5 min was followed by 40 cycles of 94°C for 1 min 55°C for 1 min and 72°C for 2 min with a final cycle cycle of 10 min at 72°C.
mRNA analysis: RNA was isolated from leaf material by grinding 500 mg with 500 μL lysis buffer (0.1 M Tris HCl, pH 8.0, 50 mM EDTA, 50 mM NaCl, 2% SDS, 0.75% β-mercaptoethanol) and 500 μL buffer-saturated phenol. The lysate was extracted once with phenol and twice with phenol:chloroform (1:1), followed by extraction in 100% chloroform. RNA was precipitated with 8 M LiCl and washed with 0.1% DEPC in 70% ethanol. RT-PCR was performed with the isolated RNA. The Qiagen Omniscript™ Reverse Transcriptase kit for 1st strand cDNA synthesis was used.
Protein content and enzyme assays: Total soluble protein was extracted from plant material (100 mg mL-1) using extraction buffer (50 mM sodium phosphate buffer pH 7.0, 1 mM EDTA, 1% v/v Triton X-100, 1% PVP). The extract was filtered through two layers of Miracloth and total protein was measured according to a standard method (Biorad Hercules, CA, USA), using bovine serum albumin as the standard protein. To determine peroxidase activity, 50 μg protein extract was mixed with 1 mL of a solution containing 0.3 mM guaiacol and 0.1 mM H2O2 in 100 mM Na2HPO4, pH 7.0. The increase in absorbance at 436 nm was followed for 3 min at 30 sec intervals. The rate of this reaction was used to express peroxidase activity.
Xylem staining: Transverse sections of petioles, taken from the tenth node from the top of each plant, at 1 cm from the main stem, were stained with phloroglucinol solution (5% v/v phloroglucinol in 78% ethanol). One drop of phloroglucinol solution was added to each section, left for 4 min and then followed by one drop of concentrated HCl. Lignin stained pink and sections were examined under the 4X lens of a Leica microscope. Samples were stored at -70°C after flash freezing in liquid nitrogen. All solutions were DEPC treated.
Electrolyte leakage: Leaf discs (1 cm diameter), cut with a cork borer were washed with deionised water and kept at 44°C for 8 h in deionised water. The electrolyte leakage (EC) was measured immediately after the temperature stress (ECinitial) using an EC215 conductivity meter (Hanna Instruments). The bathing solution (deionised water) containing the leaf discs was then boiled for 5 min and ECtotal was measured. The Index of Injury (Id) was determined from the following equation by Flint et al. (1966):
where, Rt = ECinitial/ECtotal for stressed tissues and R0=ECinitial/ECtotal for non-stressed tissues.
Heat stress of soil-grown plants: Four-week-old seedlings and 1 year old plants were transferred to 35 and 37°C, respectively, for 24 h. Plants were returned to ambient temperature to recover.
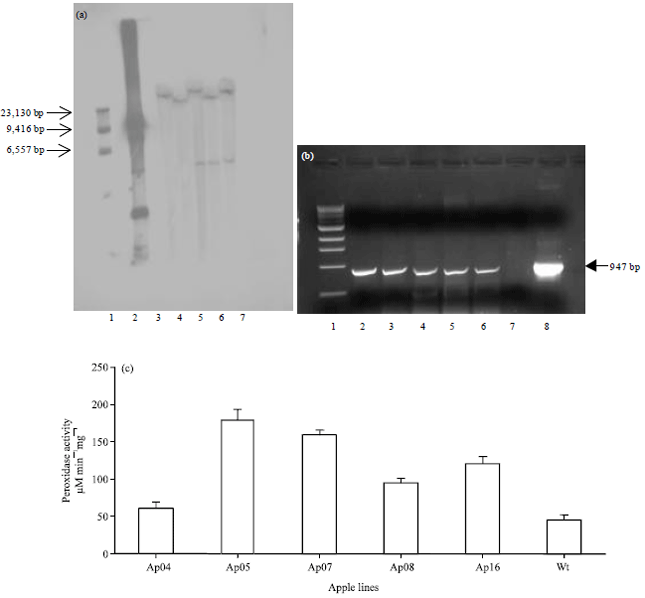 | |
| Fig. 1(a-c): | Molecular analysis and transgene expression in transgenic apple plants, (a) Southern blot analysis, Apple transgenic lines transformed with the prx8 vector via., Agrobacterium-mediated transformation. Lane 1: λ/HindIII ladder; lane 2: Positive control, lanes 3-7: Positive apple transgenic lines Ap04, Ap05, Ap07, Ap08 and Ap16, lane 8: Wild type apple, blot was hybridized with DIG-labelled prx8 probe, (b) RT-PCR products from prx8 transgenic apple plants. Lane 1: 1 Kb ladder (Promega), lanes 2-6: Positive transgenic apple lines Ap 04, Ap05, Ap07, Ap08 and Ap16, lane 7: Wild type tobacco, lane 8: Pcontrol and (c) Peroxidase activity levels, transgenic apple lines Ap04, Ap05, Ap07, Ap08 and Ap16 as well as wild type (wt) apple plants were tested for peroxidase activity |
RESULTS
PCR, Southern blot and gene expression analysis: The presence of the prx8 gene in the transgenic lines was shown by PCR and confirmed by Southern blot analysis (Fig. 1a). All independent transgenic lines tested contained one or two copies of the prx8 gene. RNA isolated from the transformed lines was used as the template for RT-PCR analyses (Fig. 1b). The cDNA obtained from prx8 mRNA is represented by a band of 947 bp. No corresponding cDNA was obtained from non-transformed wild-type plants.
Figure 1c, shows the peroxidase activities of T0 transgenic lines that were positive for the insertion of the prx8 gene. When compared to wild type plants, all the transgenic lines show higher levels of peroxidase activity, with activity levels varying from 1.3-fold (Ap04) to 4-fold increases (Ap05).
Growth and development of transgenic plants: Four weeks after rooting and transfer to soil, all the transgenic lines showed signs of faster development (Fig. 2a), compared with non-transformed plants. After 6 months, all the transgenic plants were taller than the wild-type (Fig. 2b). The mean height of wild type plants was 82 cm while that of transgenic plants varied between 120 (Ap05) and 163 (Ap08) cm.
 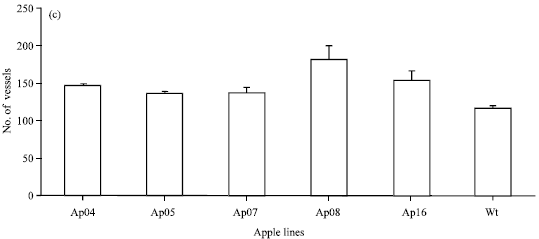 | |
| Fig. 2(a-c): | Growth and xylem production in transgenic apple plants. Three plants of each transgenic line and three non-transformed control lines, were grown in soil. Representative plants are shown: (a) Four-week old and (b) Six-month (height scale in cm.) old plants L-R: Transgenic lines Ap04, Ap05, Ap07, Ap08 and Ap16 and wild type apple plant and (c) Xylem vessel number. The number of xylem vessels were counted in transverse petiole sections of prx8 transgenic apple plants Ap04, Ap05, Ap07, Ap08 and Ap16 and wild type apple plants. Data are the mean of 10 different sections, taken from each of three plants for each line, Bars represent the standard error of the mean |
Transgenic apple plants also had a higher number of nodes (58-72) when compared to the wild-type apple plants (33).
Sections from petioles taken from the tenth node of six-month old plants were stained and photographed and the number of xylem vessels or fibres counted (Fig. 2c). The mean number of vessels/fibres is higher in all the transgenic lines when compared to wild-type plants.
Effects of high temperature stress: Electrolyte leakage measurements on leaf discs (Fig. 3a) showed that after heat stress smaller Ids were obtained for all the plants over-expressing the peroxidase gene. One-month old apple plants were incubated at 35°C overnight. After 24 h, transgenic plants over-expressing the prx8 gene showed no signs of heat stress (Fig. 3b) and had intact green leaves. Wild type apple plants showed damage caused by heat, with brown areas and dehydrated leaves. When 1-year old plants were subjected to 37°C overnight and then returned to normal growth conditions, almost complete loss of leaves occurred in the wild type plants, while all the transgenic lines retained most of their leaves (data not shown).
DISCUSSION
Six transgenic apple plants expressing the barley prx8 gene were obtained in the present study. All the transgenic plants showed increased but differing peroxidase levels, the differences being unrelated to transgene copy number. The prx8 gene product is targeted to the cell wall (Kristensen et al., 1997), where it is likely to influence development and defense responses. In a previous study, we transformed tobacco plants with the same prx8 gene and showed an increase in peroxidase activity levels of between 2 and 4.5 times (Vicuna et al., 2011).
Apple plants over-expressing the prx8 gene showed a faster rate of growth when compared to the wild type plants. The influence of peroxidase activity on tree growth rate has been studied previously.
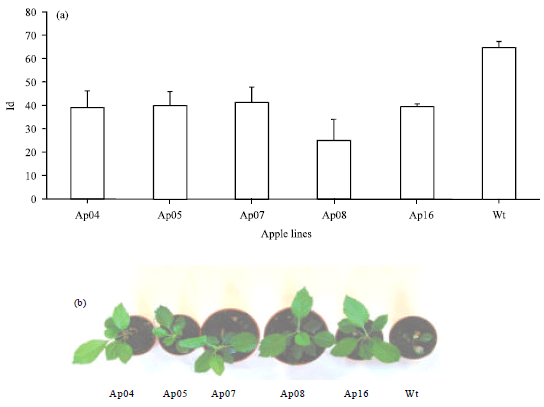 | |
| Fig. 3(a-b): | Response to heat stress in transgenic plants, (a) Index of Injury (Id) obtained by conductivity measurements of leaf discs of wild type and transgenic apple plants overexpressing the prx8 gene, exposed at 44°C, after 8 h, transgenic plants are labelled Ap04, Ap05, Ap07, Ap08 and Ap16. Values are means of values from ten leaf discs taken from each of three plants and the bars represent the standard errors of the mean and (b) Representative four-week old apple plants, photographed after being incubated at 35°C overnight |
Kawaoka et al. (2003) analyzed transgenic aspen trees over-expressing a horseradish peroxidase (prxC1a) and demonstrated that this increased activity stimulated growth. The authors suggest that the product from the prxC1a gene has an activity toward ascorbate and that it might act as an ascorbate oxidizer in the process of cell elongation. Peroxidase levels may also influence plant architecture through the established effect they have on Indole Acetic Acid (IAA) levels (Normanly, 1997).
Another factor which may contribute to the results obtained here is the involvement of peroxidases in the process of lignifications (Quiroga et al., 2000). In order to study the influence of peroxidase over-expression on lignification and xylogenesis, sections of petioles from apple plants over-expressing the prx8 gene as well as wild type apple plants, were stained with phloroglucinol to visualise lignin. The transgenic plants exhibited a higher number of xylem vessels and/or fibres in these sections. By stimulating the lignification process, the increase in peroxidase levels found in the present study could influence cytodifferentiation, leading to the observed increase in xylem vessels. Previous investigations have shown a correlation between peroxidase levels and lignin content. Quiroga et al. (2000) studied the correlation between the expression of a tomato peroxidase in tobacco plants and the synthesis of lignin. Expression of this peroxidase was found mainly in cells undergoing active lignification. To our knowledge the present results are the first to demonstrate a relationship between recombinant peroxidase production and xylem formation in a woody species. Several studies show a correlation of heat stress with the production of reactive oxygen species (Jiang and Huang, 2001). Transgenic apple plants over-expressing the cell wall peroxidase from barley showed an increased resistance to heat stress, both at a cellular level (reduced membrane damage) and in whole plants. Previous studies have shown that heat stress enhances the activity of enzymes capable of regulating the levels of ROS in plants, including peroxidases (Edreva et al., 1998; Chaitanya et al., 2002; Gulen and Eris, 2004). Recent studies with tobacco suggest that type III peroxidases (such as the barley peroxidase prx8) can have a role in tolerance to several abiotic stresses, including heat (Vicuna et al., 2011). The present study illustrates that these findings, obtained with herbaceous plants, can be extended to tree species.
The results discussed in this study contribute to a better understanding of the biochemical and physiological role of peroxidases in plants. They could also be of economic interest in the production of crops with improved agronomic traits, such as increased resistance to harsh abiotic stress conditions as well as more environmentally friendly crops to be used in the paper and pulp industry.
ACKNOWLEDGMENTS
We would like to thank Dr. Soren Rassmussen at Risoe National Laboratory, Denmark for supplying the prx8 clone and Dr. Ilona Dix for assistance in preparing the figures. We would also like to extend our appreciation to Dr. Elizabeth E. Hood, Arkansas State University, for her encouragement and her invaluable assistance in the preparation of this manuscript.
REFERENCES
- Chaitanya, K.V., D. Sundar, S. Masilamani and A.R. Reddy, 2002. Variation in heat stress-induced antioxidant enzyme activities among three mulberry cultivars. Plant Growth Regul., 36: 175-180.
CrossRefDirect Link - Fang, W.C. and C.H. Kao, 2000. Enhanced peroxidase activity in rice leaves in response to excess iron, copper and zinc. Plant Sci., 158: 71-76.
CrossRefPubMedDirect Link - Frey, J.E., 1999. Genetic flexibility of plant chloroplasts. Nature, 398: 115-116.
CrossRefDirect Link - Gulen, H. and A. Eris, 2004. Effect of heat stress on peroxidase activity and total protein content in strawberry plants. Plant Sci., 166: 739-744.
CrossRefDirect Link - Heggie, L., M.A. Jansen, E.M. Burbridge, T.A. Kavanagh, R.N. Thorneley and P.J. Dix, 2005. Transgenic tobacco (Nicotiana tabacum L. cv. Samsun-NN) plants over-expressing a synthetic HRP-C gene are altered in growth, development and susceptibility to abiotic stress. Plant Physiol. Biochem., 43: 1067-1073.
CrossRefPubMedDirect Link - Jiang, Y. and B. Huang, 2001. Drought and heat stress injury to two cool-season turfgrasses in relation to antioxidant metabolism and lipid peroxidation. Crop Sci., 41: 436-442.
CrossRefDirect Link - Kawaoka, A., E. Matsunaga, S. Endo, S. Kondo, K. Yoshida, A. Shinmyo and H. Ebinuma, 2003. Ectopic expression of a horseradish peroxidase enhances growth rate and increases oxidative stress resistance in hybrid aspen. Plant Physiol., 132: 1177-1185.
CrossRefPubMedDirect Link - Kristensen, B.K., J. Brandt, K. Bojsen, H. Thordal-Christensen and K.B. Kerby et al., 1997. Expression of a defence-related intercellular barley peroxidase in transgenic tobacco. Plant Sci., 122: 173-182.
CrossRefDirect Link - Lagrimini, L.M., V. Gingas, F. Finger, S. Rothstein and T.T.Y. Liu, 1997. Characterization of antisense transformed plants deficient in the tobacco anionic peroxidase. Plant Physiol., 114: 1187-1196.
Direct Link - McCabe, M.S., J.B. Power, A.M. de Laat and M.R. Davey, 1997. Detection of single-copy genes in DNA from transgenic plants by nonradioactive Southern blot analysis. Mol. Biotechnol., 7: 79-84.
CrossRefDirect Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Passardi, F., D. Longet, C. Penel and C. Dunand, 2004. The class III peroxidase multigenic family in rice and its evolution in land plants. Phytochemistry, 65: 1879-1893.
CrossRefPubMedDirect Link - Quiroga, M., C. Guerrero, M.A. Botella, A. Barcelo and I. Amaya et al., 2000. A tomato peroxidase involved in the synthesis of lignin and suberin. Plant Physiol., 122: 1119-1128.
CrossRefPubMedDirect Link - Thordal-Christensen, H., J. Brandt, B.H. Cho, S.K. Rasmussen, P.L. Gregersen, V. Smedegaard-Petersen and D.B. Collinge, 1992. cDNA cloning and characterization of two barley peroxidase transcripts induced differentially by the powdery mildew fungus Erysiphe graminis. Physiol. Mol. Plant Pathol., 40: 395-409.
CrossRef - Vicuna, D., R.P. Malone and P.J. Dix, 2011. Increased tolerance to abiotic stresses in tobacco plants expressing a barley cell wall peroxidase. J. Plant Sci., 6: 1-13.
CrossRef