
Ze-Hou Liu
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, Sichuan 611130, China
Yong Zhou
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, Sichuan 611130, China
Jia Li
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, Sichuan 611130, China
Yu-Jiao Liu
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, Sichuan 611130, China
Ya-Xi Liu
Triticeae Research Institute, Sichuan Agricultural University, Wenjiang, Sichuan 611130, China
Journal of Plant Sciences
Year: 2013 | Volume: 8 | Issue: 1 | Page No.: 1-14







ABSTRACT
Alpha-amylase enzyme is the important member in physiological metabolism of high plant, especially during seed germination stage. The current study identified 19 amy1 genes from 19 hexaploid wheat (Triticum aestivum L.) accessions. The nucleotide and deduced amino acid sequences of amy1 genes were analyzed in detail, respectively. It showed that two main variation types were characterized due to the base substitution and/or indel mutations in genomic nucleotide sequences and the exon and intron domains were presented different variation degree. This study identified 8 clustered haplotypes from hexaploid wheat accessions through characterization of exon domains. The haplotype 8 was the major variation types and the others were low relative frequency. The haplotype 7 was a special kind of haplotypes among amy1 genes in wheat accession PI243793 because of the variation of amino acid sequences at 155-161 sites. These results would contribute to the understanding in functional aspects and efficient utilization of amy1 genes in hexaploid wheat cultivars.
PDF Abstract XML References Citation
Received: February 23, 2013;
Accepted: March 14, 2013;
Published: June 21, 2013
How to cite this article
Ze-Hou Liu, Yong Zhou, Jia Li, Yu-Jiao Liu and Ya-Xi Liu, 2013. Molecular Characterization of the Amy1 Gene in Hexaploid Wheat. Journal of Plant Sciences, 8: 1-14.
DOI: 10.3923/jps.2013.1.14
URL: https://scialert.net/abstract/?doi=jps.2013.1.14
DOI: 10.3923/jps.2013.1.14
URL: https://scialert.net/abstract/?doi=jps.2013.1.14
INTRODUCTION
In hexaploid wheat, Triticum aestivum (2n = 6x = 42), alpha-amylase enzyme has received much attention in functional and evolutionary studies and also been regarded as the important enzyme for both agricultural (germination of cereal grains) and industrial (the brewing industry) interests. The genotypes analysis of amy gene also attracted more scientists to study including chromosome location, functional analysis, enzyme activity and allelic variation. Gale et al. (1983) also developed variety of genetic population to study the amy genotype. Alpha-amylase contributes to the grain germination in wheat but it regulates the germination with different tissue of seed (embryo and aleurone) during developing grain (Garcia-Maya et al., 1990; Appleford and Lenton, 1997).
Alpha-amylase has been participated in many physiological processes and metabolic activities (Laurie et al., 2003). The alpha-amylase activity in the grain was used as an indicator for pre-harvest sprouting (Li et al., 2003; Lin et al., 2008). During the germination of wheat grains, alpha-amylase encoded by amy gene play a key role in the mobilization of the energy reserves stored in insoluble starch granules (Sugimoto et al., 1998). Furthermore, the transcription of alpha-amylase was regulated by hormone including gibberellins and abscisic acid (Jacobsen and Beach, 1985; Higgins et al., 1976). Alpha-amylase activity is also negative association with Hagberg Falling Number (HFN), an important test of bread-making quality in wheat. Kindred et al. (2005) also reported that high alpha-amylase activity could result in the production of sticky doughs which would be difficult to industrial process.
Many alpha-amylase genes have been cloned from various cereals (Mitsui and Itoh, 1997). Great progress has been made in the analysis of alpha-amylase multigene families in cereal, including characterization of protein structure and secretion of the enzyme. Alpha-amylase genes of wheat are expressed at least twice during the life cycle of the plant. They are expressed at a high level during germination in the aleurone cells when the controlling influence at the cellular level is the plant hormone gibberellic acid (Baulcombe et al., 1987).
In current study, 19 amy1 genes from hexaploid wheat (Triticum aestivum L.) accessions were cloned and identified. The present study was conducted to isolate and characterize the amy1 genes from hexaploid wheat accessions and to analysis the allelic variation of these genes in the wheat accessions derived from different regions.
MATERIALS AND METHODS
Plant materials and growth conditions: In total, 19 hexaploid wheat accessions derived from various regions including Canada, Eritrea, Mexico, UK, Iraq, China, UK, Ukraine, Australia, Iran, Peru, Ethiopia, US and Tajikistan (Table 1) were used to characterize amy1 genes. These experimental wheat materials were grown under a randomized complete block design with two replicates at the Experimental Station of Sichuan Agricultural University at Chengdu-Wenjiang, Sichuan, during the 2012 crop seasons.
DNA extraction and PCR amplification: Genomic DNA was extracted from the seeding leaves of each accession about 2 weeks old using a modified CTAB (Hexadecyltrimethylammonium Bromide) extraction method described by (Murray and Thompson,1980). The specific primers (Forward: 5’-CAACCGGAGAAGAAGAGTGAC-3’, Reverse: 5’-ATGGTGGATCAGTGGAG ACTT-3’) were designed by bioinformatics software DNAMAN 5.0 (LynnonBioSoft, USA) and Primer Premier 5.0 (Premier Biosoft International, Palo Alto, CA) on the basis of the wheat tests sequences derived from NCBI (http://www.ncbi.nlm.nih.gov/) and selected for amplification the target Taamy1 gene.
| Table 1: | Plant materials used in this study |
 | |
These TaESTs sequences were aligned by homology gene, Hvamy1 (Genbank No.: FN179389).
Statistical analysis: The multiple alignment of sequences was carried out by DNAMAN 5.0 (LynnonBiosoft corporation, US), respectively. The phylogenetic tree based on the phylogenetic analysis was performed by Mega 4.0 (Tamura et al., 2007).
RESULTS
Variation analysis: Currently, 19 nucleotide sequences of amy1 gene sequences were obtained from the hexaploid wheat accessions basis on the homology-cloning according to Hvamy1 gene (Genbank No.: FN179389). The similarity of nucleotide sequences of exon domains between amy1 genes in wheat accessions and Hvamy1 gene was 97.18% and the similarity of deduced amino acid residue sequences among them was 91.30%. Amy1 gene derived from hexaploid wheat contained 4 exons and 3 introns (Fig. 1). In current study, the genomic sequences of amy1 genes obtained from hexaploid wheat accessions contained 1691 to 1715 nucleotides (exon domains, 1314 to 1339 nucleotides and intron domains, 376 to 402 nucleotides). The nucleotide sequences of nineteen amy1 genes shared a high sequence identity (94.07%) and the coding and noncoding domains sequences shared 96.71 and 91.69%, respectively. Furthermore, these nucleotides in entire coding domains might encode 437 to 440 amino acid residues. There were 51 variation positions in entire exon domains and it could be divided into two large genotypes (Table 2). Among those variation positions, it presented 42 Single Nucleotide Polymorphisms (SNPs) and 9 polynucleotide variation sites (Table 2), including 4 indel sites. These SNPs of coding sequences resulted in diverse variation of deduced amino acids sequences. Most of them might lead to the structural variation of alpha-amylase I enzyme protein and contribute to different function due to the different binding sites.
Haplotype diversity: Currently, it indicated that seven haplotypes were evaluated in accordance with SNP and indel variation analysis. The haplotype 8 was presented most frequently among the genotypes and it was involved in 9 genes simples (PI70713, PI94390, PI94460, PI94504, PI113933, PI124820, PI648908, PI651298 and PI654219). The second frequent haplotype was haplotype 1, which were amy1 genes from four genotypes including CItr6618, CItr8195, CItr14811 and CItr17599. The other 6 cluster of haplotypes was only had one genotype respectively. It demonstrated that amy1 genes from hexaploid wheat accessions presented high polymorphism. Referring to amy1 gene in hexaploid wheat, the diverse variation of coding domains could be lead to variation of amino acid sequences, even result in the change of base protein structure. More new alleles of amy1 genes could be frequently observed in hexaploid wheat.
Deduced amino acid analysis: The deduced amino acid sequences of alpha-amylase presented certain polymorphism. Herein, the coding domains of 18 amy1 gens could encode the normal amino acid reduces and develop the normal alpha-amylase proteins and only one from PI477864 could not conduct the transcript process because of the abnormal present of termination codon in coding domain of amy1 gene. The changes in the deduced amino acid residues at 35 positions could be presented high polymorphism (Table 3), among which four deduced amino acid sequences showed the high homology with the same variation sites differing in the others.
| Table 2: | Distribution patterns of SNPs and indels of haplotypes in coding domain of nineteen amy1 genes from hexaploid wheat accessions |
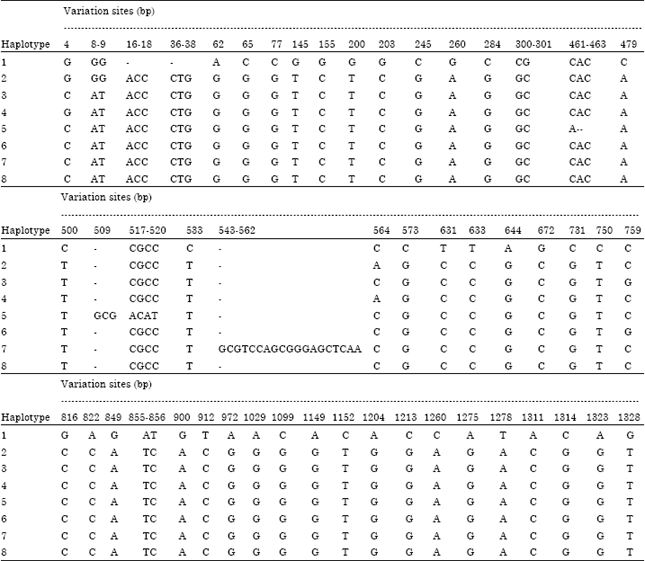 | |
Interestingly, PI243793 indicated that the only variation pattern from 153 to 176 amino acids reduces (Fig. 2). Possibility, it would contribute to the special mechanism function of alpha-amylase though the whole plant lifespan.
Phylogenetic analysis: The phylogenetic relationships among the amy1 genes in these hexaploid wheat accessions derived from different countries around the world was evaluated and indicated by phylogenetic distances in neighbor-joining trees (Fig. 3) developed by MEGA 4.0. The clustering results indicated that the amy1 gene was obviously divided into two groups. Four genes from CItr6618, CItr8195, CItr14811 and CItr17599 were clustered into one group and the other genes were included in the second group (Fig. 3). Furthermore, amy1 gene from wheat accession PI243793 with special variation of amino acid sequences had genetic relationship from the others in the second clustered group.
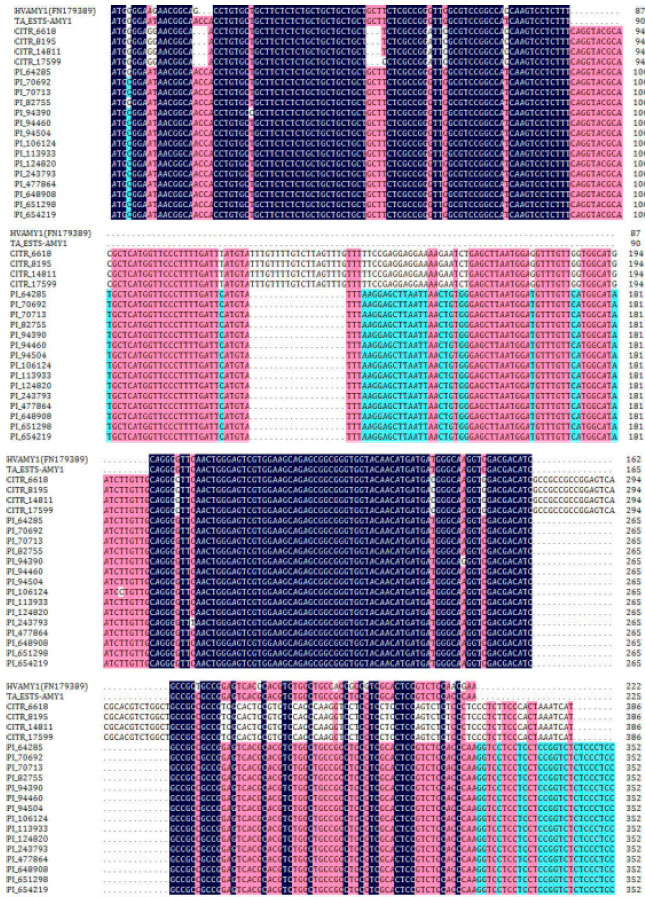   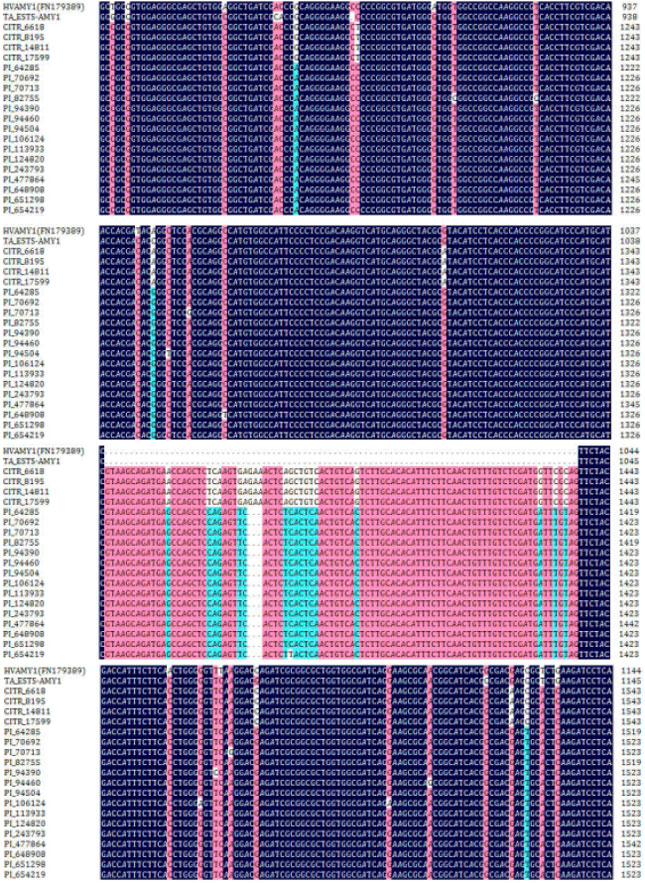  | |
| Fig. 1: | The chart of alignment genomic sequence of amy1 genes from hexaploid wheat accessions |
DISCUSSION
Alpha-amylases encoded by amy genes play an essential role in the germination and has been regarded as the most important enzymes and the trigger for regulating germination in wheat grain (Huang et al., 1993; Karrer and Rodriguez, 1992). Variety of alpha-amylase genes including amy1, amy2 and amy3 (Baulcombe et al., 1987) and Late Maturity Alpha-amylase (LMA) gene (Mrva and Mares, 1996, 2001; Mrva et al., 2009) have also been reported in previous published literatures. The three types of amy1 genes shared a common evolutionary ancestor (Baulcombe et al., 1987). Similarly, the SNPs of amy1 genes in barley were analyzed and indicated many variation sites were associated with malting quality index (MQI) (Matthies et al., 2009). The allelic variation of amy1 genes from 19 hexaploid wheat accessions was identified in this study. Integrated genomic sequence of Taamy1 gene was not reported in NCBI (http://www.ncbi.nlm.nih.gov/). This study provided the reference genomic sequence of Taamy1 gene for further research. The Taamy1 gene contained 1691 to 1715 nucleotides and 4 exons and 3 introns and the noncoding domain shared more variation than coding domain. Nevertheless, the structure of Taamy1 in this study was different from Hvamy1gene which just contained two introns (Knox et al., 1987).
In current study, by the alignment and phylogenetic analysis, the amy1 gene sequences from 19 hexaploid wheat accessions could be distinguished into two different types (Fig. 3) and had relative great variation in nucleotides and amino acids sequences (Fig. 1, 2). Apparently, many studies just compared the difference of amy multigene family (Lazarus et al., 1985; Baulcombe et al., 1987). Little was known about the characterization of amy1 gene.
 | |
| Fig. 2: | The chart of alignment of amino acid sequences of amy1 genes from hexaploid wheat accessions |
| Table 3: | Variation of amino acids in amy1 genes from hexaploid wheat accessions |
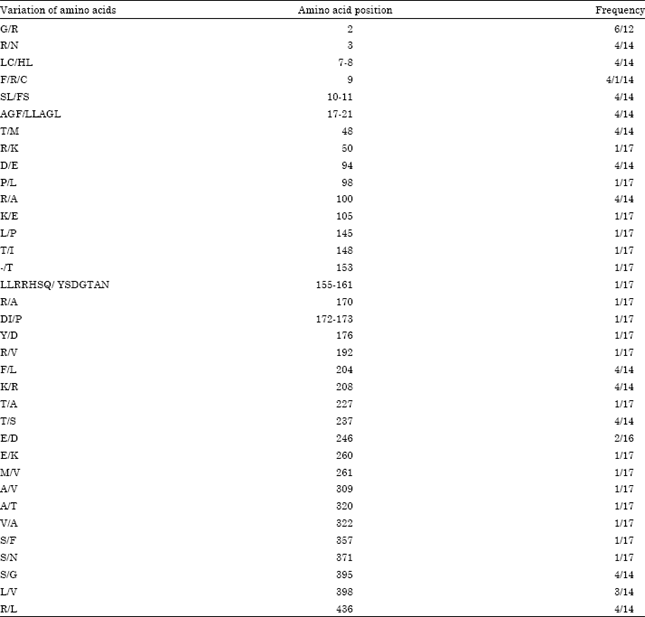 | |
Possibility, this study provided a new insight that was two copies of amy1 gene from hexaploid wheat. Additionally, the sequence alignment and haplotypes analysis revealed 8 haplotypes in 19 hexaploid wheat accessions from the diverse countries. It showed that amy1 gene has more variation and haplotype 8 was found to be the major type. Furthermore, the haplotype 7 would be used to develop the potential marker for different function in plant metabolize.
Variation of amy1 gene influences the grain germination in wheat greatly, especially in PHS (Lin et al., 2008; Li et al., 2004). And the activation of alpha-amylases was reported to associated with the grain quality and adaption of cereal crops by high temperature (Hakata et al., 2012). It would be important for identification of germplasm resources from current hexaploid wheat accessions via characterization of amy1 genes. Two clustered types might perform different molecular function in plant physiological development.
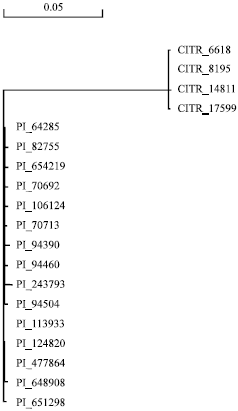 | |
| Fig. 3: | Phylogenetic tree of amy1 genes |
Moreover, the special variation of amino acid sequences was at 155 to 161 sites (Table 3, Fig. 2) and the relevant nucleotide sequence of amy1 gene would be the potential marker for new function developing. A further understanding for allelic variation of amy1 gene in hexaploid wheat would benefit our breeder to improve our wheat cultivars during breeding process, particularly in resistance to PHS. All discovered SNPs could be converted into high-throughput markers for pyrosequencing and might be used for marker assisted selection.
CONCLUSION
In current study, we have isolated the amy1 genes in 19 hexaploid wheat accessions and identified natural variation for nucleotides and amino acids sequences of amy1 genes. amy1 gene in hexaploid wheat accessions presented high variation degree. It demonstrated that two main variation types and eight clustered haplotypes appeared in hexaploid wheat accessions. Furthermore, it was interesting that haplotype 7 as a special kind of haplotypes among amy1 genes was just cloned in wheat accession PI243793 due to the unique variation sites of nucleotides and amino acids sequences, which might be severed as a special material for future utilization of amy1 genes in hexaploid wheat cultivars.
ACKNOWLEDGMENTS
This study was supported by Specialized Research Fund for the Doctoral Program of Higher Education (20115103120008), Science Foundation for Youths of the Education Department of Sichuan Province (10ZB039) and National Natural Science Foundation of China (31171555; 31230053).
REFERENCES
- Appleford, N.E.J. and J.R. Lenton, 1997. Hormonal regulation of α-amylase gene expression in germinating wheat (Triticum aestivum) grains. Physiol. Plantarum, 100: 534-542.
CrossRefDirect Link - Garcia-Maya, M., J.M. Chapman and M. Black, 1990. Regulation of α-amylase formation and gene expression in the developing wheat embryo. Planta, 181: 296-303.
Direct Link - Gale, M.D., C.N. Law, A.J. Chojecki and P.A. Kempton, 1983. Genetic control of α-Amylase production in wheat. Theor. Applied Genet., 64: 309-316.
CrossRef - Hakata, M., M. Kuroda, T. Miyashita, T. Yamaguchi and M. Kojima et al., 2012. Suppression of α-amylase genes improves quality of rice grain ripened under high temperature. Plant Biotech. J., 10: 1110-1117.
CrossRefPubMedDirect Link - Higgins, T.J.V., J.A. Zwar and J.V. Jacobsen, 1976. Gibberellic acid enhances the level of translatable mRNA for α-amylase in barley aleurone layers. Nature, 260: 166-169.
CrossRefDirect Link - Huang, N., J. Chandler, B.R. Thomas, N. Koizumi and R.L. Rodriguez, 1993. Metabolic regulation of α-amylase gene expression in transgenic cell cultures of rice (Oryza sativa L.). Plant Mol. Biol., 23: 737-747.
CrossRefPubMedDirect Link - Jacobsen, J.V. and L.R. Beach, 1985. Control of transcription of α-amylase and rRNA genes in barley aleurone protoplasts by gibberellins and abscisic acid. Nature, 316: 275-277.
CrossRefDirect Link - Karrer, E.E. and R.L. Rodriguez, 1992. Metabolic regulation of rice α-amylase and sucrose synthase genes in planta. Plant J., 2: 517-523.
CrossRefDirect Link - Kindred, D.R., M.J. Gooding and R.H. Ellis, 2005. Nitrogen fertilizer and seed rate effects on Hagberg falling number of hybrid wheats and their parents are associated with α-amylase activity, grain cavity size and dormancy. J. Sci. Food Agric., 85: 727-742.
CrossRefDirect Link - Knox, C.A.P., B. Sonthayanon, G.R. Chandra and S. Muthukrishnan, 1987. Structure and organization of two divergent α-amylase genes from barley. Plant Mol. Bio., 9: 3-17.
CrossRefDirect Link - Laurie, S., R.S. McKibbin and N.G. Halford, 2003. Antisense SNF1-related (SnRK1) protein kinase gene represses transient activity of an α-amylase (α-Amy2) gene promoter in cultured wheat embryos. J. Exp. Bot., 54: 739-747.
PubMedDirect Link - Lazarus, C.M., D.C. Baulcombe and R.A. Martienssen, 1985. α-amylase genes of wheat are two multigene families which are differentially expressed. Plant Mol. Biol., 5: 13-24.
CrossRefDirect Link - Li, C., P. Ni, M. Francki, A. Hunter and Y. Zhang et al., 2004. Genes controlling seed dormancy and pre-harvest sprouting in a rice-wheat-barley comparison. Funct. Integr. Genomics, 4: 84-93.
CrossRefPubMedDirect Link - Li, C.D., A. Tarr, R.C.M. Lance, S. Harasymow and J. Uhlmann et al., 2003. A major QTL controlling seed dormancy and pre-harvest sprouting/grain α-amylase in two-rowed barley (Hordeum vulgare L.). Aust. J. Agric. Res., 54: 1303-1313.
CrossRefDirect Link - Lin, R.S., R.D. Horsley and P.B. Schwarz, 2008. Associations between caryopsis dormancy, α-amylase activity and pre-harvest sprouting in barley. J. Cereal Sci., 48: 446-456.
CrossRefDirect Link - Matthies, I.E., S. Weise and M.S. Roder, 2009. Association of haplotype diversity in the α-amylase gene amy1 with malting quality parameters in barley. Mol. Breeding, 23: 139-152.
CrossRefDirect Link - Mitsui, T. and K. Itoh, 1997. The α-amylase multigene family. Trends in Plant Sci., 2: 255-261.
CrossRefDirect Link - Murray, M.G. and W.F. Thompson, 1980. Rapid isolation of high molecular weight plant DNA. Nucl. Acids Res., 8: 4321-4326.
CrossRefPubMedDirect Link - Mrva, K. and D.J. Mares, 1996. Expression of late maturity α-amylase in wheat containing gibberellic acid insensitivity genes. Euphytica, 88: 69-76.
CrossRefDirect Link - Mrva, K. and D.J. Mares, 2001. Quantitative trait locus analysis of late maturity α-amylase in wheat using the doubled haploid population Cranbrook Halberd. Aust. J. Agric. Res., 52: 1267-1273.
CrossRefDirect Link - Mrva, K., J. Cheong, B. Yu, H.Y. Law and D. Mares, 2009. Late maturity α-amylase in synthetic hexaploid wheat. Euphytica, 168: 103-411.
CrossRef - Sugimoto, N., G. Takeda, Y. Nagato and J. Yamaguchi, 1998. Temporal and spatial expression of the α-amylase gene during seed germination in rice and barley. Plant Cell Physiol., 39: 323-333.
Direct Link - Tamura, K., J. Dudley, M. Nei and S. Kumar, 2007. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol., 24: 1596-1599.
CrossRefPubMedDirect Link