
M. G. AL-Saghir
Department of Environmental and Plant Biology, Ohio University Zanesville, Ohio, 43701, USA
Abdel-Salam G. Abdel Salam
Department of Statistics, Oklahoma University, Stillwater, 74078, USA
Journal of Plant Sciences
Year: 2011 | Volume: 6 | Issue: 1 | Page No.: 36-42







ABSTRACT
The objectives of this study were to evaluate the genetic diversity of the North American soybean (Glycine max L.) accessions using Random Amplified Polymorphic DNA (RAPD) molecular marker; and to evaluate RAPD markers to be used in soybean as genetic markers and improve such techniques as suitable strategies for soybean germplasm characterization. Twenty soybean accessions were included in this study and were subjected to RAPD molecular markers analysis. Twenty-seven RAPD primers produced 210 amplification products of which 78 (27.3%) were polymorphic. In conclusion, this study reported a successful fingerprinting of G. max accessions using RAPD markers and demonstrated the usefulness of these markers in estimating the extent of genetic variation in Soybean germplasm.
PDF Abstract XML References Citation
Received: October 25, 2010;
Accepted: December 02, 2010;
Published: March 25, 2011
How to cite this article
M. G. AL-Saghir and Abdel-Salam G. Abdel Salam, 2011. Genetic Diversity of North American Soybean (Glycine max L.) Cultivars as Revealed by RAPD Markers. Journal of Plant Sciences, 6: 36-42.
DOI: 10.3923/jps.2011.36.42
URL: https://scialert.net/abstract/?doi=jps.2011.36.42
DOI: 10.3923/jps.2011.36.42
URL: https://scialert.net/abstract/?doi=jps.2011.36.42
INTRODUCTION
Soybean is one of the important oil and protein crops in the world. Soybean breeders in the USA have successfully developed hundreds of improved cultivars (Fehr, 1984; Sneller, 1994; Ude et al., 2003). Knowledge of diversity patterns will allow breeders to better understand the evolutionary relationships among accessions, to sample germplasm in a more systematic fashion and to develop strategies to incorporate useful diversity in their breeding programs (Bretting and Widrlechner, 1995).
Different approaches were used to assay genetic diversity in crop plants including morphological traits and isozyme electrophoresis, however, these techniques are insufficient to serve as accurate markers due to environmental influences on morphological traits and insufficient polymoprhism produced among closely related genotypes (Li and Nelson, 2001; Matus and Hayes, 2002).
Certain properties are desirable for a molecular marker such as highly polymorphic behavior, co dominant inheritance and frequent occurrence in the genome, even distribution throughout the genome, selectively neutral behavior, easy access, easy and fast assay and high reproducibility (Weining and Langridge, 1991; Weining and Henry, 1995). Examples of such DNA molecular markers are: Random Amplified Polymorphic DNA (RAPDs) (Welsh and McCelland, 1990; Williams et al., 1990). RAPD markers had proved to be good genetic markers to assay and evaluate the genetic diversity between and within the same species, populations and individuals (Weising et al., 1995; Warburton and Bliss, 1996). RAPD marker depends on the amplification of DNA sequence by polymerase chain reaction using only a single primer of arbitrary nucleotide sequence. The technique has proved to be fast and simple needs small quantities of template DNA, beside its ability to detect relatively small amounts of genetic variation (Williams et al., 1990; Warburton and Bliss, 1996).
A series of studies have been conducted on soybean using isozymes (Griffin and Palmer, 1995; Li and Nelson, 2001; Malik et al., 2009), Microsatellites (Narvel et al., 2000; Li and Nelson, 2001; Li et al., 2010) and AFLPs (Ude et al., 2003).
The objectives of this study were to characterize the molecular diversity of the soybean accessions by analyzing the DNA amplification products using RAPD molecular markers; evaluate RAPD method to be used in soybean as genetic markers and improve such techniques as suitable strategies for soybean germplasm characterization.
MATERIALS AND METHODS
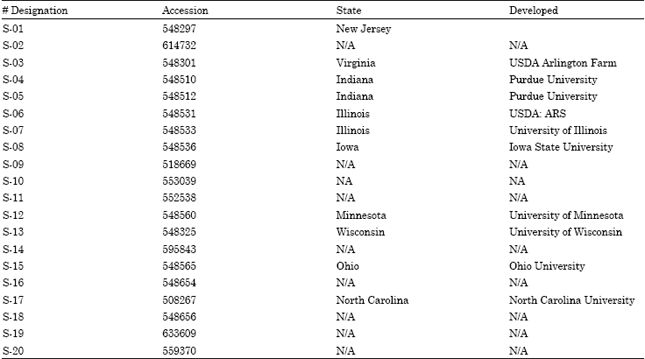
Plant material and DNA extraction: This study was conducted between 04/2009 and 04/2010 at Ohio University Zanesville. Seeds represented 20 genotypes of soybean were obtained from US department of Agriculture (USDA-ARS germplasm) (Table 1).
DNA was extracted from the young leaves of green house planted seedlings (4 week old) using DNeasy Plant Mini Kit (QIAGEN Inc., Valenica, CA, USA).
RAPD amplification: Thirty random primers (10-mer) from two kits (A and B) (Eurfins MWG Operon, Huntsville, AL, USA) of arbitrary sequence were used in this study (Table 2).
RAPD reactions were done in a total volume of 20 μL containing 20-60 ng of template DNA, 60 ng of each primer, 10 mL of Taq PCR Master Mix (QIAGEN Inc., Valencia, CA, USA) and 8 mL of PCR water (QIAGEN Inc., Valencia, CA, USA).
| Table 1: | USDA Glycine max germplasm collection used in this study |
 | |
| Table 2: | Random primers used to screen soybean germplasm for RAPDs |
 | |
Template DNA was initially denatured at 94°C for 2 min followed by 35 cycles for 1 min at 94°C, at annealing temperature of 37°C for 1 min and at 72°C for 2 min as an extension step. The final extension step was done for 8 min at 72°C and the reactions were kept at soak file at 4°C.
The RAPD-PCR amplified products were analyzed by gel electrophoresis in 1.5% ultrapure agarose in 1X TBE buffer stained with ethidium bromide (0.5 μg mL) at 100 volts using horizontal gel electrophoresis apparatus (Sigma Chemical Co. Louis, MO, USA). The amplified products were visualized under UV light and photographed with digital Olympus C-7070 camera (Olympus imaging America Inc., Melville, NY, USA). One kilobite ladder was used as a DNA standard to estimate the molecular weights of the amplified products.
Statistical analysis: For each individual RAPD primer, PCR amplified products were designated. Data were scored on the basis of the presence or absence of the amplified products. If the product is present in a genotype, it was scored as 1, if absent, it was designated as 0. Using the SAS statistical computer program SAS 9.2 (SAS Institute Inc., 2000-2009), genetic similarities between the soybean genotypes were calculated using the simple matching coefficient and clustered by unweighted pairs group method with arithmetic average (UPGMA) based on the average linkage method of calculating distance between clusters in hierarchical cluster analysis to construct dendrograms (Rohlf, 1993).
RESULTS AND DISCUSSION
Out of a total of 210 amplification products (0.2 to 4 kilo base pairs) using twenty-seven primers, 78 (27.3%) were polymorphic and 132 products were shared among all genotypes (not polymorphic). The twenty-four primers (OPA-02, OPA-03, OPA-04, OPA-05, OPA-07, OPA-09, OPA-10, OPA-11, OPA-12, OPA-13, OPA-14, OPA-15, OPA-16, OPA-17, OPA-18, OPA-19, OPA-20, OPB-03, OPB-04, OPB-05, OPB-06, OPB-07, OPB-10, OPB-11, OPB-12) produced different banding patterns for all genotypes. While sixteen primers (OPA-01, OPA-06, OPA-08, OPB-01, OPB-02, OPB-08, OPB-09, OPB-13, OPB-14, OPB-15, OPB-16, OPB-17, OPB-18, OPB-19 and OPB-20) detected no polymorphism although they did successfully amplify a range of monomorphic bands.
Two major clusters resulted: One cluster included by itself (S1 to S5) and the second cluster included the rest of the accessions (S6 to S20). This second cluster was subdivided into two sub clusters (sub cluster included S6 to S10 and the other sub cluster included S11 to S20) (Fig. 1). The genetic distance between first and the second cluster was 4.1.
Cluster analysis of each RAPD profiles (Fig. 1) showed that a significant genetic variation was detected in soybean accessions in particular samples S1 to S10 which could reflect different genetic background. Moreover, cluster analysis showed that samples S11 to S20 were genetically close (they showed 96% similarity).
The dendrogram (Fig. 1) showed that the RAPD methodology was sensitive enough to detect low levels of variation in soybean accessions.
The current study aimed on using the PCR-based protocols to assess genetic variability and to fingerprint genotypes of the North American soybean. Random amplified polymorphic DNA (RAPD) had been used effectively to assess the amount of genetic diversity in germplasm collections. Using wheat, barley, rye and wheat-barley addition lines, Weining and Langridge (1991) detected polymorphism using conserved, semi random and random primers. With different combinations of primers, they were able to detect both inter and intra specific diversity. In this study it was possible to show that the amplification products from 27 random primers (RAPD assay) were sufficient to discriminate among individual genotypes of soybean.
Few genetic studies have been conducted on North American soybean cultivars. In recent studies, fingerprints based on different markers were compared using genotypes from different species. There were both agreements and disagreements in findings based on different markers and species: The study of Brassica oleracea by Lanner-Herrera et al. (1996), a moderate spearman’s rank correlation (r = 0.38) between RAPD and isozyme distances was found.
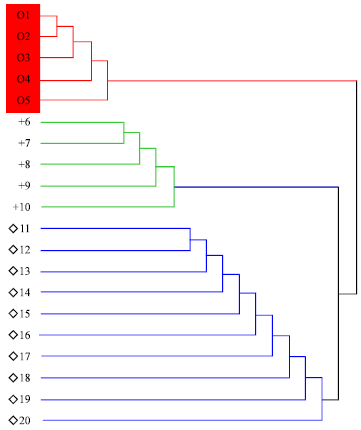 | |
| Fig. 1: | Dendrogram derived from the UPGMA procedure using genetic distances generated from the AMOVA program depicting the relationships among soybean accessions. Genetic distances are estimated from RAPD markers. Red = First cluster; Green and Blue collectively = Second cluster |
Russell et al. (1997) compared the levels of genetic variation among barley accessions revealed by RFLP, AFLP, SSRand RAPD and reported that when the spearman’s rank correlation was used, the correlation between SSR and RAPD was 0.235, the highest correlation was found between RFLP and AFLP (0.708).
Because of the low number of the genetic studies and the poor resolution of some employed molecular markers in these studies, more studies like the current study are urgently needed in order to evaluate and estimate the genetic diversity in the North American soybean germplasm.
In conclusion, this study reported a successful fingerprinting of G. max accessions using RAPD and demonstrated the usefulness of these markers in estimating the extent of genetic variation in soybean germplasm. The current results are supported by many recent molecular studies (Thompson and Nelson, 1998a, b; Thompson et al., 1998; Li and Nelson, 2001; Ude et al., 2003).
The use of RAPD markers in Glycine must be further continued in order to drive specific linkage between RAPD markers and genes controlling agronomically important characters. These diagnostic molecular tools will greatly assist in the identification of new and different sources of diversity which may help breeders to decide what genotypes to cross for making new genetic combinations and to determine which genetic resources should be retained in a collection in order to conserve maximum genetic diversity in the gene bank.
ACKNOWLEDGMENT
The author is grateful for US Department of Agriculture for providing the soybean cultivars and for Ohio University Zanesville for funding this project.
REFERENCES
- Malik, M.F.A., S. Afsari, Qureshi, M. Ashraf, M.R. Khan and A. Javed, 2009. Evaluation of genetic diversity in soybean (Glycine max) lines using seed protein electrophoresis. Aust. J. Crop Sci., 3: 107-112.
Direct Link - Griffin, J.D. and R.G. Palmer, 1995. Variability of thirteen isozyme loci in the USDA soybean germplasm collections. Crop Sci., 35: 897-904.
Direct Link - Lanner-Herrera, C., M. Gustagsson, A.S. Falt and T. Bryngelsson, 1996. Diversity in natural populations of wild Brassica olereacea as estimated by isozyme and RAPD analysis. Genet. Resour. Crop Evol., 43: 13-23.
CrossRef - Li, Z. and R.L. Nelson, 2001. Genetic diversity among soybean accessions from three countries measured by RAPDs. Crop Sci., 41: 1337-1347.
Direct Link - Li, Y.G., W. Li, C. Zhang, L. Yang, R.Z. Chang, B.S. Gaut and L.J. Qiu, 2010. Genetic diversity in domesticated soybean (Glycine max) and its wild progenitor (Glycine soja) for simple sequence repeat and single-nucleotide polymorphism loci. New Phytol., 188: 242-253.
CrossRef - Matus, I.A. and P.M. Hayes, 2002. Genetic diversity in three groups of barley germplasm assessed by simple sequence repeats. Genome, 45: 1095-1106.
CrossRefDirect Link - Narvel, J.M., W.R. Fehr, W. Chu, D. Grant and R.C. Shoemaker, 2000. Simple sequence repeat diversity among soybean plant introductions and elite genotypes. Crop Sci., 40: 1452-1458.
Direct Link - Russell, J.R., J.D. Fuller, M. Macaulay, B.G. Hatz, A. Jahoor, W. Powell and R. Waugh, 1997. Direct comparison of levels of genetic variation among barley accessions detected by RFLPs, AFLPs, SSRs and RAPDs. Theor. Applied Genet., 95: 714-722.
Direct Link - Weining, S. and R.J. Henry, 1995. Molecular analysis of the DNA polymorphism of wild barley (Hordeum spontaneum) germplasm using the polymerase chain reaction. Genet. Res. Crop Evol., 42: 237-281.
CrossRef - Thompson, J.A. and R.L. Nelson, 1998. Core set of primers to evaluate genetic diversity in soybean. Crop Sci., 38: 1356-1362.
Direct Link - Thompson, J.A. and R.L. Nelson, 1998. Utilization of diverse germplasm for soybean yield improvement. Crop Sci., 38: 1362-1368.
Direct Link - Thompson, J.A., R.L. Nelson and L.O. Vodkin, 1998. Identification of diverse soybean germplasm using RAPD markers. Crop Sci., 38: 1348-1355.
CrossRefDirect Link - Ude, G.N., W.J. Kenworthy, J.M. Costa, P.B. Cregan and J. Alvernaz, 2003. Genetic diversity of soybean cultivars from China, Japan, North America and North American ancestral lines determined by amplified fragment length polymorphism. Crop Sci., 43: 1858-1867.
CrossRefDirect Link - Warburton, M.L. and F.A. Bliss, 1996. Genetic diversity in peach (Prunus persica L. Batch) revealed by randomly amplified polymorphic DNA (RAPD) markers and compared to inbreeding coefficients. J. Am. Soc. Hortic. Sci., 121: 1012-1019.
Direct Link - Weining, S. and P. Langridge, 1991. Identification and mapping of polymorphisms in cereales based on the polymerase chain reaction. Teor. Applied Gent., 82: 209-216.
Direct Link - Welsh, J. and M. McClelland, 1990. Fingerprinting genomes using PCR with arbitrary primers. Nucleic Acids Res., 18: 7213-7218.
CrossRefPubMedDirect Link - Williams, J.G.K., A.R. Kubelik, K.J. Livak, J.A. Rafalski and S.V. Tingey, 1990. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res., 18: 6531-6535.
CrossRefPubMedDirect Link