
Y. X. Liu
Key Laboratory of Crop Genetic Resources and Improvement in Southwest China, Ministry of Education, Sichuan Agricultural University, Yaan, Sichuan 625014, China
W. Li
College of Agronomy, Sichuan Agricultural University, Yaan, Sichuan 625014, China
H. P. Chen
College of Biology and Science, Sichuan Agricultural University, Yaan, Sichuan 625014, China
Y. M. Wei
Key Laboratory of Crop Genetic Resources and Improvement in Southwest China, Ministry of Education, Sichuan Agricultural University, Yaan, Sichuan 625014, China
G. Y. Chen
Key Laboratory of Crop Genetic Resources and Improvement in Southwest China, Ministry of Education, Sichuan Agricultural University, Yaan, Sichuan 625014, China
Y .L. Lu
Key Laboratory of Crop Genetic Resources and Improvement in Southwest China, Ministry of Education, Sichuan Agricultural University, Yaan, Sichuan 625014, China
Y .L. Zheng
Key Laboratory of Crop Genetic Resources and Improvement in Southwest China, Ministry of Education, Sichuan Agricultural University, Yaan, Sichuan 625014, China
Journal of Plant Sciences
Year: 2008 | Volume: 3 | Issue: 4 | Page No.: 266-276







ABSTRACT
Genetic variation of high-molecular-weight glutenin subunits (HMW-GS), low-molecular-weight glutenin subunit (LMW-GS) genes, waxy locus (Wx) and quality characters, including protein content, wet gluten content, sodium dodecyl sulfate (SDS) sedimentation value and dough rheologic properties, were investigated in sixty-seven Sichuan landraces wheat in China. The relationship between these genetic variation and quality characters were also estimated. Alleles Glu-A1c (98.5%), Glu-B1b (98.5%) and Glu-D1a (100%), were dominant alleles at Glu-A1, Glu-B1 and Glu-D1 locus, respectively. Five, three and five types of different LMW-GS allele compositions were also identified and a (frequencies 74.5%), f (61.2%) and i (55.2%) were the dominant types at the Glu-A3, Glu-B3 and Glu-D3 loci, respectively. Significant difference (p<0.05) between types a and d at Glu-A3 locus were found in protein content, sedimentation value, wet gluten content and stability time, difference (p<0.01) between types g and h at Glu-B3 locus in sedimentation value, development time and breakdown time and difference (p<0.01) between types k and m on wet gluten content. All of the landraces carried the wild type Wx-A1a and Wx-B1a alleles. These information could be useful to marker-assisted select for different end-use quality wheat.
PDF Abstract XML References Citation
How to cite this article
Y. X. Liu, W. Li, H. P. Chen, Y. M. Wei, G. Y. Chen, Y .L. Lu and Y .L. Zheng, 2008. Variation for Glutenin and Waxy Alleles and Their Effect on Quality Properties in Sichuan Wheat Landraces. Journal of Plant Sciences, 3: 266-276.
DOI: 10.3923/jps.2008.266.276
URL: https://scialert.net/abstract/?doi=jps.2008.266.276
DOI: 10.3923/jps.2008.266.276
URL: https://scialert.net/abstract/?doi=jps.2008.266.276
INTRODUCTION
Modern wheat (Triticum aestivum) breeding programs place high priority on selecting lines with favorable qualities for the intended end-use. Numerous genetic and biochemical studies have focused on the characterization of alleles of wheat quality genes and their influence on end-use quality (D’Ovidio and Masci, 2004; Gianibelli et al., 2001; Graybosch, 1992). Among the various quality components, glutenin subunit composition and waxy alleles are of particular interest to wheat breeders.
Glutenin proteins are the major factors responsible for the unique viscoelastic dough characteristics of wheat flour, which determine mixing and bread baking performance (Gianibelli et al., 2001). These proteins can be separated into two groups by sodium dodecylsulfate polyacrylamidegel electrophoresis (SDS-PAGE): the high molecular weight glutenin (HMW-GS) and low molecular weight glutenin subunits (LMW-GS). The genes encoding HMW-GS reside at the Glu-1 loci (Glu-A1, Glu-B1 and Glu-D1), located on the long arms of group 1 homologous chromosomes (Lawrence and Shepherd, 1981; Payne et al., 1980). Each Glu-1 locus comprises two tightly linked genes (designated x and y), which exhibit multiple alleles among different varieties. Payne (1983) and Lawrence GJ (1983) proposed a nomenclature system to facilitate the genotyping of HMW-GS. Different HMW-GS alleles and allelic frequencies have been reported for various sets of wheat germplasm since then (Branlard et al., 2003; Graybosch, 1992; Payne and Lawrence, 1983; Redaelli et al., 1997; Wei et al., 2000). Relationships have also been established between Glu-1 alleles, especially at the Glu-D1 locus and bread making quality (Gupta et al., 1994; Payne et al., 1979, 1981; Pane, 1987). Payne (1987) assigned scores to each identified Glu-1 allele, which made it possible to predict the approximate bread making quality of wheat cultivars.
The LMW-GS are mostly encoded by the Glu-3 loci (Glu-A3, Glu-B3 and Glu-D3) on the short arms of group 1 homologous chromosomes (Singh and Shepherd, 1988). The estimates of gene copy number varied from 10-15 (Harberd et al., 1985) to 35-40 (Cassidy et al., 1998; Sabelli and Shewry, 1991). Over the past few years, more efforts have been focused on analysis at the molecular level. To date, more than 90 cDNA and genomic DNA clones of LMW-GS gene have been reported and the gene structure has been well established. On the basis of the sequence data, Van Campenhout et al. (1995) designed several primer sets specific for each of the Glu-3 loci and successfully determined the chromosomal locations of five LMW-GS genes. D'Ovidio et al. (1997) and Zhang et al. (2004) developed primer sets specific for Glu-B3 locus and Glu-A3 alleles, respectively. Ikeda et al. (2002) isolated several LMW-GS gene clones from a soft wheat cultivar and classified them into 12 groups based on the N- and C-terminal sequences. And recently, Long et al. (2005) analyzed 69 known LMW-GS gene from Triticum, classified them into nine groups and developed group-specific primers to detected each group of LMW-GS genes.
Wheat waxy proteins are Granule-Bound, Starch Synthase I (GBSSI) enzymes responsible for amylose biosynthesis (Nakamura et al., 1993). Wheat cultivars that carry mutant (null) waxy alleles lack one or more waxy proteins and produce flour with special starch qualities important for white salted noodle making (McLauchlan et al., 2001; Zhao et al., 1998). Positive effects on bread making quality due to a null waxy allele have also been reported (Martin et al., 2004). Waxy genes are situated on chromosomes 7A (Wx-A1), 7D (Wx-D1) and 4A (Wx-B1) and these genes have been cloned and sequenced (Murai et al., 1999). Codominant markers are available for identifying the normal and null alleles from all three waxy loci by one simple Polymerase Chain Reaction (PCR) assay (McLauchlan et al., 2001), which provides an advantage over the relatively laborious protein analysis method for waxy alleles.
The objectives of this study for Sichuan wheat landraces in China, were to determine the HMW-GS composition, to document for the first time the LMW-GS genes composition, to determine waxy allele composition with the PCR method and to illustrate relationships between these alleles and quality characteristics.
MATERIALS AND METHODS
Plant Material
A total of 67 accessions of wheat (Triticum aestivum ssp. aestivum) landraces were used in this study (Table 1). All these landraces were collected in Sichuan province and maintained at the Triticeae Research Institute of Sichuan Agricultural University. Each landrace was tested on three lines at normal seed density, with classic nitrogen and mineral supply under full fungicide and herbicide protection from October 2006 to May 2007. Grain harvested in bulk, for each accessions, was used for quality testing in June 2007.
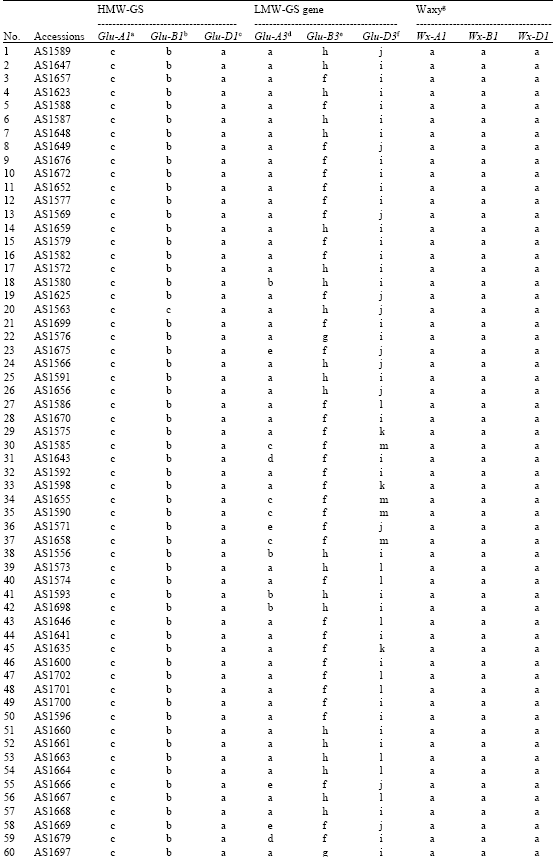
| Table 1: | HMW-GS, LMW-GS genes and waxy allele compositions of 67 accessions of Sichuan wheat landraces |
 | |
 | |
| aGlu-A1 allele designations are a = subunit 1, c = null, bGlu-B1 allele designations are b = subunits 7+8, c = subunits 7+9, cGlu-D1 allele designations are a = subunits 2+12, da, included the first, the second and the eight group of LMW-GS genes; b, included the first and the second group of LMW-GS genes; c, included the second and the eight group of LMW-GS genes; d, included the first and the eight group of LMW-GS genes; e, included the second group of LMW-GS genes, ef, included the third and the fourth group of LMW-GS genes; g, included the third group of LMW-GS genes; h, included the fourth group of LMW-GS genes, fi, included the five, the sixth, the seventh and the ninth group of LMW-GS genes; j, included the fifth, the sixth and the seventh group of LMW-GS genes; k, included the sixth, the seventh and the ninth group of LMW-GS genes; l, included the fifth, the seventh and the ninth group of LMW-GS genes; m, included the fifth, the sixth and the ninth group of LMW-GS genes, ga, wild type; b, null allele | |
| Table 2: | Group-specific primer sets and chromosome locations of each group of LMW-GS genes |
 | |
Genotyping and Nomenclature
According to the procedure of Ng and Bushuk (1987), HMW-GS were separated by polyacrylamide gel electrophoresis in the presence of sodium dodecyl sulfate (SDS-PAGE). Using two common wheat lines, Chinese Spring (subunits: null, 7+8 and 2+12) and Chuanyu 12 (subunite: 1, 7+8 and 5+10), as references. Five random seeds were initially analyzed for each accession with an additional 15 seeds analyzed for those which were found to be heterogeneous. HMW-GS were designated according the nomenclature of Payne and Lawrence (1983) and the international nomenclature of McIntosh et al. (2003). LMW-GS genes were analyzed according to the PCR method using 7 specific primer sets by Long et al. (2005). These group-specific primer sets and chromosome location of each group of LMW-GS genes showed in Table 2. And waxy alleles were also screened using the PCR method by McLauchlan et al. (2001).
Quality Tests
The harvested grain was tempered to 14.0% moisture and milled on an automatic mill (Brabender, Germany). SDS sedimentation volume (SDS) was determined according to AACC approved method 56-70 by the mixer made by China Agriculture University, a 5 g sample was test per line. Protein content of flours was determined by BUCHI 321 (Kjeldahl Nx5.7), according to ICC standard No. 105. Dough rheologic properties were estimated in a 10 g Brabender Farinograph. The dry gluten, wet gluten and gluten index of the flour and semolina were determined based on ICC 155 using a Gluten Index System made by Perten. The total starch contents were measured by the method NY/T11. The amylose content was measured according to the standard methods NY 147-88 and then the amylopectin content was calculated accordingly. All of the experiments except for farinagraph were done in duplicate.
Statistical Analysis
All data analysis was performed under the software DPS (Data Proceeding System) version 6.10.
RESULTS
Variation of the HMW-GS
The HMW-GS (Glu-1) alleles in sixty-seven Sichuan wheat landraces are shown in k3. All landraces had the same Glu-D1a allele (subunits 2+12). Only two different alleles at Glu-A1 and Glu-B1 were detected, respectively. At the Glu-A1 locus, one landrace, AS1673, contained allele Glu-A1a (subunit 1), while the other landraces had the Glu-A1c (null). At the Glu-B1 locus, all landraces carried the Glu-B1b (subunits 7+8), except AS1563 possessed allele Glu-B1c (subunits 7+9) at the Glu-B1 locus. In these wheat landraces, only three HMW-glutenin compositions were observed, where sixty-five out of sixty-seven landraces (97%) had identical HMW-GS compositions with Chinese Spring (subunits: null, 7+8, 2+12).
Variation of the LMW-GS Genes
This set of Sichuan wheat landraces displayed abundant LMW-GS genes variation (Table 1). Five types of different LMW-GS genes composition at the Glu-A3 locus, three types of different LMW-GS genes composition at the Glu-B3 locus and five types of different LMW-GS genes composition at the Glu-D3 locus were identified. The most common type at Glu-A3 locus was a (74.5%). The other types were found at frequencies of 6.0% (b), 7.5% (c), 4.5% (d) and 7.5% (e), respectively. f (61.2%) was the most common type at the Glu-B3 locus, followed by h (34.3%) and g (4.5%). At the Glu-D3 locus, the frequencies of five types were i (55.2%), j (17.9%), k (4.5%), l (14.9%) and m (7.5%), respectively.
Combined across the three loci, these types form diverse combination patterns with a total of 12 different LMW-GS gene patterns found in all materials. The most common LMW-GS patterns at Glu-A3, -B3 and -D3 were a/f/i (16 materials), a/h/i (11 materials) and a/f/l (6 materials).
Variation of the Waxy Alleles
The PCR analysis of waxy alleles revealed that all of the Sichuan wheat landraces carried the wild type Wx-A1a and Wx-B1a alleles (Table 1).
Variation of the Quality Parameters
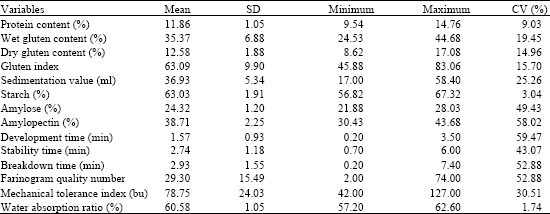
The 67 Sichuan wheat landraces exhibited a large variation for each of the 14 quality parameters recorded. Grain Protein content varied from 9.54 up to 14.76%. Amylopectin content varied from 30.43 to 43.68%. The large variation also was revealed by the Farinograph parameters (Table 3). For example, Farinograph development time had the maximum values of the coefficient of variation (CV = 59.47%) and the stability time comprised between 0.7 up to 6 min. As the Farinograph parameters, the determined values of the dry gluten, wet gluten and gluten index of the flour and semolina also showed large variation (Table 3).
| Table 3: | Value of mean standard deviation, minimum, maximum value and coefficient of variation calculated on the 14 quality parameters |
 | |
| Table 4: | Differences in protein content, sedimentation value, wet gluten content, development time, stability time and breakdown time among types at Glu-A3, Glu-B3 and Glu-D3, respectively |
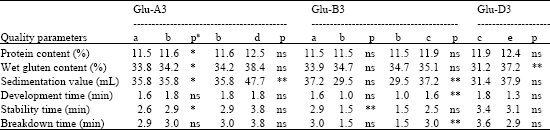 | |
| aSignificance level: *0.01<p<0.05; **0.001<p<0.01, ns: Not significance | |
Association Between LMW-GS Genes and Quality Parameters
Variance analysis was first employed to determine the effect of different LMW-GS gene composition at Glu-A3, Glu-B3 and Glu-D3 loci. Six quality parameters (Protein content, Sedimentation value, Wet gluten content, Development time, Stability time, Breakdown time) have been analyzed (Table 4). All types of LWM-GS genes composition were not significantly different in development time and breakdown time at Glu-A3 locus. But significant difference (p<0.05) between types a and d were found in protein content, sedimentation value, wet gluten content and stability time, whereas the significant difference (p<0.01) between b and d in sedimentation value. At Glu-B3 locus, the three types were nonsignificant difference with each other in protein and wet gluten content. However, the significant difference (p<0.01) between f and g was detected in stability time and between g and h in sedimentation value, development time and breakdown time. At the Glu-D3 locus, significant difference among the five types in all the 6 quality parameters were not detected, except wet gluten content between k and m (p<0.01).
Among the 12 different combination patterns of LMW-GS gene type at Glu-3 locus, variance analysis was also used to determine the difference (Table 5). The pattern (d/f/i) had significantly higher sedimentation value (p<0.01), stability time (p<0.01) and breakdown time (p<0.01) than the pattern (a/g/i). And it also had significantly higher sedimentation value (p<0.01) than the other three patterns (a/f/j, a/f/k and a/f/l). Furthermore, it had significantly higher value (p<0.01) than the pattern (a/f/j) in protein and wet gluten content. The other two patterns (c/f/m and a/h/l) had significantly higher protein content (p<0.01) and wet gluten content (p<0.05), respectively than the pattern (a/f/j). No significant differences were found in development time among 12 different combination patterns.
| Table 5: | Differences in protein content, sedimentation value, wet gluten content, development time, stability time and breakdown time among different LMW-GS gene combined patterns at Glu-3 loci |
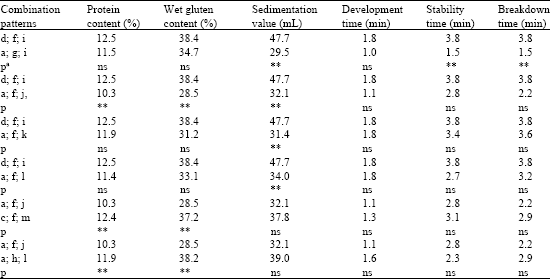 | |
| aSignificance level: *0.01<p<0.05; **0.001<p<0.01, ns: Not significance | |
DISCUSSION
Present results indicated a very small variation of the HMW-GS alleles (with 5 alleles) in Sichuan wheat landraces. Sixty six out of sixty-seven landraces (98.5%) carried Glu-D1a (subunits 2+12) and Glu-A1c allele (null), which were of poor quality for bread making (Payne, 1987). However, at the Glu-B1 locus, all landraces carried the Glu-B1b (subunits 7+8), which were considered by Payne (1987) to have a positive effect on bread making quality, except AS1563. Furthermore, we observed that there was a narrow genetic variation of HMW-GS in Sichuan wheat landraces. This was consistent with the report by Wei et al. (2000). Previously, Zhang et al. (2002) only detected 28 HMW-GS alleles in a core collection of 3,459 Chinese landraces that represented 26.26% of the 13,171 accessions obtained from the National Crop Gene Bank. However, Liu et al. (2007) detected 16 HMW-GS alleles when only analyzing a collection of 111 landraces from Hubei province of China recently. So, the variation frequencies of the HMW-GS alleles may be quite different among populations in china.
According to earlier reports the dominant alleles at the three Glu-1 loci in Chinese wheat landraces were the Glu-A1c (subunit null), Glu-B1b (subunits 7+8) and Glu-D1a (subunits 2+12) (Wei et al., 2000; Zhang et al., 2002; Liu et al., 2007). Present analysis of landraces from Sichuan are consistent with these reports, but the frequencies of the dominant alleles differed in the three populations. In addition, significant differences also occurred in the frequencies of minor alleles between these Chinese landrace collections. For example, whereas low frequencies of the Glu-A1a allele (subunit 1) were observed in the accessions from Sichuan and in the Chinese core collection (Zhang et al., 2002), relatively a high frequency of this allele was found in lines from Hubei (Liu et al., 2007). The frequency of the Glu-B1c (subunits 7+9) alleles at the Glu-B1 locus was also lower in our study than Hubei wheat landraces (Liu et al., 2007). It can also be noted that all sixty-seven landraces from Sichuan had the Glu-D1a (subunits 2+12) alleles compared with 108/111 (97.64%) in the Hubei accessions (Liu et al., 2007).
Screening for LMW-GS was difficult, not only because of the large number of bands, some of which overlapped, but also because of different patterns resolved under different gel conditions and reported in different publications (Branlard et al., 2003; Gupta and Shepherd, 1990). So, we directly identified LMW-GS gene groups by PCR method (Long et al., 2005), which was more accurate and efficient. The method would be very useful for establishing allele identification standards.
Similar to the efforts to establish associations between HMW-GS alleles and dough quality, studies have also been conducted to link LMW-GS variants to quality traits. Both positive and negative effects of specific LMW-GS on wheat flour quality have been reported by Branlard et al. (2001) and Eagles et al. (2002). In present study, we directly identified LMW-GS gene groups by PCR method and nine groups (Long et al. 2005) have been determined at the Sixty-seven Sichuan wheat landraces Glu-3 loci. But it was not that all of the groups had been found at every landrace Glu-3 loci. Five, three and five types of different LMW-GS gene group composition have been found at Glu-A3, Glu-B3 and Glu-D3 locus respectively. Significant difference (p<0.05) had been found in protein content, sedimentation value, wet gluten content and stability time between a and d at Glu-A3. From the Table 1, type a contained the first, the second and the eighth groups of LMW-GS genes, type d contained the first, the eighth groups of LMW-GS genes. And type a always associated with low protein content, sedimentation value, wet gluten content and stability time when compared with type d (date not shown). So, the second group of LMW-GS genes might have a negative effect on bread making quality. Long et al. (2005) clarified that the LMW-GS genes of group 2 contained less than 90 amino acid residues and included only 13 repeat in its repetitive domain, which was the shortest among the nine LMW-GS groups. The number of repeats present in the repetitive domain of LMW-GS is mainly responsible for the general hydrophilic character (D’Ovidio et al., 1997; D’Ovidio and Masci, 2004).
At Glu-B3 locus, stability time is significant difference (p<0.01) between f (include the third and fourth groups) and g (only include the third group). It is indicated the fourth group of LMW-GS genes might play an important role in determining stability tine. Four LMW-GS gene groups were assigned to the short arm of chromosome 1D at Glu-D3 locus, while three and two groups were located on the short arms of chromosome 1A and 1B at Glu-A3 and Glu-B3 loci, respectively (Long et al., 2005). It was more complicated than Glu-A3 and Glu-B3 loci for analyzing. But significant difference (p<0.01) was also found in wet gluten content between types k and m. From the Table 1, type k contained the sixth, the seventh and the ninth groups of LMW-GS genes, type m contained the fifth, the sixth and the ninth groups of LMW-GS genes. The difference between the fifth group and the seventh group might cause significant difference (p<0.01) in wet gluten content between types k and m. In marker-assisted breeding, we can choose the fifth group or the seventh group for high or low wet gluten content wheat according to our intention.
Sixty seven Sichuan wheat landraces didn’t have null alleles at Wx-A1, Wx-B1 and Wx-D1 locus. Du et al. (2007) determined 1739 China wheat landraces and found only 31 waxy mutation types (mutation frequency was 1.8%). So, the natural mutation frequency of waxy allele was very low.
CONCLUSION
In present study, only 5 alleles were detected at the three HMW-GS Glu-1 loci in sixty-seven Sichuan wheat landraces of China. The lower variation of HMW-GS alleles in these materials was found compared with Hubei wheat landraces. A total of 13 types of different LMW-GS genes composition at Glu-3 loci were observed. Materials containing the type d had higher protein content, sedimentation value, wet gluten content and stability time than materials with type a. And significant differences were found in sedimentation value, development time and breakdown time between type g and h. Among the 12 different combination patterns of LMW-GS gene type at Glu-3 loci, the pattern (d/f/i) had significantly higher sedimentation value, stability time and breakdown time than the pattern (a/g/i). All of the landraces carried the wild type Wx-A1a and Wx-B1a alleles. This study documents the variability at important quality loci and illustrated the relationships between the types of LMW-GS gene composition and quality characteristics in Sichuan wheat germplasm. These results will be useful to plan crossing and selection strategies to improve end-use quality and to breed high-quality variety.
ACKNOWLEDGMENTS
This study was supported by the National High Technology Research and Development Program of China (863 program 2006AA10Z179 and 2006AA10Z1F8), the Key Technologies R and D Program (2006BAD01A02-23) and the FANEDD project (200357 and 200458) from Ministry of Education, China. Y.-M.Wei was supported by the Program for New Century Excellent Talents in Universities of China (NCET-05-814). Y.-L. Zheng was supported by the Program for Changjiang Scholars and Innovative Research Teams in Universities of China (IRT0453).
REFERENCES
- Branlard, G., M. Dardevet, R. Saccomano, F. Lagoutte and J. Gourdon, 2001. Genetic diversity of wheat storage proteins and bread wheat quality. Euphytica, 119: 59-67.
CrossRef - Branlard, G., M. Dardevet, N. Amiour and G. Igrejas, 2003. Allelic diversity of HMW and LMW glutenin subunits and omega-gliadins in french bread wheat (Triticum aestivum L.). Genet. Resour. Crop. Evol., 50: 669-679.
CrossRef - Cassidy, B.G., J. Dvorak and O.D. Anderson, 1998. The wheat low-molecular-weight glutenin genes: Characterization of six genes and progress in understanding gene family structure. Theor. Applied Genet., 96: 743-750.
CrossRefDirect Link - D'Ovidio, R., M. Simeone, S. Masci and E. Porceddu, 1997. Molecular characterization of a LMW-GS gene located on chromosome 1B and the development of primers specific for the Glu-B3 complex locus in durum wheat. Theor. Applied Genet., 95: 1119-1126.
CrossRef - Eagles, H.A., G.J. Hollamby, N.N. Gororo and R.F. Eastwood, 2002. Estimation and utilization of glutenin gene effects from the analysis of the unbalanced data from the wheat breeding programs. Aust. J. Agric. Res., 53: 367-377.
CrossRefDirect Link - Gianibelli, M.C., O.R. Larroque, F. MacRitchie and C.W. Wrigley, 2001. Biochemical, genetic and molecular characterization of wheat glutenin and its component subunits. Cereal Chem., 78: 635-646.
CrossRef - Graybosch, R.A., 1992. High molecular weight glutenin subunit composition of cultivars, germplasm and parents of US red winter wheat. Crop Sci., 32: 1151-1155.
Direct Link - Gupta, R.B. and K.W. Shepherd, 1990. Two-step one-dimensional SDS-PAGE analysis of LMW subunits of glutelin. Theor. Applied Genet., 80: 65-74.
CrossRef - Gupta, R.B., J.G. Paul, G.B. Cornish, G.A. Palmer and F. Bekes et al., 1994. Allelic variation at glutenin subunit and gliadin loci, Glu-1, Glu-3 and Gli-1, of common wheat, I: Its additive and interaction effects on dough properties. J. Cereal Sci., 19: 9-17.
CrossRef - Harberd, N.P., D. Bartels and R.D. Thompson, 1985. Analysis of the gliadin multigene loci in bread wheat using nullisomic-tetrasomic lines. Mol. Gen. Genet., 198: 234-242.
CrossRefDirect Link - Ikeda, T.M., T. Nagamine, H. Fukuoka and H. Yano, 2002. Identification of new low-molecular-weight glutenin subunit genes in wheat. Theor. Applied Genet., 104: 680-687.
CrossRefPubMedDirect Link - Lawrence, G.J. and K.W. Shepherd, 1981. Chromosomal location of genes controlling seed proteins in species related to wheat. Theor. Applied Genet., 59: 25-31.
CrossRef - Liu, Y., Z.Y. Xiong, Y.G. He, R.S. Peter and G.Y. He, 2007. Genetic diversity of HMW glutenin subunit in Chinese common wheat (Triticum aestivum L.) landraces from Hubei province. Genet. Resour. Crop. Evol., 54: 865-874.
CrossRef - Long, H., Y.M. Wei, Z.H. Yan, B. Baum, E. Nevo and Y.L. Zheng, 2005. Classification of wheat low-molecular-weight glutenin subunit genes and its chromosome assignment by developing LMW-GS group-specific primers. Theor. Applied Genet., 111: 1251-1259.
CrossRefPubMedDirect Link - Martin, J.M., L.E. Talbert, D.K. Habernicht, S.P. Lanning and J.D. Sherman et al., 2004. Reduced amylose effects on bread and white salted noodle quality. Cereal Chem., 81: 188-193.
Direct Link - McLauchlan, A., F.C. Ogbonnaya, B. Hollingsworth, M. Carter and K.R. Gale et al., 2001. Development of robust PCR-based DNA markers for each homoeoallele of granule-bound starch synthase and their application in wheat breeding programs. Aust. J. Agric. Res., 52: 1409-1416.
Direct Link - Murai, J., T. Taira and D. Ohta, 1999. Isolation and characterization of the three waxy genes encoding the granule-bound starch synthase in hexaploid wheat. Genet, 234: 71-79.
CrossRef - Nakamura, T., M. Yamamori, H. Hirano and S. Hidaka, 1993. Identification of three wX proteins in wheat (Triticum aestivum L.). Biochem. Genet, 31: 75-86.
CrossRef - Payne, P.I., K.G. Corfield and J.A. Blackman, 1979. Identification of a highmolecular-weight subunit of glutenin whose presence correlates with bread-making quality in wheats of related pedigree. Theor. Applied Genet, 55: 153-159.
CrossRef - Payne, P.I., C.N. Law and E.E. Mudd, 1980. Control by homologous group 1 chromosomes of the high-molecular-weight subunits of glutenin, a major protein of wheat endosperm. Theor. Applied Genet., 58: 113-120.
CrossRef - Payne, P.I., K.G. Corfield, L.M. Holt and J.A. Blackman, 1981. Correlations between the inheritance of certain high-molecular-weight subunits of glutenin and breadmaking quality in progenies of six crosses of bread wheat. J. Sci. Food Agric., 32: 51-60.
CrossRef - Redaelli, R., P.K.W. Ng and N.E. Pogna, 1997. Allelic variation at the storage protein loci of 55 uS-grown white wheats. Plant Breed., 116: 429-436.
CrossRef - Singh, N.K. and K.W. Shepherd, 1985. The structure and genetic control of a new class of disulphide-linked proteins in wheat endosperm. Theor. Applied Genet., 71: 79-92.
CrossRefDirect Link - Sabelli, P. and P.R. Shewry, 1991. Characterization and organization of gene families at the Gli-1 loci of bread and durum wheat. Theor. Applied Genet., 83: 209-216.
CrossRefDirect Link - Van Campenhout, S., S.J. Vander, L. Sagi and G. Volckaert, 1995. Locus-specific primers for LMW glutenin genes on each of the group1 chromosomes of hexaploid wheat. Theor. Applied Genet., 91: 313-319.
CrossRef - Zhao, X.C., P.J. Sharp, G. Crosbie, I. Barclay and R. Wilson et al., 1998. A single genetic locus associated with starch granule properties in a cross between wheat cultivars of disparate noodle quality. J. Cereal Sci., 27: 7-13.
CrossRef - Zhang, W., M.C. Gianibelli, L.R. Rampling and K.R. Gale, 2004. Characterisation and marker development for low molecular weight glutenin genes from Glu-A3 alleles of bread wheat (Triticum aestivum. L). Theor. Applied Genet., 108: 1409-1419.
CrossRef - Ng, P.K.W. and W. Bushuk, 1987. Glutenin of Marquis wheat as a reference for estimating molecular weights of glutenin subunits by sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Cereal Chem., 64: 324-327.
Direct Link - D'Ovidio, R. and S. Masci, 2004. The low-molecular-weight glutenin subunits of wheat gluten. J. Cereal Sci., 39: 321-339.
CrossRefDirect Link