
H. Reguieg Yssaad
Laboratoire de Biologie Faculte des Sciences, Universite de Mostaganem, route de Belhacel BP 923 27000, Mostaganem Algerie
A. Chibani
Laboratoire de Biologie Faculte des Sciences, Universite de Mostaganem, route de Belhacel BP 923 27000, Mostaganem Algerie
A. Bachir Bouyadjra
Laboratoire de Physiologie Vegetale, Faculte des Sciences, Universite d`Oran, Senia 31 000 Oran, Algerie
Journal of Plant Sciences
Year: 2012 | Volume: 7 | Issue: 1 | Page No.: 1-12







ABSTRACT
In order to improve the agricultural product, especially leguminous plants (Vicia faba L.) in sandy soils, we used bentonite to ameliorate their physical and chemicals properties. To evaluate the ecological advantage of this clay in these soils, an increasing amount of bentonite were added to the sandy soil, then the physical and chemical characteristics of the mixture were identified such as pH, EC (Electrical Conductivity), total calcareous, active calcareous, total phosphorus, organic carbon, organic matter, total nitrogen and exchange cations capacity. The drought causes a rise of salts in the rhizosphere particularly in arid and semi-arid regions. This work is an analysis of the response of the leguminous plant Vicia faba L. to increasing concentrations of bentonite and salt as well as to understand the variability of certain metabolic activities, mineral content and morpho-physiological behavior of the plant response to this abiotic stress. Results show that variability exists in physical and chemical characteristics and morphological growth of the plant according to the bentonite amount mixed in the sandy soil sample.
PDF Abstract XML References Citation
Received: November 25, 2011;
Accepted: December 29, 2011;
Published: March 09, 2012
How to cite this article
H. Reguieg Yssaad, A. Chibani and A. Bachir Bouyadjra, 2012. Effect of Salinity and Bentonite on Mineral Soil Characteristics: Behavior Study of Leguminous Plant (Vicia faba L.). Journal of Plant Sciences, 7: 1-12.
DOI: 10.3923/jps.2012.1.12
URL: https://scialert.net/abstract/?doi=jps.2012.1.12
DOI: 10.3923/jps.2012.1.12
URL: https://scialert.net/abstract/?doi=jps.2012.1.12
INTRODUCTION
The progressive reduction of vegetation cover in arid and semi-arid areas, due to desertification and soil erosion becomes a major problem in ecosystems of these regions (Martinez et al., 2005). Salinity is a worldwide growing environmental problem, particularly in North African countries. This is an important abiotic factor limiting plant growth and productivity (Khan and Panda, 2008) and more than 40% of cultivated land in arid and semi-arid areas are affected by salinity (Hamdy, 1999; Munns, 2002).
The decrease of water resources in these regions and the expansion of irrigated land cause salinization of soils (Munns et al., 2006; Ben Naceur et al., 2001; Rochdi et al., 2005; De Araujo et al., 2006). Salinity is not only related to climate conditions (Djili and Daoud, 2000) but also to poorly controlled cultivation practices, such as the use of large quantities of poor quality water (Mouhhouche and Boulassal, 1999; Belkhodja and Bidai, 2004). Moreover, the excessive use of chemical fertilizers also causes soil salinity (Zid and Grignon, 1991; Smith and Compton, 2004; Qadir and Oster, 2004). The presence of high quantities of salts in the soil does not only limit productivity, but also causes an ionic, osmotic and nutritional stress (Levigneron et al., 1995).
Exposed to these environmental constraints, plants develop strategies for tolerance and adjustment reactions resulting in changes in metabolic, physiological and morphological nature (Jacoby, 1994; Qian et al., 2001; Liu and Baird, 2003; Aleberto et al., 2005). One major mechanism of adaptation to ionic and osmotic stress results in an active accumulation of K+, Na+ by the plant (Wang et al., 2002; Parida and Das, 2005), Cl¯ (Munns et al., 2006; Teakle et al., 2007) and certain amino acids such as proline (Belkhodja and Bidai, 2004; Kartashov et al., 2008).
For a sustainable agriculture will be more advantageous to choose a rational management of cultivated lands, based on a deep understanding of biological interactions and using available natural resources.
It is in this perspective that the action of setting value of the cultivated land integrates by introducing the clay-rich bentonite. This action will lead to increase cation exchange capacity (Petr, 1985; Dejou, 1987). Other authors assert that bentonite supplemented sandy soil has an improved physical and water properties (Benkhalifa and Daoud, 1998; Halilat and Tessier, 2006).
The aim of present research is to study the influence of some doses of bentonite, coupled with abiotic salinity constraints on the mineral and water behavior of Vicia faba L. a species of legume commonly grown in Algeria. The mineral analysis of broad beans leaves and roots allows the understanding the combined effects of salinity and bentonite on the mineral nutrition of this species.
MATERIALS AND METHODS
Plant material: The plant used was a local variety of broad beans (Vicia faba L.). The broad beans seeds were from the station of the Institute of Development Crops of Sidi Bel Abbes. The seeds were disinfected for 5 min with bleach 8% and rinsed several times with distilled water to remove any trace of chlorine. In a first stage, the germination of seeds was performed in plastic trays wrapped in moist filter paper and placed in an oven at a temperature of 22°C for 7 days.
METHODS
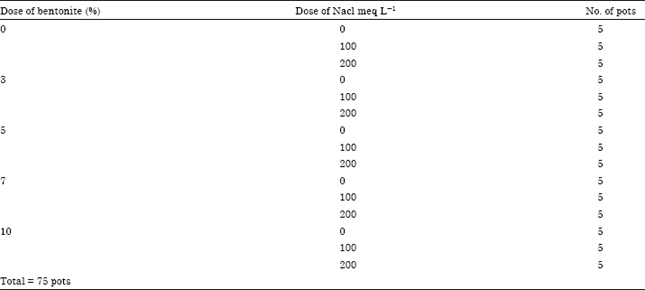
Substrate culture: The sand samples collected from the beach of Sidi Mansour were first sieved toobtain homogenous sand and to eliminate all waste then, washed repeatedly with tap water and acid solution to remove allcarbonates and chlorides. Then, this sand was rinsed several times with demineralised water to remove all traces of chlorides finally the sand was dried in open air. Adesalination test was carried out using silver nitrate to confirm the purity of the substrate. The bentonite in its natural granular form was previously milled using an electric grinder and passed through a sieve mesh of 2 mm to obtain a fine powder to facilitate its disposal and mixing. The prepared substrate consisted of two volumes of sand mixed with one volume of compost. Four doses of bentonite were used: 3, 5, 7 and 10%. The quantity of bentonite added to the treated soil matches with doses expressed in percentage of soil dry weight (Table 1). This substrate was vigorously mixed manually to obtain a homogeneous substrate. The pots used were made of plastic, the bottoms of which were covered with a layer of gravel to ensure good draining. Each pot was filled with 600 g of mixed substrate and arranged according to the method of complete randomized blocks with five replicates (Table 4).
Seedling transfer: After germination, seedlings were carefully transferred to pots (one seedling/pot) and placed in a green house located at the University of Oran, where temperature, humidity and wind factors are under control.
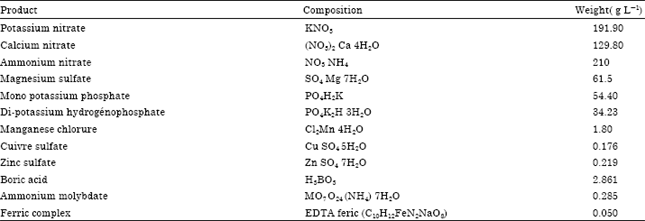
| Table 1: | Weight of bentonite of each dose used in substrate dry weight |
| Table 2: | Composition of the nutrient solution of Hoagland and Arnon (1938) |
 | |
| Table 3: | Composition of the saline solution |
 | |
| Table 4: | Experimental Dispositif adopted in the greenhouse |
 | |
| M±σm: Statistical average σ: Ecart type ns:not significant s: Significant ss: Highly significant | |
We proceeded to irrigation at 60% of the substrate retention capacity which is 80 mL per pot, after determining the retention capacity of the substrate-soil. Watering was done 3 times per week, twice with demineralised water and once with the nutrient solution (Hoagland and Arnon, 1938) diluted to 10-3, solution’s concentration is given in Table 2 and that of saline solution is given in Table 3.
The pots were divided into 5 treatments with bentonite (0, 3, 5, 7 and 10%). Each dose of bentonite receives 3 concentrations of sodium chloride (0, 100.200 meq L-1) with 5 replicates for each salt treatment.
 | |
| Fig. 1: | The seeds of Vicia faba L. |
The control pots were irrigated only with distilled water during the period of stress. After 35 days, application of stress was performed one week before plant samples were taken for analyses (Fig. 1).
Sample collection: Plant organs leaves, stems and roots were carefully separated, rinsed with tap water and dried with blotting paper. Each organ was weighed, wrapped in aluminum paper and dried in an oven at 80°C for 48 h. After that, dried samples were weighed and water content of organs was determined according to the following formula:
Extraction of minerals: The process of extraction and determination of minerals were carried out on leaves and roots of stressed plants and controls by the analytical method described by Lafon et al. (1996) which determines the elements composition of a plant by calcination and then the complete destruction of the organic material (Martin-Prevel et al., 1984). The residue was then analyzed. Measurements of Na+ and K+ contents were made by flame spectrophotometry.
RESULTS AND DISCUSSION
Sodium and potassium content (leaves and roots)
A-Sodium content
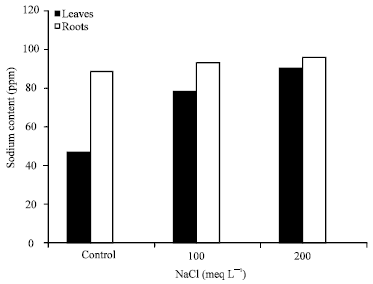
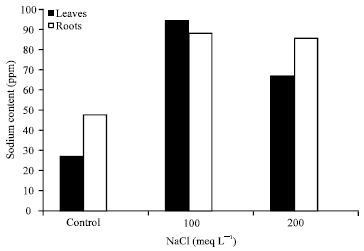
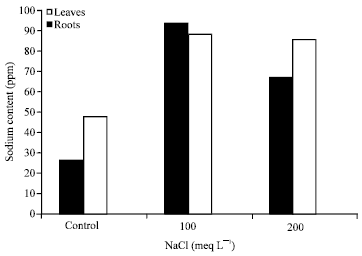
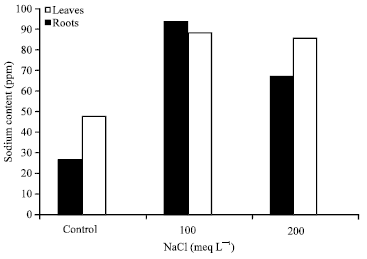
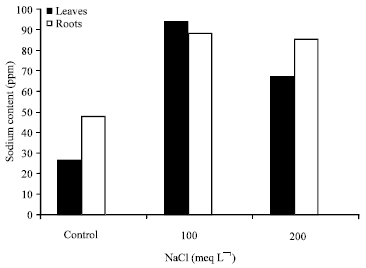
In bentonite-free substrate: The addition of NaCl in the irrigation solutions increased significantly the rate of Na+ in plant roots. The level of sodium was relatively less important in leaves. In roots we recorded more elevated concentration. In the control treatment recorded a very low rate of sodium of about 16.18 ppm in leaves. The accumulation of this ion in studied organs of broad beans increased proportionately with the dose of saline. The values increased to 35.84 ppm 100 meq L-1 and went up to 83.76 ppm when the salt concentration doubled.
The roots and leaves showed the same behavior in the presence of NaCl, control samples for roots recorded levels of sodium of 50.28 ppm, which passed to 72.84 then to 85.42 ppm for salt concentrations of 100 and 200 meq L-1, respectively (Fig. 2). We should note that sodium content in roots remained higher than that of leaves in bentonite-free substrates.
In 3% of bentonite substrate: In substrates treated with bentonite at 3%, levels of sodium obtained in studied plant organs became higher compared to the soil without bentonite (Fig. 3). Indeed, these soils at 3% of bentonite recorded sodium content of 17.38 ppm in leaves in non-stressed plants, this level increased to 52.2 and 61.68 ppm for salt concentrations of 100 and 200 meq L-1 . However, for roots a significant increase in this element was noticed up to 40.94 ppm in the absence of salt. Once the plant receives 100 meqL-1 a net increase of sodium was observed which goes up to 78.14 ppm. The level of sodium increased only to 9% in the roots 200 meq L-1.
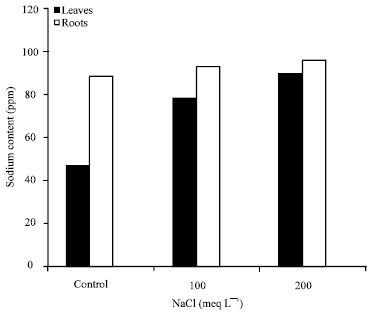 | |
| Fig. 2: | Sodium content in roots and leaves of broad beans in substrates without bentonite and salt-stressed for a week |
 | |
| Fig. 3: | Sodium content in roots and leaves of the broad beans in substrates at 3% of bentonite and salt-stressed for a week |
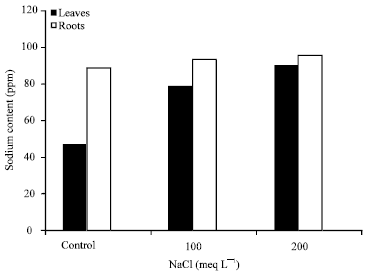 | |
| Fig. 4: | Sodium content in roots and leaves of the broad beans in substrates at 5% of bentonite and salt-stressed for a week |
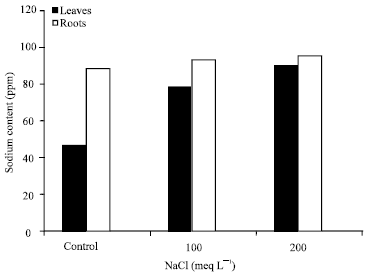 | |
| Fig. 5: | Sodium content in roots and leaves of the broad beans in substrates at 7% of bentonite and salt-stressed for a week |
At 5% of bentonite: According to the results (Fig. 4) and compared to the substrate at 3% of bentonite, a significant increase was noticed for the sodium levels in leaves and roots, 7.02 and 8.10 ppm dose of 100 meq L-1 saline and 14.56 ppm, 5.34 ppm dose 200 meq L-1, respectively for leaves and roots of the plants. The roots showed the highest levels of sodium for salt treatments (0, 100.200 meq L-1) (Fig. 4) while in leaves levels were less important where there is the lowest recorded content in the control (0 meq L-1). In the control substrate, sodium levels were still low ranging between 34.4 and 78.04 ppm.
At 7% of bentonite: The sodium levels found in the broad bean leaves on substrate at 7% of bentonite are relatively low ( 37.14 ppm) for non-stressed control plants. Then, a major change appears in the sodium level of the leaves at salt concentrations of 100 and 200 meq L-1 where the mean values vary between 76.92 and 61.56 ppm. This increase in sodium levels is due mainly the combined effect of the presence of bentonite and salt.
According to results recorded in soils with 7% of bentonite (Fig. 5) we noted that the levels of sodium in roots were higher compared to those recorded in leaves and this is true for non-stressed plants and those subjected to salt concentrations. As soon as plants receive saline solutions of 100 and 200 meq L-1, a considerable accumulation of Na+ is recorded in roots, where concentrations ranged from 89.28 to 90.64 ppm. Bentonite promoted the accumulation Na+ in roots, which explains the content variability of sodium in broad beans roots exposed to two variables salinity and bentonite.
At 10% of bentonite: The treatment of the substrate soil with 10% of bentonite resulted in a considerable load of sodium ions in leaves and roots. Over all, the increase in sodium is very consistent in both parts of the plant at 200 meq L-1 saline doses, still with maximum recorded values (89 and 92.6 ppm).
The sodium levels found in both plant organs (Fig. 6) were the highest compared to previous bentonite treatments and this is valid for unstressed plants and the two saline doses of 100 and 200 meq L-1. Moreover, in roots, sodium increases much more than in leaves. In addition, the Sodium levels analysed in soils amended with salt (NaCl) are much higher than those obtained in the plant without salt stress.
Potassium content
Without treatment with bentonite: In bentonite-free substrate the potassium plant content (Fig. 7) varies between 26.14 and 89.14 ppm with a remarkable difference in the potassium content between non-stressed the plants and those under saline stress. At 100 meq L-1 saline, potassium content became higher in leaves than in roots. In contrast, at 200 meq L-1 we noticed a significant drop in potassium in leaves compared to roots down to 87.58 ppm. The roots displayed the highest level of 89.14 ppm.
Moreover, under the application of salt stress, the content of potassium was much higher than that obtained in the salt-free culture.
At 3% of bentonite: According to results (Fig. 8) in substrate sat 3% of bentonite, there was a significant increase in potassium levels in leaves and roots at 0 and 100 meq L-1 compared with bentonite-free soil, however, these concentrations significantly decreased at 200 meq L-1 in both leaves and roots with values ranging from 68.48 to 57.46 ppm, respectively.
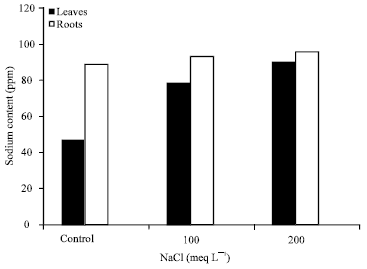 | |
| Fig. 6: | Sodium content in roots and leaves of the broad beans in substrates at 10% of bentonite and salt-stressed for a week |
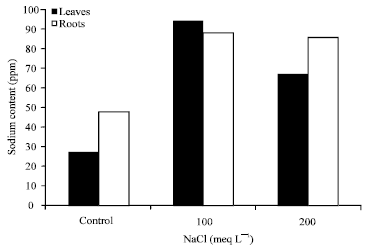 | |
| Fig. 7: | Potassium content in roots and leaves of the broad beans in substrates without bentonite and salt-stressed for a week |
 | |
| Fig. 8: | Potassium content in roots and leaves of the broad beans in substrates at 3% of bentonite and salt-stressed for a week |
This decrease of potassium was due to the strong absorption that occurred in the plant at the salt concentration of 100 meq L-1. In general, levels of potassium were more consistent in leaves than in roots even when maximum values are recorded in leaves (68.48 and 89.2 ppm).
At 5% of bentonite: According to the results (Fig. 9) and compared to the substrate at 3% of bentonite, a consequent reduction of the potassium contenting leaves was noted for all salt treatments, 32.22 ppm for the salt stress-free plant and 68.24 for the saline dose of 100 meq L-1 and 63.78 ppm for salt concentration of 200 meq L-1.
We noted that the accumulation of potassium in leaves and roots was higher in stressed plants than in the non-stressed plant. Whereas, potassium in plant roots increased regardless of saline treatment. Compared to substrate at 3% bentonite, potassium content remains low in non-stressed plants (55.8 ppm) at 100 meq L-1, potassium content in roots increases much more in cultivations of 200 meq L-1 salt.
 | |
| Fig. 9: | Potassium content in roots and leaves of the broad beans in substrates at 5% of bentonite and salt-stressed for a week |
 | |
| Fig. 10: | Potassium content in roots and leaves of the broad beans in substrates at 7% of bentonite and salt-stressed for a week |
At 7% of bentonite: The results obtained in plants that were not under saline stress how that the variation of potassium is very low between leaves and roots. The content of potassium was higher in plants of salt concentration of 100 and 200 meq L-1 where the values vary between 67.98 and 75.32 ppm, It was noted that the accumulations in this element was more substantial in roots than in leaves (Fig. 10). On the other hand for the dose of 200 meq L-1, it was observed that a significant decrease of this element up to 67.98 and 74.7 ppm for leaves and roots, respectively.
At 10% of bentonite: According to the results obtained in the substrate at 10% of bentonite (Fig. 11), it was noted that the potassium levels in leaves of non-stressed plants were the lowest compared to those of previous substrates. But this level remained high for the roots.
Moreover, in plants at 100 meq L-1 NaCl, potassium increased much more than in plants of 200 meq L-1 of salt. In addition, the analysed levels of potassium in leaves at 100 meq L-1 NaCl were much higher (93.86 ppm) than those obtained in roots (87.76 ppm). The accumulation of potassium was much higher in cultivations of 100 meq L-1 compared to 200 meq L-1.
 | |
| Fig. 11: | Potassium content in roots and leaves of the broad beans in substrates at 10% of bentonite and salt-stressed for a week |
CONCLUSION
The sodium content in the roots remains higher than that of the leaves in substrates without treatment with bentonite. Over all, the increase of sodium level is very consistent in the leaves and roots of non-stressed plants in substrates rich in bentonite. The accumulation of sodium is very significant in both organs of plants grown in the absence of bentonite to salt doses 100 and 200 meq L-1. The leaves accumulate more sodium under the combined effect of different doses of bentonite and practiced salt concentrations. Leaves and roots of plants grown in substrates of 3% bentonite and stressed to 200 meq L-1 NaCl accumulate lower levels of sodium than in substrates without bentonite. The addition of increasing concentrations of salt loads the bean leaves with Na+. In fact, the same results are mentioned in the work on this legume (Belkhodja, 1996; Taibi, 2009). The sodium accumulation occurs in plants grown in saline conditions and leaves are more vulnerable than roots, because salts are accumulated in the leaf edges, causing an ionic imbalance, deficiency symptoms and disturbance of metabolites (Tester and Davenport, 2003). The potassium content increases dramatically in plants subjected to a combination of salt and bentonite. Once the plants receive salt solutions of 100 and 200 meq L-1 potassium levels become high in leaves and roots of plants grown in substrates at 5 and 7% of bentonite. With the combination of bentonite and salts, plants record higher rates of potassium.
REFERENCES
- Aleberto, C., R. Laiz-Carrion, M.P.M. Del Rio, J. Maseguer, J.M. Mancera and M.A. Esteban, 2005. Salinity influences the humoral immune parameters of gilthead seabream (Sparus aurata L.). Fish Shellfish Immunol., 18: 255-261.
CrossRefPubMedDirect Link - De Araujo, S.A.M., J.A.G. Silveira, T.D. Almeida, I.M.A. Rocha, D.L. Morais and R.A. Viegas, 2006. Salinity tolerance of halophyte Atriplex nummularia L. grown under increasing NaCl levels. J. Agric. Environ. Eng., 10: 848-854.
Direct Link - Hoagland, D.R. and D.I. Arnon, 1938. The water culture method for growing plants without soil. Calif. Agric. Exp. Station, C347: 1-39.
Direct Link - Kartashov, A.V., N.L. Radyukina, Y.V. Ivanov, P.P. Pashkovskii, N.I. Shevyakova and V.V. Kuznetsov, 2008. Role of antioxidant systems in wild plant adaptation to salt stress. Russ. J. Plant Physiol., 55: 463-468.
CrossRef - Khan, M.H. and S.K. Panda, 2008. Alterations in root lipid peroxidation and antioxidative responses in two rice cultivars under NaCl-salinity stress. Acta Physiol. Planta., 30: 81-89.
CrossRefDirect Link - Munns, R., 2002. Comparative physiology of salt and water stress. Plant Cell Environ., 25: 239-250.
CrossRefDirect Link - Munns, R., R.A. James and A. Lauchli, 2006. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot., 57: 1025-1043.
CrossRefPubMedDirect Link - Parida, A.K. and A.B. Das, 2005. Salt tolerance and salinity effects on plants: A review. Ecotoxicol. Environ. Saf., 60: 324-349.
CrossRefPubMedDirect Link - Qadir, M. and J.D. Oster, 2004. Crop and irrigation management strategies for saline-sodic soils and waters aimed at environmentally sustainable agriculture. Sci. Total Environ., 323: 1-19.
CrossRefPubMedDirect Link - Smith, M. and J.S. Compton, 2004. Origin and evolution of major salts in the darling pans, Western Cape, South Africa. Applied Geochem., 19: 645-664.
CrossRefDirect Link - Teakle, N., T.J. Flowers, D. Real and T.D. Colmer, 2007. Lotus tenuis tolerates the interactive effects of salinity and waterlogging by 'excluding' Na+ and Cl- from the xylem. J. Exp. Bot., 58: 2169-2180.
CrossRef - Tester, M. and R. Davenport, 2003. Na+ tolerance and Na+ transport in higher plants. Ann. Bot., 91: 503-527.
CrossRefDirect Link - Wang, S., W. Zheng, J. Ren and C. Zhang, 2002. Selectivity of various types of salt-resistant plants for K+ over Na+. J. Arid Environ., 52: 457-472.
CrossRefDirect Link - Liu, X. and W.V. Baird, 2003. Differential expression of genes regulated in reponse to drought or salinity stress in sunflower. Crop Sci., 43: 678-687.
Direct Link - Ben Naceur, M., C. Rahmoune, H. Sdiri, M.L. Meddahi and M. Selmi, 2001. Effect of salt stress on germination, growth and grain yield in some North African wheat varieties. Drought, 12: 167-174.
Direct Link - Halilat, M.T. and D. Tessier, 2006. Improvement of sandy material water content by addition of bentonite. Cah. Agric., 15: 347-353.
Direct Link - Martinez, J.P., J.M. Kinet, M. Bajji and S. Lutts, 2005. NaCl alleviates polyethylene glycol-induced water stress in the halophyte species Atriplex halimus L. J. Exp. Bot., 56: 2421-2431.
CrossRef