
Qing Chen
Triticeae Research Institute, Sichuan Agricultural University, Yaan, Sichuan 625014, China
Pengfei Qi
Triticeae Research Institute, Sichuan Agricultural University, Yaan, Sichuan 625014, China
Renlin Xu
Agriculture and Agri-Food Canada, ECORC, Canada
James T. Tambong
Agriculture and Agri-Food Canada, ECORC, Canada
Zeinab R. Djama
Agriculture and Agri-Food Canada, ECORC, Canada
Wei Li
Triticeae Research Institute, Sichuan Agricultural University, Yaan, Sichuan 625014, China
Journal of Plant Sciences
Year: 2011 | Volume: 6 | Issue: 2 | Page No.: 52-65







ABSTRACT
Pseudomonads are of significant environmental importance as the members including plant and animal/human pathogens, xenobiotic degraders, plant growth promoters and biocontrol agents. Three typing methods were evaluated, utilising 143 strains of fluorescent pseudomonads isolated from maize-oat-alfalfa-alfalfa rotation plots in Canada, to assess their usefulness as tools to study the microbial community diversity. The methods used were repetitive element sequence-based PCR (rep-PCR) utilising BOX and Enterobacterial Repetitive Intergenic Consensus (ERIC) primers, another new PCR method using an engineered polymerase and 10-nucleotide “miniprimers” expands the scope of detectable sequences beyond those detected by standard methods using longer primers and Taq polymerase. Cluster analysis of the results clearly demonstrated the groups can be divided into two big clusters in line with the morphological variation. Rep-PCR proved to be highly discriminatory, interesting tools for the delineation or genotyping of bacterial species. We concluded that a combination of Box-, Eric- and Miniprimer-PCR results is a rapid and reliable alternative for discrimination among P. fluorescens isolates, contributing to epidemiological studies.
PDF Abstract XML References Citation
Received: March 08, 2011;
Accepted: April 20, 2011;
Published: June 23, 2011
How to cite this article
Qing Chen, Pengfei Qi, Renlin Xu, James T. Tambong, Zeinab R. Djama and Wei Li, 2011. Comparison of Three Typing Methods for Evaluating the Diversity of Pseudomonas fluorescens in the Rhizosphere. Journal of Plant Sciences, 6: 52-65.
DOI: 10.3923/jps.2011.52.65
URL: https://scialert.net/abstract/?doi=jps.2011.52.65
DOI: 10.3923/jps.2011.52.65
URL: https://scialert.net/abstract/?doi=jps.2011.52.65
INTRODUCTION
Plant growth-promoting rhizobacteria (PGPR) are root-associated bacteria that have the ability to improve the growth of plants when applied to seeds, soil, or subterranean plant parts (Shaukat et al., 2006). Growth promotion occurs as a result of direct stimulation of the plant, inhibition of plant pathogens, or induction of host defence mechanisms against pathogens (Thomashow and Weller, 1996; Van loon et al., 1998).
One of the most important and best-studied bacterial taxa in soil is the genus Pseudomonas (Noura et al., 2009). This genus includes several functional groups of environmental interest, such as plant growth promoters (Patten and Glick, 2002) plant pathogens (Samson et al., 1998) and xenobiotic degraders (Clausen et al., 2002). Moreover, Pseudomonas species can also play important roles as biological control agents against soil-borne plant pathogens (Siddiqui and Shakeel, 2006; Nezarat and Gholami, 2009). P. fluorescens is commonly found in soil and water, associated with food spoilage, in clinical speci-mens and also in association with plants and this species can be divided into five biovars (Soleimani et al., 2005).
Microbial DNA fingerprinting has been reported extensively using PCR-based techniques such as Amplified Fragment Length Polymorphism (AFLP) and is now in common use. Among PCR-based molecular markers, RAPD (random amplified polymorphic DNA) and repetitive sequence-based PCR genomic fingerprinting have been found to be particularly efficient for bacterial analysis (Olive and Bean, 1999; Tikoo et al., 2001; Coenye et al., 2002; Seurinck et al., 2003; Szczuka and Kaznowski, 2004).
One of these methods appeared interesting for the delineation of species, subspecies (Louws et al., 1998) or pathovars (Louws et al., 1994) for instance. BOX-PCR, independent from the other rep-PCR techniques, has revealed the possibility of delineating P. syringae genomospecies (Marques et al., 2000), as well as for typing Aeromonas spp. strains (Tacao et al., 2005) and for identification of races and biovars of Ralstonia solanacearum (Galal et al., 2003). The technique has also been used to investigate bacterial inoculum sources, as a tool for unequivocal identification of strains belonging to a unique pathovar or to define new species, as a part of a polyphasic approach (Catara et al., 2002).
Another type of rep-PCR is the amplification of genomic DNA located between Enterobacterial Repetitive Intergenic Consensus (ERIC) elements (Versalovic et al., 1991). ERIC elements are 126 bp in size and are distributed throughout extragenic regions of the genomes of many Gram-negative bacterial genera (Hulton et al., 1991; Versalovic et al., 1991, 1994). ERIC-PCR generates multiple distinct amplification products of sizes ranging from approximately 50 to 3000 bp. The unique locations of ERIC elements in bacterial genomes allows discrimination at genus, species and strain levels based on the electrophoretic pattern of amplification products (De Bruijn, 1992). Besides that, new DNA polymerases have recently been developed that arecapable of utilizing primers shorter than the standard minimum length of approx. 20 to 30 nt recognized by Taq DNA polymerase. Based on this, Isenbarger et al. (2008) developed a new PCR method, Miniprimer PCR, using 10-nucleotide ‘miniprimers’ and an engineered Taq polymerase for PCR amplification specific to 16S rRNA. This method has been applied to the exploration of new dimensions of microbial diversity and genotyping for identification purposes (Xu et al., 2010).
Traditional classifications based on phenotypic features do not always correlate with the molecular taxonomy (Woese, 1987). DNA-based techniques have provided new approaches to bacterial taxonomy and are leading the way forward in the bacterial diversity studies (Rossello-Mora and Amann, 2001).
The aim of this investigation was to evaluate and compare the new Miniprimer-PCR to the two DNA-based typing techniques (ERIC- and BOX-PCR) and the phenotypic characteristics for the purpose of assessing the diversity of microbial community isolated as collection of fluorescent pseudomonads.
MATERIALS AND METHODS
History of field plot and soil sample collection: The land at the Eugene Whelan Experimental Farm site was cleared of deciduous trees in the late 1800s or early 1900s and the long-term study started in 1959. Detailed descriptions of the experimental site and soils have been reported (Tan and Drury, 2002). Briefly, the soil at the experimental site, located at Woodslee, Ontario (42° 13' N, 82° 44' W), was a Brookston clay loam (Humic Gleysol) with mean particle distribution of 28.0% sand, 35% silt and 37% clay. Based on climate data over the period 2006 to 2008, the average annual temperature of this site is 10.6°C; the average maximum temperature when samples were collected (November 2008) is 24.9°C and the average minimum temperature is 14.4°C and average monthly precipitation of 139.6 mm (Environment Canada 2008: http://climate.weatheroffice.ec.gc.ca/climateData/monthlydata_e.hml). Although the complete records of agricultural management of the experimental site prior to 1954 are not available, it is known that maize (Zea mays L.), alfalfa (Medicago sativa L.) and red clover (Trifolium pratense L.) were grown between 1940 and 1954. In 1959, 12 plots (76.2 m long by 12.2 m wide), consisting of six cropping treatment plots with fertilizer and six without, were established. Two fertilized plots (alfalfa, S2 and corn, S5) and two unfertilized plots (alfalfa, S8 and corn, S7) from maize-oat-alfalfa-alfalfa rotation were used for the study. Each crop in the rotation was grown every year. Chemical fertilized treatments received annual applications of 16.8 kg N ha-1, 29.3 kg P ha-1 and 27.4 kg K ha-1, applied to all crops. Fertilized maize received an additional application of 112 kg N ha-l per year of side-dressed incorporated ammonium nitrate. A grass treatment (S11) was mowed to a height of 13-15 cm four to five times each year when the grass reached a height of 25 cm and the residues were left on the plots after mowing. Planting densities and management including herbicides to control weeds have been reported (Tan and Drury, 2002).
Surface soil samples (0 to 10 cm depth) were collected in November 2008 from the plot S2 under corn or alfalfa rotations. The field-moist soil samples were passed through a 2-mmsieve and stored at 4°C.
Isolation and preservation of isolates: Ten grams of various rhizospheric soil samples were suspended in 90 mL of 0.85% NaCl and the suspensions were vigorously mixed. Suspensions were serially diluted and plated on King’s B agar (Proteose peptone No. 3, 20 g L-1; K2HPO4, anhydrous, 1.5 g L-1; MgSO4.7H2O, 1.5 g L-1; 1.5% glycerol, 1.5% agar) in order to isolate fluorescent pseudomonads. Plates were inoculated for 48 h at 28°C. After incubation, colonies with different phenotypes were screened for fluorescense under UV light.
Make all isolates single colony on the pseudomonads selective plates. All bacterial strains were stored for preservation in Luria-Bertani medium (tryptone, 10 g L-1; yeast extract, 5 g L-1; NaCl, 10 g L-1) supplemented with 25% (w/v) glycerol.
Genomic DNA extraction: For all PCR analyses, DNA was extracted from bacterial strains using the wizard SV Genomic DNA purification system (Promega, Madison, WI, USA) and the concentration was determined using NanoDROP® ND-1000 Spectrophotometer (NanoDrop Technologies, Wilmington, DE, USA). Purified DNA was stored at -20°C.
BOX-PCR and ERIC-PCR: The amplication and electrophoresis of BOX- and ERIC-PCR were finished as described previously (Dawson et al., 2002). BOX-PCR fingerprinting was carried out using one primer of sequence 5’-CTACGGCAAGGCGACGCTGACG-3’ which corresponded to the bacterial repetitive BoxA subunit (Koeuth et al., 1995). ERIC-PCR was carried out using the primer sequences ERIC-1R, 5’-CACTTAGGGGTCCTCGAATGTA-3’ and ERIC-2, 5’-AAGTAAGTGACTGGGGTGAGCG-3’ to amplify the regions in the bacterial genome positioned between the ERIC sequences (Versalovic et al., 1991). Cycling conditions for ERIC-PCR were as follows: 95°C for 7 min and then 30 cycles of 94°C for 1 min, 51°C for 1 min and 65°C for 8 min, followed by a 16 min incubation at 65°C; after cooling to 4°C, the PCR products were stored at-20°C. Cycling conditions for BOX-PCR were the same except that the annealing temperature was 50°C. Amplified products were separated on 1.5% agarose gels in 0.53 Tris-borate-EDTA buffer for 5 to 6 h at 140 V and 10°C. All of the amplifications were performed at least twice in separate assays, to ensure the reproducibility of the patterns and only bands common to the replicate amplifications were scored.
Miniprimer-PCR: The miniprimer Uni-BacF-10(GATCCTGGCT) / Uni-BacR-10(GGACTACCAG) pair was designed from the Uni-BacF / Uni-BacR ‘long-primers’ set and synthesized by Invitrogen Inc. (Carlsbad, CA, USA). PCR conditions for miniprimers were optimized using 10 ng of template DNA. TitaniumTM Taq (Clontech, Inc, Palo Alto, CA, USA) DNA polymerases were used for PCR. The reaction mixture included 1x buffer, 200 μmol L-l dNTP, 400 nmol L-1 of each miniprimer and 0.75 U Taq DNA polymerase. Optimized PCR amplifications were performed in a Tprofessional thermal cycler (Biometra GmbH i.L., Göttingen, Germany) with an initial denaturation step at 95°C for 3 min, followed by 35 cycles of 95°C for 1 min, 44°C for 1 min and 72°C for 2 min and a final extension at 72°C for 8 min. Amplicons were separated by agarose gel electrophoresis (1.5%) at 100 V for 90 min. Each DNA sample replicated three times.
Gel image analysis: Gel images were acquired as a tif file using the AlphaEase Gel Imaging System (Alpha Innotech Corp., San Leandro, CA, USA) interfaced to a computer. Images were normalized, analysed and bands assigned and quantified after local background subtraction using GelCompar II ver 5.1 (Applied Maths, Kortrijk, Belgium). A 5% tolerance in band position was applied. Similarities among profiles were calculated based on Dice and Pearson coefficients and the clustering was performed with the unweighted pair-group method using arithmetic averages as implemented in GelCompar II (Applied Maths).
RESULTS
Phenotypic diversity: A total of 143 bacterial isolates were isolated from our soil sample. The strains were identified to belong to Pseudomonadaceae based on their ability to grow on the pseudomonads selective plates and on about 1 kb 16S rRNA sequences (data not shown), besides they all are fluorescent under UV except isolates 2-21 and 2-97-1. We also found the existence in many isolates of two different forms which can be distinguished on the basis of colony morphology and their moving ability on the solid media. One form I, is characterized by dull colonies with flat edges and yellow green pigmented appearance. This form also moves fast to be full of media plate. The other form II, is characterized by glossy, sticky colonies that tend to be slightly raised than the dull colonies. This form does not move so fast either. We labeled a letter “L” to distinguish the isolates showed like form II, however, there was also form III for some isolates colonies appearance between form I and II which we didn’t add the L. The phenptypes of the three variants were stable and maintained in subsequent cultures.
BOX-PCR: Of the 143 isolates, BOX-PCR produced 12 distinct patterns, with sizes ranging from 1,400 to 300 bp. The amplified banding profiles were clearly distinguishable, Visual inspection of the DNA fingerprints followed by analysis with GelCompar II ver 5.1 software enabled the identification of novel genotypes. A dendrogram (Fig. 1) that was calculated with Dice’s similarity coefficients with Unweighted Pair Group Method Clustering (UPGMA). The groups in BOX-PCR dendrogram can be classified into 2 big clusters: cluster I included 96 non L and 6 L isolates; cluster II included 27 L and 10 non L. To analysis further, at a similarity level of 40%, there were 15 unique genotypic groups and 4 groups consisting of single isolates (2-6, 2-41, 2-43-1, 2-90) in the 143 isolates with BOX-PCR fingerprinting.
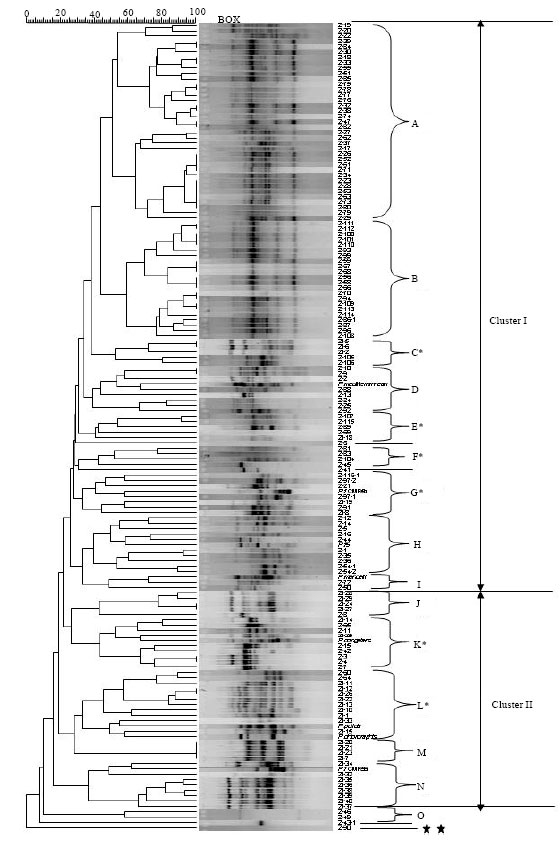 | |
| Fig. 1: | Dendrogram obtained by the cluster analysis of the BOX-PCR data. Similarities were calculated using the Dice coefficient and the data clustered using the unweighted pair group method of averages. Type strains are included. Group name with “*” means the group mixed L with non L. Pentagram stands for single-isolated group |
L-type isolates mixed with non L-type ones in 6 of the 15 groups. In group C, 2l-2, 2l-5 and 2l-6 mixed with 2-105 and 2-106; in group E, 2l-18 mixed with 2-107, 2-115, 2-89 and 2-69; in group G, 2l-19 and 2l-8 mixed with 2-116-1, 2-97-2, 2-21, 2-97-1 and 2-91; in group J, 2-8 mixed with 2l-28, 2l-29, 2l-24 and 2l-27; in group K, 2l-14 and 2l-25 mixed with 2-65, 2-11, 2-15, 2-42, 2-3, 2-4 and 2-7; in group L, 2-60 and 2-64 mixed with 9 L-type isolates.
ERIC-PCR: The patterns of ERIC-PCR for all investigated strains consisted of 12 fragments ranging in size from approximately 100 to 1400 bp. Nearly identical cluster patterns between BOX- and ERIC-PCR fingerprintings at the same similarity level. Nineteen groups were formed with ERIC-PCR fingerprinting (Fig. 2). Analogously, the groups in ERIC-PCR dendrogram can be classified into 2 big clusters too: cluster I included 93 non L and 3 L isolates 2l-1,2l-8 and 2l-18; cluster II included 30 L and 10 non L. L-type isolates mixed with non L-type ones in 6 of the 19 groups. In group A, 2l-1 mixed with 2-21 and 2-91; in group F, 2l-18 mixed with 24 non L-type isolates; in group I, 2l-8 mixed with 2-10, 2-9, 2-12, 2-5 and 2-89; in group L, 2-65, 2-2 and 2-6 mixed with 2l-38, 2l-39 and 2l-37; in group M, 2l-32 mixed with 2-15, 2-42, 2-3, 2-4, 2-13 and 2-7; in group S, 2l-6 and 2l-14 mixed with 2-8.
Miniprimer-PCR: The Miniprimer-PCR analysis revealed narrower band patterns than BOX- and ERIC-PCR, because the miniprimer was designed on the basis of 16S rRNA sequences. The band patterns generated by Miniprimer-PCR ranged from 1 to 9 in number and from about 100 to 1300 bp in size. The DNA fingerprints with the three primer sets, respectively were not exactly identical (Fig. 3). However, there were 16 groups and 4 single-isolate “groups” (2-44, 2-43-1, 2-49 and 2-72) with Miniprimer-PCR. Similarly, L-type isolates mixed with non L-type ones in 6 of the 19 groups. In group A, 2l-2, 2l-5 and 2l-6 mixed with 2-24, 2-25 and 2-81; in group E, 2l-21 and 2l-23 mixed with 11 non L-type isolates; in group F, 2-108 mixed with 21-15, 2l-8, 2l-32; in group G, 2l-25 mixed with 2-92, 2-65; in group I, 2l-7 and 2l-20 mixed with 2-15, 2-42, 2-3, 2-4, 2-13, 2-7,2-99, 2-2 and 2-8; in group L, 2-77 and 2-78 mixed with 7 L-type isolates.
Compare to the former two fingerprings, with Miniprimer-PCR the isolates were still classified into 9 clusters: cluster I included only isolates of the same non L type like the similar clusters in BOX- and ERIC-PCR, cluster II, III, V, VI and IX consisted mostly of non L isolates mixed with a few of L isolates while cluster IV, VII and VIII consisted mostly of L isolates mixed with a few of non L (colony morphology form III).
The combination of three methods: To gain information about the effect using 3 different methods to study microdiversity of the isolates, the Rep-PCR (BOX and ERIC) and Miniprimer PCR fingerprinting technique, were combined.
The 3 methods for all the 143 isolates yielded complex genomic fingerprints consisting of 9 to 12 amplified bands of varying intensity. A dendrogram that was calculated with Dice’s similarity coefficients with Unweighted Pair Group Method Clustering (UPGMA) for the combination of the 3 fingerprinting results segregated the 143 isolates into two big distinct clusters and 2 small clusters, even 3 single-isolate “clusters” (Fig. 4). Cluster I included 94 non L which can be divided into 5 unique genotypic groups and 1 single-isolate group: E was like A in BOX-PCR, E in ERIC-PCR while O in Miniprimer-PCR to compose mainly cluster I in the three fingerprintings, respectively. Cluster II included 29 L and 9 non L which can be divided into 3 subgroups: C was similarly with Kin BOX-PCR, M in ERIC-PCR while I in Miniprimer-PCR.
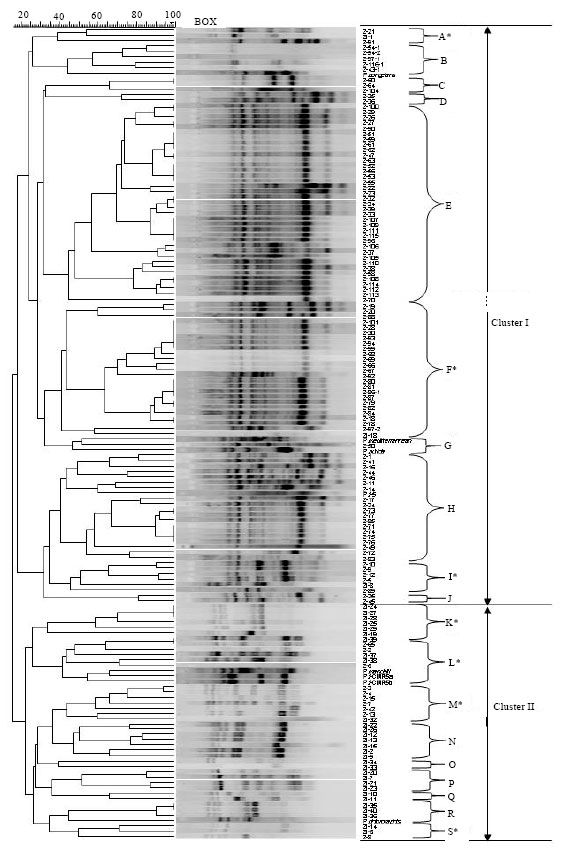 | |
| Fig. 2: | Dendrogram obtained by the cluster analysis of the ERIC-PCR data. Similarities were calculated using Dice coefficient and the data clustered using the unweighted pair group method of averages. Type strains are included. Group name with “*” means the group mixed L with non L |
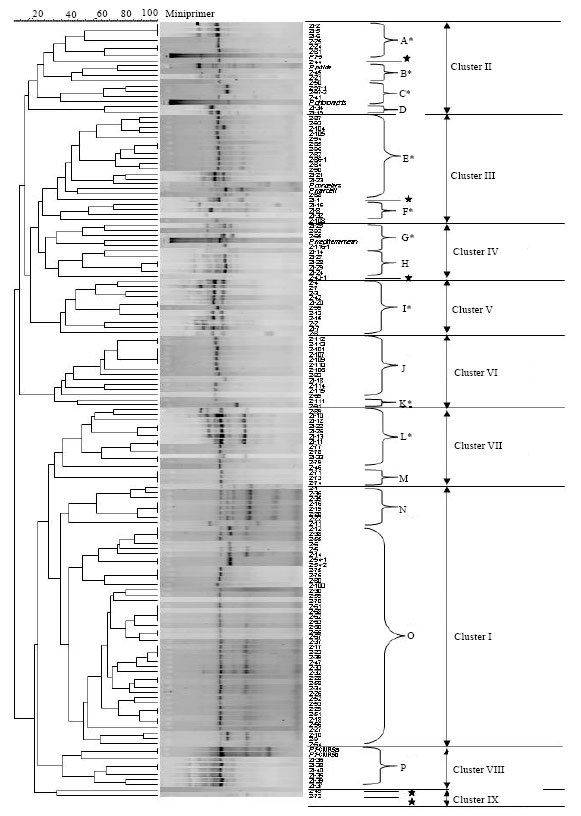 | |
| Fig. 3: | Dendrogram obtained by the cluster analysis of the Miniprimer-PCR data. Similarities were calculated using Dice coefficient and the data clustered using the unweighted pair group method of averages. Type strains are included. Group name with “*” means the group mixed L with non L. Pentagram stands for single-isolated group |
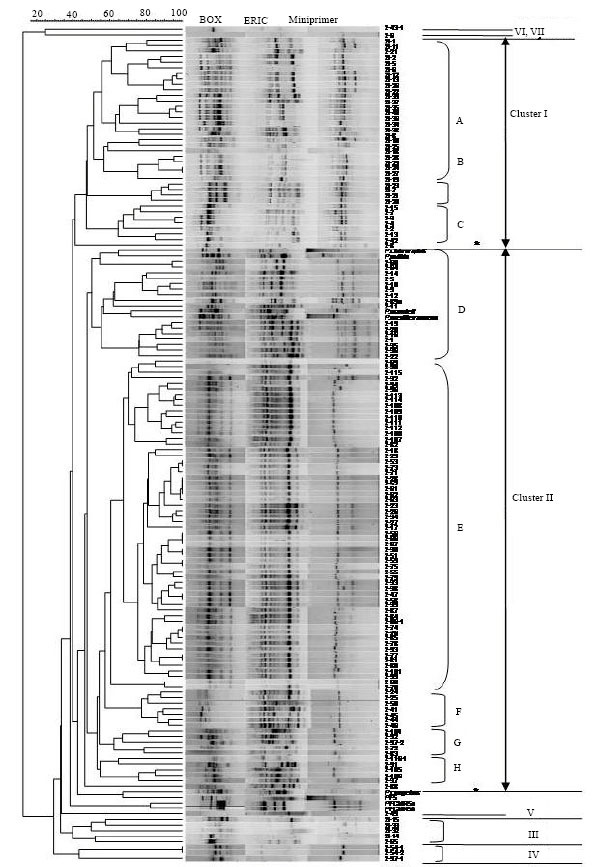 | |
| Fig. 4: | Cluster analysis of combined genomic fingerprint patterns of 143 isolates and 8 references |
Cluster III was consist of 2l-15, 2l-33, 2l-32 and 2l-14, 2-65 which were high-frequency isolates mixed with other isolates. Cluster IV was made up of 2-54-1, 2-54-2 and 2-97-1. In accordance with the result from phenotypic data, most of the L isolates (colony morphology form II) were grouped in the same cluster except some little differences.
DISCUSSION
In the present study, different PCR-based techniques were used to assess the genetic diversity of P. fluorescens isolated from maize-oat-alfalfa-alfalfa rotation plots in Canada. However, the study was mainly focused on the comparison among the three methods.
It is known that the total number of microorganisms in any ecological niche is up to 108-1012 per gram of soil (Alef and Nannipieri, 1995) and probably 103 different types are present (Torsvik and Ovreas, 2002).It is also recognized that the complexity and variability of microbial diversity has to be assessed at different biological levels (Garland and Mills, 1991). Microbial diversity can be measured by various methodologies such as direct and plate counts as well as fatty acid profiles and molecular methods (Nannipieri et al., 2003). Genetic diversity among microorganisms has been widely studied by PCR-based techniques which have opened avenues for the development of new and inventive molecular typing methods (Vaughan-Smith et al., 1995). Genotypic relationships among microorganisms have been determined by analyzing the genomic DNA with PCR-based methods (Pooler et al., 1996). Repetitive sequence-based PCR (Rep-PCR) has been used successfully to generate DNA fingerprints to distinguish between genetically unrelated isolates and closely related bacterial strains (Dombek et al., 2000). It involves the use of primers based on the short repetitive elements derived from highly conserved palindromic inverted repeat regions dispersed throughout the prokaryotic kingdom (Jersek et al., 1999; Laguerre et al., 1996). Amplification of the regions between adjacent repetitive extragenic elements gives strain-specific DNA fingerprints (Dombek et al., 2000). The inter-REP and inter-ERIC profiles are specific for bacterial strains within a species (Appuhamy et al., 1997; Versalovic et al., 1991).
Repetitive-DNA markers have been used with success in the identification of a large number of Gram-negative bacteria (Giselle and Lindstorm, 1994; Louws et al., 1994; Smith et al., 1995), including Escherichia coli (Seurinck et al., 2003), Salmonella (Tikoo et al., 2001), Aeromonas (Szczuka and Kaznowski, 2004), Burkholderia (Coenye et al., 2002), Vibri (Wong and Lin, 2001), among others. Rep-PCR provided the highest resolution of all the techniques investigated. This conclusion is in agreement with the conclusions from studies using 138 isolates of 2,4-diacetyl phloroglucinol producing fluorescent pseudomonads and 62 P. savastanoi isolates (Gardener et al., 2000; Marques et al., 2000). With both BOX and ERIC PCR, defined groups were formed among clinical isolates from different sources; these were separated from the environmental strains and subgroups within these clusters were resolved (Kim et al., 2003).
Miniprimer PCR is also as reproducible as other methods and provides a different portrait of microbial communities than is attained with current methods (Isenbarger et al., 2008). Also, miniprimer sequences provide a small amount of additional sequence data that may be helpful for making fine discriminations among closely related sequences. Xu et al. (2010) reported miniprimer PCR is a new, simple, rapid and reliable tool for genotyping the pathogen when compared to PCR-RFLP. Their first miniprimer PCR is sensitive enough to differentiate between subspecies of P. stewartii. This is significant given that distinguishing these subspecies is always problematic.
This article first reports the comparison among miniprimer PCR and rep-PCR (box- and eric-PCR). We also first use miniprimer PCR to study microbial community diversities of Pseudomonads. It is very important to increase our knowledge of the fluorescent pseudomonads which are organisms of great industrial, biotechnological and pathogenic importance, it is vital that an improved classification and a reliable identification scheme are developed. To achieve these objectives, more information is required on the diversity between and within species and techniques which can provide us with an abundance of this type of data, are essential.
All the three method can get some big groups which included most of non L type isolates when they were put together. In dendrogram obtained by Box-PCR, they were group A, B and H; in Eric-PCR, they were E, F and H; in Miniprimer PCR, they were J, N and O. Box- and Eric-PCR almost got the same result. There were distinctly two clusters distinguished from two mainly morphology variation. Besides that, these three methods all got 6 mixed groups. We found some of isolates were similar in three fingerprintings, such as 2-3, 2-4, 2-7, 2-8, 2-15, 2-42, 2l-8 and so on. In this case, the miniprimer specific for the 16S element was more effective than were the Box and Eric primer sets in obtaining fingerprints for grouping the strains effectively. With Miniprimer-PCR, we got more clusters than other 2 methods. In our study, the isolates’ colony morphology can be divided into 3 forms, i.e. 2 clearly different forms (I and II) and 1 ambiguous from (III) which can be called phase variation. Through former 2 analyses, isolates belonged to form I and III were grouped into cluster I while form II belonged to cluster II. Interestingly, we found form III mixed with form II in Miniprimer’s result. This suggests the amplimer with miniprimer could be significant in genotyping bacterial genomes, providing banding patterns that could enhance identification (Xu et al., 2010).
In conclusion, miniprimer may be useful to supplement traditional methods like Box- and Eric-PCR either to reduce or make apparent the biases that occur in analysis of environmental samples and allow a more accurate description of microbial communities. We concluded that a combination of Box-, Eric- and Miniprimer-PCR results is a rapid and reliable alternative for discrimination among P. fluorescens isolates, contributing to epidemiological studies.
ACKNOWLEDGMENTS
This research was supported by MOE-AAFC Joint PhD Research Program. Special thanks to Vic Bernyk and the field staff at the Eugene Whelan Experimental farm for maintaining this long-term site. We are grateful to Dr Guoyue Chen for reviewing this article.
REFERENCES
- Appuhamy, S., R. Parton, J.G. Coote and H.A. Gibbs, 1997. Genomic fingerprinting of Haemophilus somnus by a combination of PCR methods. J. Clin. Microbiol., 35: 288-291.
PubMed - Catara, V., L. Sutra, A. Morineau, W. Achouak, R. Christen and L. Gardan, 2002. Phenotypic and genomic evidence for the revision of Pseudomonas corrugata and proposal of Pseudomonas mediterranea sp. nov. Int. J. Syst. Evol. Microbiol., 52: 1749-1758.
CrossRefDirect Link - Clausen, G.B., L. Larsen, K. Johnsen, J.R. de Lipthay and J. Aamand, 2002. Quantification of the atrazine-degrading Pseudomonas sp. strain ADP in aquifer sediment by quantitative competitive polymerase chain reaction. FEMS Microbiol. Ecol., 41: 221-229.
CrossRefDirect Link - Coenye, T., T. Spilker, A. Martin and J.J. LiPuma, 2002. Comparative assessment of genotyping methods for epidemiologic study of Burkholderia cepacia Genomovar III. J. Clin. Microbiol., 40: 3300-3307.
PubMed - De Bruijn, F.J., 1992. Use of repetitive (repetitive extragenic palindromic and enterobacterial repetitive intergeneric consensus) sequences and the polymerase chain reaction to fingerprint the genomes of Rhizobium meliloti isolates and other soil bacteria. Applied Environ. Microbiol., 58: 2180-2187.
PubMed - Dombek, P.E., L.K. Johnson, S.T. Zimmerley and M.J. Sadowsky, 2000. Use of repetitive DNA sequences and the PCR to differentiate Escherichia coli isolates from human and animal sources. Applied Environ. Microbiol., 66: 2572-2577.
PubMedDirect Link - Galal, A.A., Y.E.I. Kehil, Y.H. El-Daoudi, Z.A. Shihata and M.F. Ouf, 2003. A comparative study on the identification of races and biovars of some Egyptian isolates of Ralstonia solanacearum. Egypt. J. Phytopathol., 31: 103-117.
Direct Link - Gardener, B.B.M., K.L. Schroeder, S.E. Kalloger, J.M. Raaijmakers, L.S. Thomashow and D.M. Weller, 2000. Genotypic and phenotypic diversity of phlD-containing Pseudomonas strains isolated from the rhizosphere of wheat. Applied Environ. Microbiol., 66: 1939-1946.
PubMed - Giselle, N. and K. Lindstorm, 1994. Use of repetitive sequences and the polymerase chain reaction to fingerprint the genomic DNA of Rhizobium galegae strains and to identify the DNA obtained by sonicating the liquid cultures and root nodules. Syst. Applied Microbiol., 17: 265-273.
Direct Link - Hulton, C.S.J., C.F. Higgins and P.M. Sharp, 1991. ERIC sequences: A novel family of repetitive elements in the genomes of Escherichia coli, Salmonella typhimurium and other enterobacteria. Mol. Microbiol., 5: 825-834.
CrossRefPubMedDirect Link - Isenbarger, T.A., M. Finney, C. Rios-Velazquez, J. Handelsman and G. Ruvkun, 2008. Miniprimer PCR, a new lens for viewing the microbial world. Appl. Environ. Microbiol., 74: 840-849.
CrossRefDirect Link - Jersek, B., P. Gilot, M. Gubina, N. Klun and J. Mehle et al., 1999. Typing of Listeria monocytogenes strains based on repetitive element sequences-based PCR. J. Clin. Microbiol., 37: 103-109.
PubMed - Garland, J.L. and A.L. Mills, 1991. Classification and characterization of Heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization. Applied Environ. J. Microbiol., 57: 2351-2359.
PubMed - Koeuth, T., J. Versalovic and J.R. Lupski, 1995. Differential subsequence conservation of interspersed repetitive Streptococcus pneumoniae box elements in diverse bacteria. Genome Res., 5: 408-418.
PubMedDirect Link - Laguerre, G., P. Mavingui, M.R. Allard, M.P. Charnay and P. Louvrier et al., 1996. Typing of rhizobia by PCR and PCR-restriction fragment length polymorphism analysis of chromosomal and symbiotic gene regions: application to Rhizobium leguminosarum and its different biovars. Applied Environ. Microbiol., 62: 2029-2036.
PubMed - Louws, F.J., D.W. Fulbright, C.T. Stephens and F.J. de Bruijn, 1994. Specific genomic fingerprints of phytopathogenic Xanthomonas and Pseudomonas pathovars and strains generated with repetitive sequences and PCR. Appl. Environ. Microbiol., 60: 2286-2295.
PubMedDirect Link - Louws, F.J., J. Bell, C.M. Medina-Mora, C.D. Smart and D. Opgenorth et al., 1998. Rep-PCR-mediated genomic fingerprinting: A rapid and effective method to identify Clavibacter michiganensis. Phytopathology, 88: 862-868.
PubMedDirect Link - Marques, A.S.A., R. Corbiere, L. Gardan, C. Tourte, C. Manceau, J.D. Taylor and R. Samson, 2000. Multiphasic approach for the identification of the different classification levels of Pseudomonas savastanoi pv. phaseolicola. Eur. J. Plant Pathol., 106: 715-734.
CrossRefDirect Link - Olive, D.M. and P. Bean, 1999. Principles and applications of methods for DNA-based typing of microbial organisms. J. Clin. Microbiol., 37: 1661-1669.
PubMedDirect Link - Patten, C.L. and B.R. Glick, 2002. Role of Pseudomonas putida indoleacetic acid in development of the host plant root system. Applied Environ. Microbiol., 68: 3795-3801.
CrossRefPubMedDirect Link - Nannipieri, P., J. Ascher, M.T. Ceccherini, L. Landi, G. Pietramellara and G. Renella, 2003. Microbial diversity and soil functions. Eur. J. Soil Sci., 54: 655-670.
CrossRefDirect Link - Pooler, M.R., D.F. Ritchie and J. S. Hartung, 1996. Genetic relationships among strains of Xanthomonas fragariae based on random amplified polymorphic DNA PCR, repetitive extragenic palindromic PCR and enterobacterial repetitive intergenic consensus PCR data and generation of multiplexed PCR primers useful for identification of this phytopathogen. Applied Environ. Microbiol., 62: 3121-3127.
PubMed - Rossello-Mora, R. and R. Amann, 2001. The species concept for prokaryotes. FEMS Microbiol. Rev., 25: 39-67.
CrossRefPubMedDirect Link - Samson, R., H. Shafik, A. Benjamo and L. Gardan, 1998. Description of the bacterium causing blight of leek as Pseudomonas syringae pv. porri (pv. nov.). Phytopathology, 88: 844-850.
PubMedDirect Link - Seurinck, S., W. Verstraete and S.D. Siciliano, 2003. Use of 16S-23S rRNA intergenic spacer region PCR and repetitive extragenic palindromic PCR analyses of Escherichia coli isolates to identify nonpoint fecal sources. Applied Environ. Microbiol., 69: 4942-4950.
PubMed - Smith, J.J., L.C. Offord, M. Holderness and G.S. Saddler, 1995. Genetic diversity of Burkholderia solanacearum (synonym Pseudomonas solanacearum) race 3 in Kenya. Applied Environ. Microbiol., 61: 4263-4268.
PubMedDirect Link - Szczuka, E. and A.Kaznowski, 2004. Typing of clinical and environmental Aeromonas sp. strains by random amplified polymorphic DNA PCR, repetitive extragenic palindromic PCR and enterobacterial repetitive intergenic consensus sequence PCR. J. Clin. Microbiol., 42: 220-228.
CrossRefDirect Link - Tacao, M., A. Alves, M.J. Saavedra and A. Correia, 2005. BOX-PCR is an adequate tool for typing Aeromonas sp. Antoine Van Leeuwenhoek, 88: 173-179.
CrossRefDirect Link - Tikoo, A., A.K. Tripathi, S.C. Verma, N. Agrawal and G. Nath, 2001. Application of PCR fingerprinting techniques for identification and discrimination of Salmonella isolates. Curr. Sci., 80: 1049-1052.
Direct Link - Vaughan-Smith, H.C., K.S. Sriprakash, J.D. Mathews and D.J. Kemp, 1995. Long-range PCR ribotyping of nontypeable Haemophilus influenzae. J. Clin. Microbiol., 33: 1192-1195.
PubMedDirect Link - Van Loon, L.C., P.A.H.M. Bakker and C.M.J. Pieterse, 1998. Systemic resistance induced by rhizosphere bacteria. Annu. Rev. Phytopathol., 36: 453-483.
CrossRefPubMedDirect Link - Versalovic, J., T. Koeuth and R. Lupski, 1991. Distribution of repetitive DNA sequences in eubacteria and application to finerpriting of bacterial enomes. Nucl. Acids Res., 19: 6823-6831.
CrossRefPubMedDirect Link - Torsvik, V. and L. Ovreas, 2002. Microbial diversity and function in soil: From genes to ecosystems. Curr. Opin. Microbiol., 5: 240-245.
PubMed - Wong, H.C. and C.H. Lin, 2001. Evaluation of typing of Vibrio parahaemolyticus by three PCR methods using specific primers. J. Clin. Microbiol., 39: 4233-4240.
CrossRefPubMedDirect Link - Kim, W., M.O. Song, W. Song, K.J. Kim, S.I. Chung, C.S. Choi and Y.H. Park, 2003. Comparison of 16S rDNA analysis and rep-PCR genomic fingerprinting for molecular identification of Yersinia pseudotuberculosis. Antonie van Leeuwenhoek, 83: 125-133.
CrossRef - Shaukat, K., S. Affrasayab and S. Hasnain, 2006. Growth responses of Helianthus annus to plant growth promoting rhizobacteria used as a biofertilizer. J. Agric. Res., 1: 573-581.
CrossRefDirect Link - Noura, K.M. Salih, N.H. Jusuf, A.A. Hamid and W.M.W. Yusoff, 2009. High prevalence of Pseudomonas species in soil samples from Ternate Island-Indonesia. Pak. J. Biol. Sci., 12: 1036-1040.
CrossRefPubMedDirect Link - Soleimani, M.J., M. Shamsbakhsh, M. Taghavi and S. Kazemi, 2005. ogical control of stem and root-rot of wheat caused by Bipolaris spp. by using antagonistic bacteria, fluorescent Pseudomonads and Bacillus spp. J. Biological Sci., 5: 347-353.
CrossRefDirect Link - Tan, C.S., C.F. Drury, W.D. Reynolds, P.H. Groenevelt and H. Dadfar, 2002. Water and nitrate loss through tiles under a clay loam soil in Ontario after 42 years of consistent fertilization and crop rotation. Agric. Ecosyst. Environ., 93: 121-130.
CrossRef - Siddiqui, Z.A. and U. Shakeel, 2006. Use of fluorescent pseudomonads isolates for the biocontrol of wilt disease complex of pigeonpea in green house assay and under pot condition. Plant Pathol. J., 5: 99-105.
CrossRefDirect Link - Nezarat, S. and A. Gholami, 2009. Screening plant growth promoting rhizobacteria for improving seed germination, seedling growth and yield of maize. Pak. J. Biol. Sci., 12: 26-32.
CrossRefPubMedDirect Link