
Wei Li
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, Chengdu, Sichuan 611830, China
An-Jun Liu
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, Chengdu, Sichuan 611830, China
Yu-Zhen Sheng
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, Chengdu, Sichuan 611830, China
Guo-Yue Cheng
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, Chengdu, Sichuan 611830, China
Zi-En Pu
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, Chengdu, Sichuan 611830, China
Ya-Xi Liu
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, Chengdu, Sichuan 611830, China
Li Kong
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, Chengdu, Sichuan 611830, China
Journal of Plant Sciences
Year: 2013 | Volume: 8 | Issue: 1 | Page No.: 15-23







ABSTRACT
In order to exploit new genetic resources for the improving of starch quality of common wheat, the genetic diversity of null alleles of Granule-bound starch synthase I (waxy gene) was investigated by special PCR molecular markers in Triticum L. The results indicated that there was relative abundant genetic diversity of waxy alleles in all accessions. Accession AS2347, AS2356, AS2317 and AS2308 with null allele at Waxy-B1 locus and AS2310 and AS2335 with null alleles at Waxy-A1 and Waxy-B1, were observed in 81 landraces of Triticum turdigum L. from China. In 53 landraces of Triticum aestivum L. from Sichuan, China, eight accessions at Waxy-A1, Waxy-B1 and Waxy-D1 loci and accession AS1668 at Waxy-D1, were observed null alleles. In 29 Triticum macha, Accession PI361862 and PI572911 at three Waxy loci, PI572913 at Waxy-B1 and Waxy-D1, PI572910 at Waxy-A1 and Waxy-D1, PI 290507 at Waxy-B1 and PI572906 at Waxy-D1, respectively, were observed null alleles. Seven accessions with null alleles at Waxy-B1 locus was observed in 28 Triticum sphaerococcum. Specially, the accessions of two regions, Anyue in Sichuan, China and Georgia, had the high frequency of the mutations with null alleles of waxy gene. Landraces of Triticum aestivum L. with the high frequency of waxy wheat, could be considered as a unique genetic resource for improving of waxy wheat. These result suggested that the special molecular marker could be used reliably in evaluation of genetic resources and these mutations also could be directly used in the improving of common wheat.
PDF Abstract XML References Citation
Received: February 17, 2013;
Accepted: March 13, 2013;
Published: June 21, 2013
How to cite this article
Wei Li, An-Jun Liu, Yu-Zhen Sheng, Guo-Yue Cheng, Zi-En Pu, Ya-Xi Liu and Li Kong, 2013. Genetic Diversity of Null Alleles of Waxy Gene in Triticum L. Journal of Plant Sciences, 8: 15-23.
DOI: 10.3923/jps.2013.15.23
URL: https://scialert.net/abstract/?doi=jps.2013.15.23
DOI: 10.3923/jps.2013.15.23
URL: https://scialert.net/abstract/?doi=jps.2013.15.23
INTRODUCTION
Starch, accounts for 80% of wheat grain endosperm, is one of the most important components of cereal production (Hurkman et al., 2003). Starch is composed of amylose (20-30%) and amylopectin (70-80%) in cereal. The processing of starch synthesis includes three stages: the produce of ADPG, synthesis of amylose, synthesis of amylopectin (Martin and Smith, 1995). Granule-bound starch synthase I (GBSSI or WAXY protein) play a key role in synthesis of amylose in endosperm tissues of cereals (Shure et al., 1983). In wheat, GBSSI proteins were encoded by three genes: Wx-A1, Wx-B1 and Wx-D1, located on chromosome 7AS, 4AL and 7DS, respectively (Nakarnura, 1993). Either lack of GBSSI activity or absence of GBSSI protein at three loci will affect the quantitative and quality of amylose and further affect the quality of wheat product. Especially, the null allele of Wx-B1 has the most effect for amylase form of wheat, following Wx-D1 and Wx-A1 (Miura and Sugawara, 1996). There are eight types for null alleles of three Wx loci in wheat: the first type with all three alleles has 20-25% amylose content and the second to the seventh type with any one or two null alleles decreased the 1.7-5.0% amylose and the eighth type with three null alleles at three Wx loci lead to very less amylose content in wheat endosperm (Nakamura et al., 2002). Common wheat with partial null alleles at Wx loci, which lacks one or two WAXY proteins, have been identified widely, but the mutation with three null alleles at Wx loci in natural genetic resources was very less mentioned (Yamamori et al., 1998; Wang et al., 1999). Lacking all three WAXY proteins from three Wx loci could produce the waxy wheat (Nakamura et al., 1995). Through crossing the materials of Kanto 107 (null allele at Wx-A1 and Wx-B1 loci) and Baihuomiao (null allele at Wx-D1), the mutation has been generated with the entire null allele at three Waxy loci. Amylose content of this waxy wheat is 0.6-0.7% (Nakamura et al., 1993a, b, 1995). Otherwise, the starch characters and potential end use of waxy wheat have been reported (Lee et al., 2001; Bhattacharya et al., 2002).
The types of waxy proteins could be identified by 1D-SDS-PAGE (Zhao and Sharp, 1996) but the manipulation is complex and the endosperm of wheat is also damaged in the process of prepare sample. Molecular markers were also developed following know the sequence information of waxy gene in wheat (Ren et al., 2004). Using molecular marker could not only overcome the limitation of protein electrophoresis, but also detect easily in any stages of wheat development. Molecular assisted selection (MAS) of waxy gene can accelerate the improving of way wheat and wheat production. Three molecular marker of waxy gene had been applied in the breeding of waxy wheat in Austria (McLauchlan et al., 2001) and an improved method by one simple Polymerase Chain Reaction (PCR) to identify three Wx loci was also reported (Nakamura et al., 2002).
In the present study, the allele component of Waxy gene was investigated in landraces of Triticum aestivum L. (AABBDD, 2n = 42) from Sichuan, China, landraces of Triticum turgidum L. (AABB, 2n = 28) from China, Triticum macha Dekapr. et Menabde(AABBDD, 2n = 42) and Triticum sphaerococcum Perc. (AABBDD, 2n = 42) by special molecular marker. The results could help us to understanding the diversity of waxy gene in these genetic resources and exploiting new genetic materials to use in breeding of common wheat.
MATERIALS AND METHODS
Materials: A total of 81 accessions of T. turgidum L. landraces from China, 53 accession of T. aestivum L. landraces from Sichuan, China, 29 accessions of T. macha and 28 accessions of T. sphaerococcum, were used in this study (Table 1). The materials with AS were provided by the Germplasm Laboratory of Triticeae Research Institute, Sichuan Agriculture University and the other materials were kindly provided by Dr. Harold Bockelaman, USDA-ARS, National Small Grains Collection.
Methods: All seeds were germinated under the dark at 23 for 1 week, young leaves were harvested and crushed into powder with the aid of liquid nitrogen and the genomic DNA extracted by the CTAB method (Wang et al., 2008). A pair of primers, derived from the exon 4/6 region of the GBSSI gene (McLauchlan et al., 2001), was used as the special molecular markers. The forward primer: 5’-AAG AGC AAC TAC CAG T-3’ is located in exon 5 (position 1464-1481) and the reverse primer:
| Table 1: | Materials used in this study |
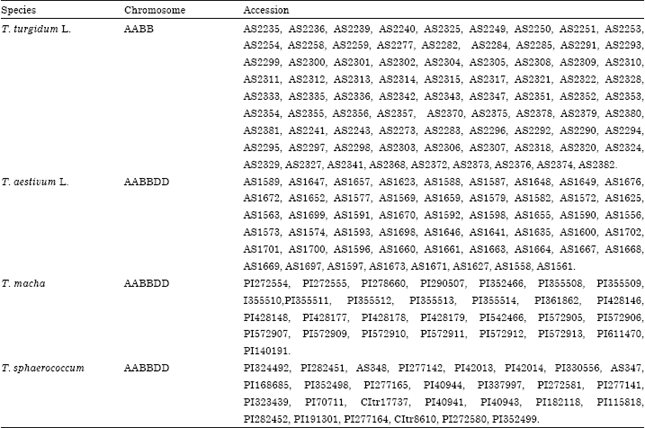 | |
5’-TCG TAC CC G TCG ATG AAG TCG A-3’ is located in exon 6(1508-1530) (Yan et al., 2000). PCR was performed in a 50 μL volume, containing 1.5 UTaq plus DNA polymerase, 100 ng of each template DNA, 5μL PCR buffer (supplied with Taq plus DNApolymerase), 1.5 mM MgCl2, 100 mM of each dNTP and 150 ng each primer. The reactions were conducted in a PTC-220 (MJ Research, USA) using the following program: 94°C for 3 min, followed by 12 cycles at 94°C for 1 min, at 64°C for 1 min and at 72°C for 30 sec and followed by 34 cycles at 94°C 1 min, 58°C 1 min and 72°C 30 sec and a final extension step of 72°C for 5 min. The PCR products were separated on 3% agarose gel.
RESULTS
Allele variation of Waxy gene in landraces of T. turgidum L.: Two bands could be amplified at Wx-A1 and Wx-B1 loci in the landraces of T. turgidum L. from China (Fig. 1a). The band with bigger size came from Wx-A1 locus and the small one from Wx-B1 locus. The different size of PCR product resulted from the different length between the fifth exon and the sixth exon of waxy gene. Both two bands were obtained in 75 accessions of all 81 accessions of tetraploid landraces. No band was obtained in accession AS2310 (Nanmai, Renshou) and AS2335 (Dabaimai, Anxi), indicated that there were null alleles at Wx-A1 and Wx-B1 loci in these two accessions. Four accessions (4.9%), AS2347 (Fushoumai, Jianhu), AS2356 (Fushoumai, Qishan), AS2317 (Zaozhuo, Batang) and AS2308 (Wugongnanmai, Suining), were identified as the mutation with null allele at Wx-B1 locus. These results suggested that the frequency of mutations with null alleles at Wx loci was very lower in landraces of T. turgidum L. from China.
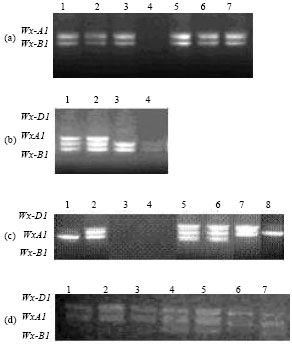 | |
| Fig. 1(a-d): | PCR assays for detection of null alleles of the waxy genes in Triticum L. (a) Landraces of T. turgidum L., 1. AS2236, 2. AS2239, 3. AS2240, 4. AS2310, 5.AS2311, 6. AS2312, 7. AS2313. (b) Landraces of T. aestivum L., 1. AS1589, 2. AS1647, 3. AS1668, 4. AS1591. (c) T. macha, 1. PI572910, 2. PI572906, 3. PI361862, 4. PI572911, 5. PI272554, 6. PI272555, 7. PI290507, 8. PI572913. (d) T. sphaerococcum, 1. PI324492, 2. PI282451, 3. AS348, 4. PI277142, 5. PI42013, 6. PI42014, 7. PI330556 |
Allele variation of Waxy gene in landraces of T. aestivum L.: Three bands were identified at Wx-A1, Wx-B1 and Wx-D1 loci in T. aestivum L. from Sichuan, China (Fig. 1b). The band with the biggest size was amplified from Wx-D1 locus and the one with the second bigger size from Wx-A1 locus and the least band from Wx-A1 locus. No bands were amplified from eight accessions (15%) in 53 landraces of common wheat. These mutations with null alleles at three Wx loci included AS1591 (Hongxiaomai, Anyue), AS1670 (Shihonghuamai, Pengxian), AS1592 (Nverhong, Anyue), AS1598 (Dahongpao, Zizhong), AS1655 (Baixiaomai, Dazhu), AS1590 (Dongmai, Anyue), AS1556 (Tuomai, Bazhong) and AS1573 (Liulengmai, Changning). Lacking the band from Wx-D1 locus was only observed in one accession, AS1668 (Youmangmai, Xinjing). Whole three bands were amplified from 43 accessions of landraces of common wheat. Three accessions from Anyue, Sichuan, China, were the mutations with null alleles at three Wx loci. The results suggested that there was abundant allele diversity of Waxy gene in landraces of common wheat from Sichuan, China.
Allele variation of Waxy gene in T. macha: Three bands could be obtained from 23 accessions of T. macha (Fig. 1c). No band was amplified in two accessions, PI361862 (from Danmark) and PI572911 (from Georgia), indicated that they belonged to the mutations with null alleles at three Wx loci. Only one band was amplified in two accessions, PI572913 (from Georgia), which showed null alleles at Wx-B1 and Wx-D1 loci and PI572910 (from Georgia), which showed null alleles at Wx-A1 and Wx-D1 loci. Only one accession, PI290507 (from Hungary), showed the null allele at Wx-B1 locus and one accession, PI572906 from Georgia, had the null allele at Wx-D1 locus. Four out of six mutations with at least one null allele at three Wx loci came from Georgia, suggested that it was very important region for genetic resources of Waxy gene.
Allele variation of Waxy gene in T. sphaerococcum: Three bands were amplified in 21 accessions of T. sphaerococcum (Fig. 1d). Null allele at Wx-B1 locus was identified in five accession from India, which including PI324492, PI282451, PI42014, PI352498 and PI282452 and two from Pakistan, which including PI40941 and PI40943. The result suggested that there was relative simple genetic diversity of allele of Waxy gene in T. sphaerococcum.
DISCUSSION
Very less information about natural genetic resources of common wheat with null alleles at three Wx loci was reported (Yamamori et al., 1998; Wang et al., 1999). The waxy wheat with three null alleles at waxy gene locus was produced by crossing between mutations with partial null alleles at different Wx loci (Nakamura et al., 1993a; b 1995). In recent years, waxy wheat is becoming more and more interesting for wheat breeders and industry, because of its unique quality characters (Lee et al., 2001; Bhattacharya et al., 2002). In addition, modification of cereal starch by various methods could provide different production suitable for food and various industrial applications (Kaur et al., 2012). Waxy gene plays an important role in synthesis of cereal starch and its activity directly affects the quantity and quality of starch. Thus, exploiting the new allele of waxy gene in genetic resources of common wheat is very important work for the improving of wheat and modification of starch. In the present study, the allele variation was investigated in the four important genetic resources of common wheat, landraces of T. turgidum L., landraces of T. aestivum L., T. macha, T. sphaerococcum. Landraces of T. turgidum L., which was grown widely in China before the 1950’s, showed the unique performance about the gliadin, HMW-glutenin, protein content and microsatellite (Li et al., 2006a, b). Landraces of T. aestivum L. from Sichuan, China, also known as “Sichuan White Wheat”, were widely planted and conserved by the peasants in the province ago (Liu et al., 2005). This population is considered to be a unique resource for genetics and breeding, because of broad hybrid compatibility with rye (Luo et al., 1992; Liu et al., 2003). T. macha and T. sphaerococcum were also displayed abundant genetic diversity and contain gene resource for quality and disease resist of wheat (Cao et al., 1998; Chen et al., 2012).
In present study, abundant genetic diversity of alleles of Waxy gene was observed in Triticum L. species. In tetraploid, T. turgidum L., two accessions with null alleles at both Wx-A1 and Wx-B1 loci, four accessions with null allele at Wx-B1 locus, suggested that there was relative lower diversity of allele of waxy gene in this landraces. In hexaploid, T. sphaerococcum also showed the similar performance of alleles of waxy gene, as only null allele at Wx-B1 locus was observed in seven accessions. Landraces of T. aestivum L. from Sichuan, China, showed the highest percentage (15%) of null alleles at three Wx loci. Including the mutations with null alleles at three, two and one Wx loci, were detected in T. macha, suggested that this hexaploid species had abundant genetic diversity of alleles of waxy gene. Actually, this species with high genetic diversity was also observed by RAPD (Cao et al., 1998). In addition, the mutations with different null allele at Wx loci, which were identified now, could be used in the improving of common wheat. The next steps for these mutations with null alleles of waxy gene should be focus on sequence cloning and gene expression to understanding the molecular mechanism of these null alleles.
Waxy mutations with null alleles occur spontaneously in cereals (Eriksson, 1970). Null alleles at the Wx-A1 locus have been found in samples from Japan, Korea and Turkey, while null alleles at the Wx-B1 locus are very common in the materials from Australia and India (Yamamori et al., 1994, Yamamori and Endo, 1996) and Italy (Boggini et al., 2001). Null alleles at the Wx-D1 locus were more rarely and identified only in Baihuo (Yamamori et al., 1994) and one Italian landrace (Boggini et al., 2001). Wang et al. (1999) also found one accessions with null allele at Wx-D1, six accessions with null alleles at Wx-B1, in 900 landraces and cultivars. These results suggested that the mutations with null alleles at Wx loci had the very lower frequency in genetic resources. In our results, all the accessions still showed the relative lower percentage of null alleles of waxy gene. In a total of 191 accessions, only two hexaploid accessions showed null alleles at both two Wx loci and 12 accessions showed null alleles at Wx-B1 locus and two accessions showed null alleles at Wx-D1 locus. But the mutations with null alleles at three Wx loci had higher percentage, especially in landraces of T. aestivum L. from Sichuan, China (15%), suggested that this landraces population is unique genetic resource for improving of waxy wheat. The reason that the landraces had high frequency of null alleles could be related with the special dietetic habit of people living in Sichuan.
The usual method to breeding waxy wheat is to crossing these materials with different null alleles of Waxy gene (Yamamori et al., 1995; Urbano et al., 2002). Now 10 accessions with null alleles at three Wx loci were identified from Triticum L. genetic resources. All the mutations could be directly used in breeding of common wheat and save the time of selection. Furthermore, the variation of Waxy gene was only investigated at DNA level. The performance of amylose and starch and the quality in these mutation materials should be further investigated. The influence of the variation of Waxy gene loci for the amylose and the relating quality characters also should be focused in the future.
Identifying null allele types of waxy gene of the materials is very important for the breeding of waxy wheat. The main methods for waxy gene expressing included measuring the amylose content, swelling power, RVA pasting characteristics, iodine staining and waxy protein electrophoresis (Nakamura et al., 1993a, b ;Crosbie, 1991; McCormick et al., 1994; Sulaiman and Morrison, 1990). Zhao and Sharp (1996) improved the 1D-SDS-PAGE method and can easy identify three WAXY protein and suggested that this method could be used in wheat breeding. Wang et al. (1999, 2000) further improved SDS-PAGE method and could identify four WAXY subunits (Wx-A1, Wx-E1, Wx-D1 and Wx-B1). Pan et al. (2000) also used the 2D-SDS-PAGE to distinguish WAXY protein. But SDS-PAGE method is difficult to apply widely in breeding of wheat, because the process of preparative of samples and manipulation process is very complex and lower effective. The molecular marker method based on PCR was applied widely after the cDNA sequence of Waxy gene reported. Shariflou and Sharp (1999) designed SSR primers to identify the Wx-D1 locus and Briney et al. (1998) designed STS-PCR primers to identify the Wx-B1 locus. Nakamura et al. (2002) confirm that the PCR markers could identify the waxy wheat with different null alleles at Wx loci and McLauchlan et al. (2001) reported that the special PCR markers, which could identify null alleles of waxy gene, were applied in Australian wheat breeding program. The present study also further confirmed that these molecular markers also could be used reliably in evaluation of genetic resources of Triticum L.
CONCLUSION
In this study, the mutations with null alleles at Wx loci were identified in landraces of T. turgidum L. from China, landraces of T. aestivum L. from Sichuan, China, T. macha and T. sphaerococcum, by one pair of specific primer. In all 191 accessions, only two hexaploid accessions showed null alleles at both two Wx loci and 12 accessions showed null alleles at Wx-B1 locus and two accessions showed null alleles at Wx-D1 locus. Interestingly, 10 hexaploid accessions with null alleles at three Wx loci and two tetraploid accessions with null alleles at two Wx loci, were observed. The mutations with null alleles of Waxy gene were easily obtained in the accessions from Anyue, Sichuan, China and from Georgia. Landraces of T. aestivum L. from Sichuan, China, is unique genetic resource for improving of waxy wheat, because of the high frequency of waxy wheat. The reason could be related with the special dietetic habit of people living in this region. Further research could help to deeply understand the performance of these mutations about sequence variation, gene expression and quality. These result suggested that the special molecular markers could be used reliably in evaluation of genetic resources and these mutations also could be used directly in the improving of common wheat.
ACKNOWLEDGMENTS
This study was supported by the National Basic Research Program of China (973 Program and 2011CB100100) and the National Basic Research Special Program of China (Grant No. 2010CB134402).
REFERENCES
- Bhattacharya, M., S.V. Erazo-Castrejon, D.C. Doehlert and M.S. McMullen, 2002. Staling of bread as affected by waxy wheat flour blends. Cereal Chem., 79: 178-182.
CrossRefDirect Link - Boggini, G., M. Cattaneo, C. Paganoni and P. Vaccino, 2001. Genetic variation for waxy proteins and starch properties in Italian wheat germplasm. Euphytica, 119: 113-116.
Direct Link - Briney, A., R. Wilson, R.H. Potter, I. Barclay, G. Crosbie, R. Appels and M.G.K. Jones, 1998. A PCR-based marker for selection of starch and potential noodle quality in wheat. Mol. Breeding, 4: 427-433.
CrossRefDirect Link - Cao, W., P. Hucl, G. Scoles and R.N. Chibbar, 1998. Genetic diversity within spelta and macha wheats based on RAPD analysis. Euphytica, 104: 181-189.
CrossRefDirect Link - Chen, S.S., G.Y. Chen, H. Chen, Y.M. Wei and W. Li et al., 2012. Mapping stripe rust resistance gene YrSph derived from Triticum sphaerococcum Perc. with SSR SRAP and TRAP markers. Euphytica, 185: 19-26.
Direct Link - Crosbie, G.B., 1991. The relationship between starch swelling properties, paste viscosity and boiled noodle quality in wheat flours. J. Cereal Sci., 13: 145-150.
CrossRefDirect Link - Kaur, B., F. Ariffin, R. Bhat and A.A. Karim, 2012. Progress in starch modification in the last decade. Food Hydrocolloids, 26: 398-404.
CrossRefDirect Link - Hurkman, W.J., K.F. McCue, S.B. Altenbach, A. Korn and C.K. Tanaka et al., 2003. Effect of temperature on expression of genes encoding enzymes for starch biosynthesis in developing wheat endosperm. Plant Sci., 164: 873-881.
CrossRefDirect Link - Lee, M.R., B.G. Swanson and B.K. Baik, 2001. Influence of amylose content on properties of wheat starch and breadmaking quality of starch and gluten blends. Cereal Chem., 78: 701-706.
CrossRefDirect Link - Li, W., Y.M. Wei, Z.H. Yan and Y.L. Zheng, 2006. Evaluation of grain protein content in Triticum turgidum ssp. turgidum landraces from Southwest China. J. Plant Sci., 1: 378-382.
CrossRefDirect Link - Liu, Y.J., Y.B. Huang and T.Z. Rong, 2005. Comparative analysis of genetic diversity in landraces of waxy maize from Yunnan and Guizhou using SSR marker. Agric. Sci. China, 4: 648-653.
Direct Link - Liu, D., Y. Zheng and X. Lan, 2003. Utilization of wheat landrace Chinese spring in breeding. Sci. Agric. Sinica, 36: 1383-1389.
Direct Link - Luo, M.C., C. Yen and J.L. Yang, 1992. Crossability percentages of bread wheat landraces from Sichuan province China with rye. Euphytica, 67: 1-8.
CrossRef - McLauchlan, A., F.C. Ogbonnaya, B. Hollingsworth, M. Carter and K.R. Gale et al., 2001. Development of robust PCR-based DNA markers for each homoeoallele of granule-bound starch synthase and their application in wheat breeding programs. Aust. J. Agric. Res., 52: 1409-1416.
Direct Link - McCormick, K.M., J.F. Panozzo and S.H. Hong, 1994. A swelling power test for selecting potential noodle quality wheat. Australia J. Agric. Res., 42: 317-323.
Direct Link - Miura, H. and A. Sugawara, 1996. Dosage effects of the three Wx genes on amylose synthesis in wheat endosperm. Theor. Applied Genet., 93: 1066-1070.
CrossRefDirect Link - Nakamura, T., M. Yamamori, H. Hirano and S. Hidaka, 1993. Decrease of waxy (Wx) protein in two common wheat cultivars with low amylose content. Plant Breeding, 111: 99-105.
CrossRefDirect Link - Nakamura, T., M. Yamamori, H. Hirano and S. Hidaka, 1993. Identification of three wX proteins in wheat (Triticum aestivum L.). Biochem. Genet, 31: 75-86.
CrossRef - Nakamura, T., M. Yamamori, H. Hirano, S. Hidaka and T. Nagamine, 1995. Production of waxy (amylose-free) wheats. Mol. Gen. Genet., 248: 253-259.
CrossRefPubMedDirect Link - Nakamura, T., P. Vrinten, M. Saito and M. Konda, 2002. Rapid classification of partial waxy wheats using PCR-based markers. Genome, 45: 1150-1156.
PubMedDirect Link - Pan, Z.F., G.B. Deng, T. Wang and M.Q. Yu, 2000. An improved 1D-SDS-PAGE method for identification of wheat waxy protein. Chinese J. Applied Environ. Biol., 6: 487-489.
Direct Link - Ren, L.J., S.B. Cai, H.X. Ma, X. Zhang and M.P. Zhou, 2004. Rapid classification of partial waxy and waxy wheats using a SSR marker. J. Triticeae Crops, 24: 9-12.
Direct Link - Shariflou, M.R. and P.J. Sharp, 1999. A polymorphic microsatellite in the 3'end of waxy genes of wheat, Triticum aestivum. Plant Breeding, 118: 275-277.
CrossRef - Shure, M., S. Wessle and N. Fedoroff, 1983. Molecular identification and isolation of the waxy locus in maize. Cell, 35: 225-233.
CrossRefDirect Link - Sulaiman, B.D. and W.R. Morrison, 1990. Proteins associated with the surface of wheat starch granules purified by centrifuging through caesium chloride. J. Cereal Sci., 12: 53-61.
CrossRef - Urbano, M., B. Margiotta, G. Colaprico and D. Lafiandra, 2002. Waxy proteins in diploid, tetraploid and hexaploid wheat. Plant Breeding, 121: 465-469.
CrossRefDirect Link - Wang, J.R., Y.M. Wei, X.Y. Long, Z.H. Yan, E. Nevo, B.R. Baum and Y.L. Zheng, 2008. Molecular evolution of dimeric α-amylase inhibitor genes in wild emmer wheat and its ecological association. BMC Evol. Biol., Vol. 8.
CrossRefDirect Link - Wang, Z.N., B.H. Guo, Y.M. Zhang, Z.Y. Wen, H.J. Li and Y.S. Shi, 1999. Discovery and analysis of wheat cultivar (T. aestivum) with Wx genes. Acta Agriculturae Boreali-Sinica, 14: 5-9.
Direct Link - Yamamori, M., T. Nakamura, T.R. Endo and T. Nagamine, 1994. Waxy protein deficiency and chromosomal location of coding genes in common wheat. Theor. Applied Genet., 89: 179-184.
Direct Link - Yamamori, M., T. Nakamura and T. Nagamine, 1995. Polymorphism of two waxy proteins in the emmer group of tetraploid wheat, Triticum dicoccoides, T. dicoccum and T. durum. Plant Breeding, 114: 215-218.
CrossRef - Yamamori, M. and T.R. Endo, 1996. Variation of starch granule proteins and chromosome mapping of their coding genes in common wheat. Theor. Applied Genet., 93: 275-281.
Direct Link - Yan, L.L., M. Bhave, R. Fairclough, C. Konik, S. Rahman and R. Appels, 2000. The genes encoding granule-bound starch synthases at the waxy loci of the A, B and D progenitors of common wheat. Genome, 43: 264-272.
CrossRefDirect Link - Zhao, X.C. and P.J. Sharp, 1996. An improved 1-D SDS-PAGE method for the identification of three bread wheat waxy proteins. J. Cereal Sci., 23: 191-193.
CrossRef