
Mulugeta Seyoum
Department of Horticulture and Plant Science, College of Agriculture and Veterinary Medicine, Jimma University, P. O. Box 307, Jimma, Ethiopia
Sentayehu Alamerew
Department of Horticulture and Plant Science, College of Agriculture and Veterinary Medicine, Jimma University, P. O. Box 307, Jimma, Ethiopia
Kassahun Bantte
Department of Horticulture and Plant Science, College of Agriculture and Veterinary Medicine, Jimma University, P. O. Box 307, Jimma, Ethiopia
Journal of Plant Sciences
Year: 2012 | Volume: 7 | Issue: 1 | Page No.: 13-22







ABSTRACT
Field experiments were conducted using fourteen rice genotypes during the main rainy seasons of 2009 and 2010 at three rainfed upland locations of Southwest Ethiopia to estimate the genetic variability, heritability and correlation coefficients of grain yield and yield contributing traits in upland rice. The experiments were laid down in a randomized complete block design with three replications. Highly significant (p<0.01) variation was obtained for days to 50% flowering, days to 85% maturity, plant height, panicle length, spikelets per panicle and thousand grains weight. Significant difference (p<0.05) were noted for panicles per plant, grains per panicle, total spikelet fertility and grain yield. Days to 50% flowering, plant height, grains per panicle, spikelets per panicle, thousand grains weight and grain yield showed relatively high GCV and PCV estimates. High heritability was obtained for plant height (92.17%), followed by 50% flowering (90.16%), thousand grains weight (83.17%), days to 85% maturity (82.45%), panicle length (79.25%) and spikelet per panicle (60.25%) which indicates high heritable portion of variation. High to medium estimates of heritability and genetic advance were obtained for plant height, days to 50% flowering, panicles per plant, spikelets per panicle, grains per panicle and thousand grains weight, indicating the roles of additive gene action and a good scope of selection using their phenotypic performance. Grains per panicle had maximum positive direct effect and highly significant (r = 0.906**) genotypic correlation coefficient with grain yield. The present study revealed that for increasing rice yield in upland ecology, a genotype should possess more number of grains per panicles, tillers per plant and panicle per plant, high spikelet fertility and large panicle size.
PDF Abstract XML References Citation
Received: December 13, 2011;
Accepted: December 22, 2011;
Published: March 09, 2012
How to cite this article
Mulugeta Seyoum, Sentayehu Alamerew and Kassahun Bantte, 2012. Genetic Variability, Heritability, Correlation Coefficient and Path Analysis for Yield and Yield Related Traits in Upland Rice (Oryza sativa L.). Journal of Plant Sciences, 7: 13-22.
DOI: 10.3923/jps.2012.13.22
URL: https://scialert.net/abstract/?doi=jps.2012.13.22
DOI: 10.3923/jps.2012.13.22
URL: https://scialert.net/abstract/?doi=jps.2012.13.22
INTRODUCTION
Rice is the fastest growing source of food in Africa. During the past three decades rice grain has seen a steady increase in consumption and demand given its important place in the strategic food security planning policies of many African countries (Norman and Otoo, 2003; Africa Rice Center, 2007; Forum for Agricultural Research in Africa, 2009). In order to meet the fastest growing demand for rice grain, development of high yielding genotypes with desirable agronomic traits for diverse ecosystem is therefore a necessity. However, grain yield is a complex trait, controlled by many genes, environmentally influenced and determined by the magnitude and nature of their genetic variability in which they grow (Singh et al., 2000). In addition, grain yield is related with other characters such as plant type, growth duration and yield components (Yoshida, 1981).
Genetic variability among traits is important for breeding and in selecting desirable types. The low heritability of grain yield characters made selection for high yielding varieties possible usually using various components traits associated with yield (Atlin, 2003). Therefore, character association of component traits with yield and among themselves is very important. The relationship between rice yield and yield component traits has been studied widely at phenotypic level. Sadeghi (2011) observed positive significant association of grain yield with grains per panicle, days to maturity, number of productive tillers and days to flowering. Ullah et al. (2011) noted that grain yield was positively and significantly associated with panicle length and grains per panicle. Hairmansis et al. (2010) also recorded a positive and significant association of grain yield with filled grains per panicle, spikelets per panicle and spikelet fertility.
Path coefficient analysis partitions the genetic correlation between yield and its component traits into direct and indirect effects and hence has effectively been used in identifying useful traits as selection criteria to improve grain yield in rice (Mustafa and Elsheikh, 2007; Kole et al., 2008; Akinwale et al., 2011; Sadeghi, 2011). Grain yield has been reported to be influenced by high direct positive effects of productive tillers, days to flowering and grains per panicle (Sadeghi, 2011), filled grains per panicle (Hairmansis et al., 2010), panicles per plant and grains per panicle (Mustafa and Elsheikh, 2007; Akinwale et al., 2011), panicles per panicle, grains per panicle, plant height and days to flowering (Kole et al., 2008).
Heritability of a trait is important in determining its response to selection. It was found out earlier that genetic improvement of plants for quantitative traits requires reliable estimates of heritability in order to plan an efficient breeding program. The broad sense heritability is the relative magnitude of genotypic and phenotypic variance for the traits and it gives an idea of the total variation accounted to genotypic effect (Allard, 1960). Akinwale et al. (2011) recorded broad sense heritability estimates of 95.1% for days to flowering, 92.1% for days to maturity, 72.4% for plant height, 72.1% for grain yield, 70.5% for grains per panicle, 59.4% for panicle per panicle, 53.6% for panicle length, 21.7% for thousand grain weight and 19.2% for tillers per plant. Sadeghi (2011) also reported broad sense heritability estimates of 98.9% for days to maturity, 93.3% for days to flowering, 93.77% for thousand grain weight, 93.15% for panicle length, 83.08% for grains per panicle, 70.2% for productive tillers, 69.21% for plant height and 68.19% for grain yield.
Most effective yield component breeding to increase grain yield could be achieved, if the components traits are highly heritable and positively correlated with grain yield. However, it is very difficult to judge whether observed variability is highly heritable or not. Moreover, knowledge of heritability is essential for selection based improvement as it indicates the extent of transmissibility of a character into future generations (Sabesan et al., 2009; Ullah et al., 2011). However, information on relationship of grain yield and yield contributing traits for upland rice improvement program of Southwest Ethiopia agro-ecology is not available. In view of the above gaps, the present study was undertaken to investigate the genetic variability, heritability and association between grain yield and yield related traits as a basis for selection of high yielding rice genotypes in upland ecology.
MATERIALS AND METHODS
Experimental sites, genotypes and procedures: Field experiments were carried out at three upland rainfed locations of Southwest Ethiopia: Sbebe (7°28' N, 36°25' E; altitude: 1370 m; annual rainfall: 1420 mm), Gomma-2 (7°57' N, 36°38' E; altitude: 1497 m; annual rainfall: 1470.4 mm) and Eladale (7°42' N, 36°48' E; altitude: 1813 m; annual rainfall: 1616 mm) during the main rainy seasons of 2009 and 2010. Fourteen rice genotypes obtained from SASAKAWA Global-2000, Ethiopia and from rice growing farmers: NERICA-1, NERICA-2, NERICA-3, NERICA-4, NERICA-7, NERICA-12, NERICA-13, NERICA-14 and NERICA-18, FOFIFA-3730, FOFIFA-3737, FOFIFA-4129, IAC-164 (Gumara) and X-Jigna (local) were used as experimental materials. The experiment was laid in a Randomized Complete Block Design (RCBD) with three replications. A five-row plot with size of 1.2x5 m was used. Dry seeds were hand drilled in rows 25 cm apart at a rate of 60 kg ha-1. Thinning was carried out at 3-4 leaf stage to maintain single plant per hill at a spacing of 10 cm. Mineral fertilizers were applied at a rate of 64 kg N and 46 kg P2O5 ha-1 as Urea and Di-Ammonim Phosphate (DAP) at each location. Nitrogen was applied in two equal splits: half at sowing and half at panicle initiation (booting) stage. Total dose of P2O5 was applied basal. Three manual weeding were carried during each season.
Data collection and analysis: Observation and data record for all traits studied was made based on the Standard Evaluation System for rice (IRRI, 1996). Plant height, panicle length, tillers per plant, panicles per plant, grains per panicle and total spikelet fertility were measured before physiological maturity from randomly selected 10 sample plants in the middle three rows of each plot. Data on days to 50% flowering and days to 85% maturity were also collected on a plot basis. At maturity, grain yield was taken from the three central rows and yield was adjusted at 14% moisture level. Thousand-grain weight was determined from bulked grains collected from central rows of each plot and recorded on 14% seed moisture basis. The data collected for all traits studied were subjected to analysis of variance for RCBD using GENRES3 (1994) version 7.01 and SAS (2003) version 9.1. The path and correlation coefficient analyses were done following methods of Dewey and Lu (1959). The phenotypic and genotypic coefficient of variability was computed as per Burton (1952). Heritability in broad sense was done using the formula suggested by Lush (1940). Genetic advance was worked out as per the formula given by Johnson et al. (1955).
RESULT AND DISCUSSION
Analysis of variance revealed significant differences among genotypes for all traits studied except for tillers per plant (Table 1), indicating presence of considerable amount of genetic variation among the study materials. The magnitude of variation between genotypes was reflected by high values of mean and range for genotype traits studied (Table 2). High genetic variability for different quantitative traits in rice was also reported earlier by Khan et al. (2009), Umadevi et al. (2009), Akinwale et al. (2011) and Ullah et al. (2011).
Coefficients of variation: The results (Table 2) revealed that the estimates of phenotypic coefficient of variation (PCV) were slightly higher than those of genotypic coefficient of variation (GCV) for all the traits studied. The extent of the environment influence on traits is explained by the magnitude of the difference between GCV and PCV. Large differences between GCV and PCV values reflect high environmental influence on the expression of traits. In this study, slight differences indicated minimum environmental influence and consequently greater role of genetic factors on the expression of traits.
| Table 1: | Analysis of variance for different traits in upland rice |
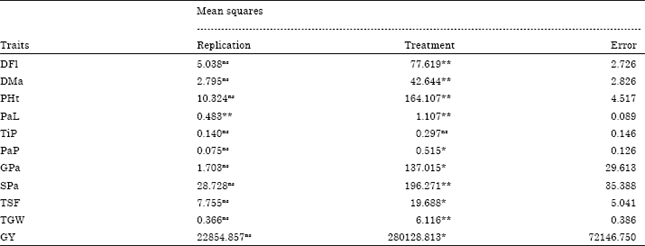 | |
| * Significant at 5% level of probability; ** Significant at 1% level of probability; ns = non significant; DFl: Days to 50% flowering; DMa: Days to 85% maturity; PHt: Plant height (cm); PaL: Panicle length (cm); TiP: Tillers per plant (No.); PaP: Panicles per plant (No.); GPa: Grains per panicle (No.); SPa: Spikelets per panicle (No.); TSF: Total spikelet fertility (%); TGW: Thousand grain weight (g); GY: Grain yield (kg ha-1) | |
Similar observations were also noted earlier by Mustafa and Elsheikh (2007) and Kole et al. (2008) in rice. In contrary to the present study which showed close values between GCV and PCV Akinwale et al. (2011) and Sadeghi (2011) recorded wide differences. The GCV ranged from 2.6045 for tillers per plant to 8.7634 for grain yield, whereas PCV ranged from 2.9311 for days to maturity to 12.5187 for grain yield. Relatively, days to 50% flowering, plant height, grains per panicle, spikelets per panicle, thousand grains weight and grain yield showed high GCV and PCV estimates. The lowest GCV was observed on days to 85% maturity and tillers per plant, whereas days to 85% maturity had the lowest PCV values. Akinwale et al. (2011) and Ullah et al. (2011) reported moderate to low GCV and PCV estimates for different quantitative traits in rice.
Heritability: In the estimates of broad sense heritability were varied from 25.82 to 92.17% (Table 2). The present study, 50% flowering, days to 85% maturity, plant height, panicle length, spikelet per panicle and thousand grains weight exhibited high heritability. High heritability suggests high component of heritable portion of variation that can be exploited by breeders in the selection of superior genotypes on the basis of phenotypic performance. Panicles per plant, grains per panicle, total spikelet fertility and grain yield exhibited moderate heritability which indicates the possibility of using for rice improvement program, but their expression can be influenced very much by the environment. High to moderate heritability was reported for different quantitative traits studied in rice (Zahid et al., 2006; Kole et al., 2008; Khan et al., 2009; Akinwale et al., 2011; Sadeghi, 2011). The study revealed very low estimate of heritability for tillers per plant which supports the results of Mustafa and Elsheikh (2007) and Akinwale et al. (2011) Very low heritability indicates greater role of environment on the expression of the trait, therefore, direct selection for tillers per plant will be ineffective.
The estimates of genetic advance as percent of mean (Table 2) were high for plant height, grain yield, grains per panicle, thousand grain weight, spikelets per panicle and days to 50% flowering, respectively; moderate for panicles per plant, panicle length, total spikelet fertility, days to maturity; and very low for tillers per plant.
| Table 2: | Estimates of parameters of variability for different traits in upland rice |
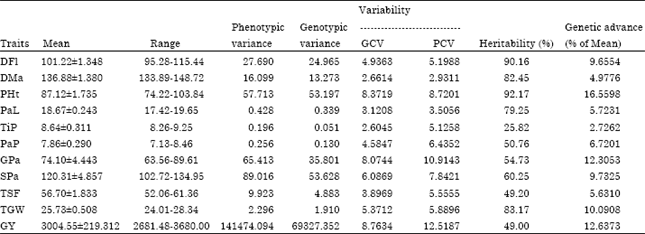 | |
| DFl: Days to 50% flowering; DMa: Days to 85% maturity; PHt: Plant height (cm); PaL: Panicle length (cm); TiP: Tillers per plant (No.); PaP: Panicles per plant (No.); GPa: Grains per panicle (No.); SPa: Spikelets per panicle (No.); TSF: Total spikelet fertility (%); TGW: Thousand grain weight (g); GY: Grain yield (kg ha-1) | |
These results were supported by earlier finding results (Kole et al., 2008; Akinwale et al., 2011; Sadeghi, 2011). Since high heritability does not always indicate high genetic gain, heritability with genetic advance should be used in predicting selection of superior genotypes (Ali et al., 2002).
In this study high to medium estimates of heritability and high genetic advance were obtained for plant height, days to 50% flowering, panicles per plant, spikelet per panicle, grains per panicle, thousand grains weight and grain yield which suggests these traits could be considered as favorable attributes for upland rice improvement through selection. Likewise, the high heritability combined with high genetic advance could be regarded as an indication of additive gene action and the consequent high-expected genetic gain from selection for these characters. High heritability and high genetic advance records were also reported earlier in rice for panicles per plant and plant height (Sadeghi, 2011), for panicles per plant, grains per panicle and grain yield (Kole et al., 2008) and for grains per panicle, thousand grain weight and grain yield (Ullah et al., 2011).
High heritability estimates with low genetic advance observed for days to 85% maturity, panicle length and total spikelet fertility indicates non-additive type of gene action and the genotype by environment interaction play a significant role in the expression of the trait. In agreement with the present findings high heritability with low genetic advance was observed for days to maturity and panicle length (Akinwale et al., 2011), for panicle length (Ullah et al., 2011) and for spikelet fertility (Qamar et al., 2005). Very low heritability and genetic advance for tillers per plant observed may be due to non-additive gene action and great influence of the environment on the trait. These findings were supported by Akinwale et al. (2011). Relatively high GCV, PCV, broad sense heritability and genetic advance obtained for days to flowering, plant height, grains per panicle, spikelet per panicle and thousand grain weight suggested these characters could be transmitted to the progeny when hybridization would be conducted and phenotypic based selection would be effective.
Correlation coefficients: The results of simple linear correlation coefficients between all pairs of traits as shown in Table 3 reveals grain yield had positive and significant association with grains per panicle (r = 0.847**), spikelets per panicle (r = 0.638*) and total spikelet fertility (r = 0.767*).
| Table 3: | Simple linear correlation coefficient between pairs of all traits in upland rice |
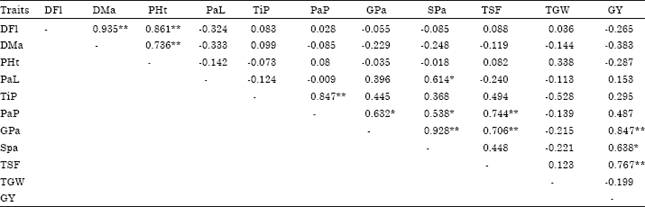 | |
| * Significant at 5% level of probability; ** Significant at 1% level of probability; DFl: Days to 50% flowering; DMa: Days to 85% maturity; PHt: Plant height (cm); PaL: Panicle length (cm); TiP: Tillers per plant (No.); PaP: Panicles per plant (No.); GPa: Grains per panicle (No.); SPa: Spikelets per panicle (No.); TSF: Total spikelet fertility (%); TGW: Thousand grain weight (g); GY: Grain yield (kg ha-1) | |
| Table 4: | Phenotypic (P) and genotypic (G) correlation coefficients for different traits in upland rice |
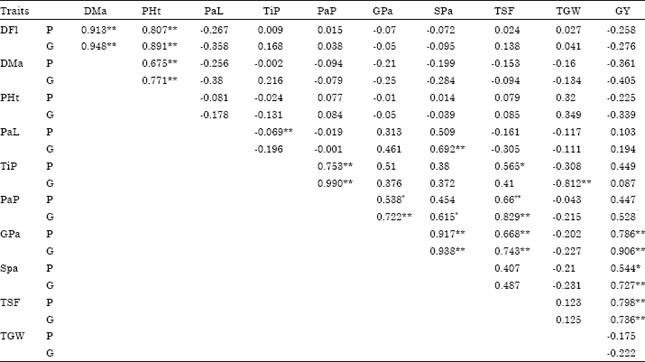 | |
| * Significant at 5% level of probability; ** Significant at 1% level of probability; DFl: Days to 50% flowering; DMa: Days to 85% maturity; PHt: Plant height (cm); PaL: Panicle length (cm); TiP: Tillers per plant (No.); PaP: Panicles per plant (No.); GPa: Grains per panicle (No.); SPa: Spikelets per panicle (No.); TSF: Total spikelet fertility (%); TGW: Thousand grain weight (g); GY: Grain yield (kg ha-1) | |
The results are in conformity with Hairmansis et al. (2010) for grains per panicle, spikelets per panicle and spikelet fertility and Ullah et al. (2011) for panicle length and grains per panicle. Genotypic and phenotypic correlation coefficients (Table 4) indicated that the genotypic and the phenotypic correlation coefficients showed similar trend but genotypic correlation coefficients were of higher in magnitude than the corresponding phenotypic correlation coefficients which might be due to the masking or modifying effect of environment on character association at the genetic level (Singh, 1980; Sarawgi et al., 1997).
| Table 5: | Path coefficient showing direct and indirect effects of traits on grain yield of upland Rice |
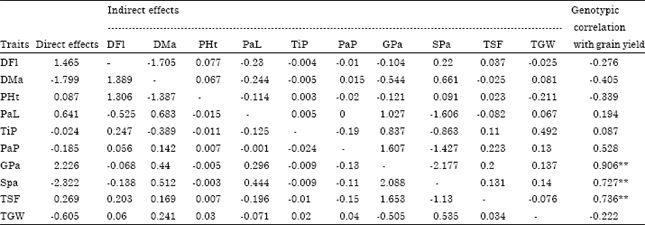 | |
| Residual effects, R = 0.13779; * Significant at 5% level of probability; ** Significant at 1% level of probability; ns = non significant; DFl: Days to 50% flowering; DMa: Days to 85% maturity; PHt: Plant height (cm); PaL: Panicle length (cm); TiP: Tillers per plant (No.); PaP: Panicles per plant (No.); GPa: Grains per panicle (No.); SPa: Spikelets per panicle (No.); TSF: Total spikelet fertility (%); TGW: Thousand grain weight (g); GY: Grain yield (kg ha-1) | |
This finding agreed with those of Zahid et al. (2006), Agahi et al. (2007) and Kole et al. (2008).
Grain yield was found to be positively and significantly associated with grains per panicle, spikelets per panicle and total spikelet fertility both at genotypic and phenotypic levels, indicating the importance of these traits for yield improvement in upland rice. The results are supported by Rokonuzzman et al. (2008) and Khan et al. (2009) for grains per panicle, by Kole et al. (2008) and Qamar et al. (2005) for panicles number and by Ramakrishman et al. (2006) for spikelet fertility. Grain yield had negative and non-significant correlation with days to 50% flowering, days to 85% maturity, plant height and thousand grains weight both at genotypic and phenotypic levels, whereas it showed positive and non-significant association with tiller per plant, panicle length and panicles per plant both at genotypic and phenotypic levels. Similar observations were reported for days to 50% flowering (Kole et al., 2008), plant height (Qamar et al., 2005) and thousand grains weight (Ullah et al., 2011), tillers per plant (Wattoo et al., 2010), panicle length (Kole et al., 2008), panicles per plant (Ullah et al., 2011). On the contrary, Khan et al. (2009) and Qamar et al. (2005) reported positive and non-significant association between grain yield and days to maturity.
Path coefficient analysis: Genotypic correlation coefficients were partitioned by using method of path analysis to find out the direct and indirect effects of yield contributing traits towards the grain yield. From the path analysis (Table 5), it was revealed that grains per panicle (2.226) exhibited maximum positive direct effect on grain yield followed by days to 50% flowering (1.465), panicle length (0.641), total spikelet fertility (0.269) and plant height (0.087). The direct effects of days to 85% maturity, tillers per plant, panicles per plant, spikelets per panicle and thousand grains weight were negative. Panicle length, tillers per plant, panicles per plant, spikelet per panicle and total spikelet fertility had positive indirect effect on grain yield through grains per panicle. The indirect effects of grains per panicle through other traits indicated that direct selection using grains per panicle to select high yielding genotypes will be effective. Grains per panicle showed the highest positive direct effect and genotypic correlation (r = 0.906) with grain yield. This strong genetic correlation resulted in high positive direct effect on grain yield. Many research works revealed similar findings (Ramakrishman et al., 2006; Zahid et al., 2006; Mustafa and Elsheikh, 2007; Khan et al., 2009; Hairmansis et al., 2010). On the other hand, the highest positive direct effect has been reported for number of productive tillers (Agahi et al., 2007; Rokonuzzman et al., 2008) for days to maturity (Qamar et al., 2005), for panicle number (Kole et al., 2008), for chlorophyll content (Ullah et al., 2011) and for percentage fertile spikelets (Agbo and Obi, 2005) in rice.
The residual effect was 0.13779 indicated that the contribution of component characters on grain yield was 86.2%, by the ten characters studied in path analysis, the rest 13.85% was the contribution of other factors, such as traits not studied.
CONCLUSION
The present study results indicated that there is adequate genetic variability present in the material studied. The GCV, PCV, broad sense heritability and genetic advance suggested days to 50% flowering, plant height, grains per panicle, spikelet per panicle and thousand grains weight were important yield influencing traits. Grains per panicle had the maximum positive direct effect and the highest genotypic correlation coefficient, followed by total spikelet fertility. Panicle length, tillers per plant, panicles per plant, spikelet per panicle and total spikelet fertility had positive indirect effect on grain yield through grains per panicle. Grain per panicle is the most important trait which should be given due attention in making selection effective for high yielding genotypes in upland ecology. Therefore, from present study it can be forwarded that for increasing rice grain yield in upland ecology, a genotype should possess more number of grains per panicles, tillers per plant and panicle per plant, high spikelet fertility and large panicle size.
ACKNOWLEDGMENTS
The authors acknowledge the Jimma University, College of Agriculture and Veterinary Medicine and Japan International Cooperation Agency-Farmers Research Group for financing the study and SASAKAWA Global-2000 for the supply of NERICA and FOFIFA seeds.
REFERENCES
- Agahi, K., M.H. Fotokian, and E.Farshadfar, 2007. Correlation and path coefficient analysis for some yield-related traits in rice genotypes (Oryza sativa L.). Asian J. Plant Sci., 6: 513-517.
CrossRef - Agbo, C.U. and I.U. Obi, 2005. Yield and yield component analysis of twelve upland rice genotypes. J. Agric. Food, Environ. Extens., 4: 29-33.
Direct Link - Akinwale, M.G., G. Gregorio, F. Nwilene, B.O. Akinyele, S.A. Ogunbayo and A.C. Odiyi, 2011. Heritability and correlation coefficient analysis for yield and its components in rice (Oryza sativa L.). Afr. J. Plant Sci., 5: 207-212.
Direct Link - Allard, R.W., 1960. Principles of Plant Breeding. 1st Edn., John Wiley and Sons Inc., New York pp: 372-372.
CrossRefDirect Link - Dewey, D.R. and K.H. Lu, 1959. A correlation and path-coefficient analysis of components of crested wheatgrass seed production. Agron. J., 51: 515-518.
CrossRefDirect Link - Hairmansis, A., B. Kustianto, Supartopo and Suwarno, 2010. Correlation analysis of agronomic characters and grain yield of rice for tidal swamp areas. Indonesian J. Agric. Sci., 11: 11-15.
Direct Link - IRRI, 1996. Standard Evaluation System for Rice. 4th Edn., International Rice Research Institute, Manila, Philippines, Pages: 52.
Direct Link - Khan, A.S., M. Imran and M. Ashfaq, 2009. Estimation of genetic variability and correlation for grain yield components in rice (Oryza sativa L.). Am. Eurasian J. Agric. Environ. Sci., 6: 585-590.
Direct Link - Kole, P.C., N.R. Chakraborty and J.S. Bhat, 2008. Analysis of variability, correlation and path coefficients in induced mutants of aromatic non-basmati rice. Trop. Agric. Res. Exten., 11: 60-64.
CrossRefDirect Link - Zia-Ul-Qamar, A.A. Cheema, M. Ashraf, M. Rashid and G.R. Tahir, 2005. Association analysis of some yield influencing traits in aromatic and non aromatic rice. Pak. J. Bot., 37: 613-627.
Direct Link - Ramakrishman, S.H., C.R. Anandakumar, S. Saravanan and N. Malini, 2006. Association analysis of some yield traits in rice (Oryza sativa L.). J. Applied Sci. Res., 2: 402-404.
Direct Link - Sabesan, T., R. Suresh and K. Saravanan, 2009. Genetic variability and correlation for yield and grain quality characters of rice grown in coastal saline low land of Tamilnadu. Electron. J. Plant Breed., 1: 56-59.
Direct Link - Sadeghi, S.M., 2011. Heritability, phenotypic correlation and path coefficient studies for some agronomic characters in landrace rice varieties. World Appied Sci. J., 13: 1229-1233.
Direct Link - Ullah, M.Z., M.K. Bashar, M.S.R. Bhuiyan, M. Khalequzzaman and M.J. Hasan, 2011. Interrelationship and cause-effect analysis among morpho-physiological traits in biroin rice of Bangladesh. Int. J. Plant Breed. Genet., 5: 246-254.
CrossRefDirect Link - Wattoo, J.I., A.S. Khan, Z. Ali, M. Babar, M. Naeem, M. Aman ullah and N. Hussain, 2010. Study of correlation among yield related traits and path coefficient analysis in rice (Oryza sativa L.). Afr. J. Biotechnol., 9: 7853-7856.
CrossRefDirect Link - Yoshida, S., 1981. Fundamentals of Rice Crop Science. 1st Edn., International Rice Research Institute, Phillipines, ISBN: 971-104-052-2, Pages: 267.
Direct Link - Zahid, M.A., M. Akhter, M. Sabar, Z. Manzoor and T. Awan, 2006. Correlation and path analysis studies of yield and economic traits in basmati rice (Oryza sativa L.). Asian J. Plant Sci., 5: 643-645.
CrossRefDirect Link