
J. Aghofack-Nguemezi
Department of Plant Biology, Faculty of Sciences, University of Dschang, P.O. Box 67, Dschang, Cameroon
G. Kanmegne
Department of Plant Biology, Faculty of Sciences, University of Dschang, P.O. Box 67, Dschang, Cameroon
J. Manka`abiengwa
Department of Plant Biology, Faculty of Sciences, University of Dschang, P.O. Box 67, Dschang, Cameroon
Journal of Plant Sciences
Year: 2008 | Volume: 3 | Issue: 4 | Page No.: 248-254







ABSTRACT
Banana fruits were treated with 2, 4-dichlorophenoxyacetic acid or naphthalenacetic acid solutions at concentrations ranging from 10-6 to 10-3 M. The treated fruits were either further anointed or not with coconut oil or palm kernel oil. Untreated fruits were used as controls. Treatments with solutions of both types of auxins at concentrations >=10-4 M strongly accelerated the ripening of bananas whereas treatments with solutions of these hormones at 10-6 M concentration induced an increase of the shelf-life of bananas. Although singly treatments with coconut oil or palm kernel oil also had retarding effects on the ripening, simultaneous treatments with auxins used at concentrations >=10-4 M and any of the two oils rather accelerated this process in banana fruits. Treatments with an auxin alone or in combination with an oil stimulated the decrease in water content in the peel from stage 1 to stage 3 of ripeness. Those treatments that prolonged the shelf-life of bananas subsequently induced a delay in the decrease in water content in the peel from stage 3 of ripening upwards. These results indicated that the role of ethylene in the regulation of ripening process in banana fruits may be modulated by the endogenous concentration of auxin.
PDF Abstract XML References Citation
How to cite this article
J. Aghofack-Nguemezi, G. Kanmegne and J. Manka`abiengwa, 2008. Effects of Auxins and Edible Oils on the Ripening and Water Content of Banana Fruits. Journal of Plant Sciences, 3: 248-254.
DOI: 10.3923/jps.2008.248.254
URL: https://scialert.net/abstract/?doi=jps.2008.248.254
DOI: 10.3923/jps.2008.248.254
URL: https://scialert.net/abstract/?doi=jps.2008.248.254
INTRODUCTION
Ethylene is the major phytohormone that directly regulates the process of ripening in fruits. In climacteric fruits such as bananas there is a sharp peak of ethylene production at the early stage of ripeness and exogenously applied ethylene induces ripening and endogenous ethylene production (Lelièvre et al., 1997; White, 2002). Treatment of mature green bananas with ethylene antagonist 1-methylcyclopropene showed that ethylene has a continuing role in integrating some biochemical process during the ripening of fruits (Golding et al., 1999). There are four levels of manipulation one can use to regulate ethylene response (Nagar and Sood, 2004): (a) control the concentration of ethylene in the tissue by addition or removal of ethylene, (b) regulate the concentration in the tissue by stimulating or inhibiting ethylene biosynthesis, (c) modify the binding characteristics of ethylene receptors and (d) manipulate ethylene-dependent gene expression. The regulation of ethylene concentration in the tissue could be achieved through synergistic or antagonistic effects of other hormones. Auxin and ethylene, for example, interact mutually in the control of many growth and development processes (Gaspar et al., 2000; Naqvi, 2002). A close relationship has been demonstrated between the rate of ethylene production and the level of the free auxin indoleacetic acid (IAA), which was regulated by the cytokinin kinetin in hypocotyl segments of mungbean seedlings (Lau and Yang, 1995). 1-Aminocyclopropane-1-carboxylic acid synthase, a key regulatory enzyme in the biosynthesis of ethylene, was encoded by divergent multigene family in Arabidopsis thaliana, comprising at least five genes, ACS1-5 (Liang et al., 1992). In etiolated pea and soybean seedlings ACS4 was specifically induced by IAA. This induction took place within 25 min and was incentive to protein synthesis inhibition, suggesting that the ACS4 gene expression is a primary response to IAA (Abel et al., 1995).
Alterations in the chemical nature of the environment of hormone receptor could result from changes in the membrane lipid composition. It is well established that lipids are constituents of plant and animal cell membranes and their chemical nature may determine the physical properties of membranes and modulate many physiological processes (Sommerville et al., 2000; Aghofack-Nguemezi, 2001; Aghofack-Nguemezi and Tatchago, 2006; Torres-Franklin et al., 2007). In flower senescence and banana ripening, short-chain fatty acids may increase the sensitivity of tissue to ethylene as a result of increase ethylene binding (Whitehead and Bossè, 1991; Whitehead and Valsijevic, 1993). Treatments with coconut oil (CNO) or Palm Kernel Oil (PKO) induced a delay in several changes that usually accompanied ripening, thereby inducing an increase of shelf-life of banana fruits (Aghofack-Nguemezi et al., 2006). When used simultaneously with CNO or PKO, treatments with CaCl2 or MgSO4 solutions which induced an increase in the level of Ca2+ in the peel had synergistic delaying effects on some processes during banana fruit ripening (Aghofack-Nguemezi and Dassie, 2007).
To the best of our knowledge, no study exists on the influence on one hand of treatments with auxins at different concentrations and on the other hand of simultaneous treatments with auxins and oils on the ripening of banana fruits. The present research was undertaken to investigate the effects of treatments with 2, 4-dichlorophenoxyacetic acid (2, 4-D) or naphthalenacetic acid (NAA) at 10-6-10-3 M concentrations and coconut oil or palm kernel oil on the color of the peel and the water content in both the peel and pulp, changes in these parameters being useful ripening-related index for banana fruits.
MATERIALS AND METHODS
Plant Materials and Treatments
The study was conducted in 2005 and 2006. Musa accuminata Colla fruits were donated by the High Penja Plantation Company (PHP) in Njombe, Cameroon. These fruits were from the second hand of bunches. The oils were obtained from the endosperm of palm kernel and coconut after grinding and squeezing. Green, unripe bananas were treated by dipping them for 24 h separately in 2, 4-dichlorophenoxyacetic acid and naphthalenacetic acid solutions at concentrations raging from 10-6 to 10-4 M. The treated fruits were further either anointed or not with 100 μL of coconut oil or palm kernel oil. Untreated fruits were used as controls. Control and treated bananas were packed in transparent polythene bags.
Visual Assessment of Ripening Stages and Determination of Water Content
The seven ripening stages were visually assessed according to Dadzie and Ochard (1997). Banana at stage 1 (green, unripe), 3 (greener than yellow), 5 (yellow with green tip) and 7 (yellow, slightly flecked) of ripening of control fruits and at extra-stage were used for the determination of water content. The stage 1 was considered as the zero, starting point for all experiments. The extra-stage was when all the controls bananas had already reached stage 7.
For the determination of the water content, fresh peel and pulp were weighed and dried in an oven at 105°C for 24 h. They were then weighed and the water content was calculated using fresh and dry matter weight according to Chapman (1976).
Statistical Analysis
Group comparison were made using One-way Analysis of Variance (ANOVA) to see if variations among the means were significantly greater than expected by chance. The Student-Newman-Keuls Test was used to compared means differences, whereby a p-value of <0.05 was considered as statistically significant.
RESULTS
The visual assessment of ripening stages showed that singly treatments with coconut oil (CNO) or Palm Kernel Oil (PKO) induced an increase of the shelf-life of banana fruits. Treatments with 2, 4-dichlorophenoxyacetic acid (2, 4-D) or naphthalenacetic acid (NAA) at concentrations ≥10-4 M either alone or simultaneously with CNO or PKO accelerated the ripening of bananas. However, treatments with solutions of NAA at 10-4 M in combination with CNO or PKO were less effective in hastening the ripening of bananas. Thus, fruits treated with 10-3 M 2, 4-D, 10-3 M 2, 4-D + CNO or PKO, 10-4 M 2, 4-D, 10-4 M 2, 4-D + CNO or PKO, 10-3 M NAA and 10-3 M NAA + CNO or PKO turned fully to yellow while control bananas were still at stage 3; bananas treated with solutions of 10-4 M NAA either singly or simultaneously with CNO or PKO reached stage 7 while the untreated fruits were still at stage 5. Contrarily to auxin solutions at concentrations ≥10-4 M, treatments with 2, 4-D or NAA solutions at concentrations ≤10-5 M induced a delay in the ripening of bananas. Simultaneous treatment with 2, 4-D or NAA solutions at concentrations ≤10-5 M and CNO or PKO were more effective in increasing the shelf-life of banana fruits.
Table 1 shows changes in the water content in the peel during the ripening of banana fruits. There were significant decreases in the water contents in the peel of most of the treated fruits from stage 1 to stage 3 of ripeness whereas this parameter did not change significantly in the peel of control fruits during this period.
| Table 1: | Effects of treatments with 2, 4-dichlorophenoxyacetic acid (2, 4-D) or naphthalenacetic acid (NAA) and/or palm kernel oil (PKO) or coconut oil (CNO) on the water content in the peel of banana fruits. Values are means of at least three replications followed by standards deviations between parentheses. The water content in the peel of green unripe banana fruits (stage 1) is considered as the starting point for all treatments |
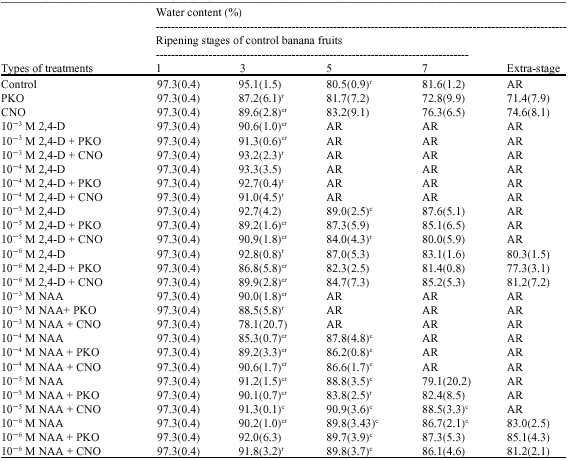 | |
| c: Significantly different from the control in the same column at p<0.05, r: Significantly different from the preceding value in the same row at p<0.05, AR: Fruits had already reached the yellow-slightly-fecked stage | |
| Table 2: | Effects of treatments with 2, 4-dichlorophenoxyacetic acid (2, 4-D) or naphthalenacetic acid (NAA) and/or palm kernel oil (PKO) or coconut oil (CNO) on the water content in the pulp of banana fruits. Values are means of at least three replications followed by standards deviations between parentheses. The water content in the pulp of green unripe banana fruits (stage 1) is considered as the starting point for all treatments |
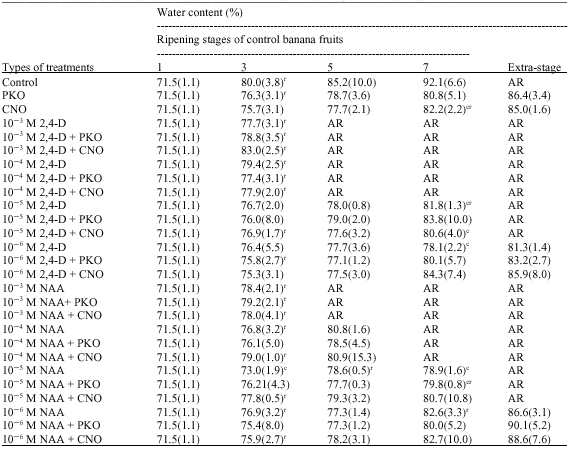 | |
| c: Significantly different from the control in the same column at p<0.05, r: Significantly different from the preceding value in the same row at p<0.05, AR: Fruits had already reached the yellow-slightly-fecked stage | |
When compared pairwise, the water contents in the peel of banana fruits at stage 3 and 5, 5 and 7 and 7 and extra-stage (stage where untreated fruits did no longer exist) were not significantly different from each other in most of the cases. Nevertheless, the water content in the peel of control as well as treated fruits indicated a general decreasing trend from stage 1 to stage 7/extra stage of ripeness. At stage 3 of ripening of control fruits, the water contents in the peel of bananas treated with CNO, 10-3 M 2, 4-D, 10-3 M 2, 4-D + PKO, 10-5 M 2, 4-D + PKO or CNO, 10-6 M 2, 4-D + CNO or PKO, 10-3 M NAA, 10-4 M NAA, 10-4 M NAA + CNO or PKO, 10-5 NAA and 10-5 M NAA + CNO or PKO were significantly lower than that observed in the peel of control fruits. At stage 5 of ripening of control fruits, the water contents in the peel of bananas treated with 10-5 M 2, 4-D, 10-4 M NAA singly or simultaneously with CNO or PKO and 10-6 M NAA in combination with CNO or PKO were significantly higher than what was found in the peel of control fruits. Apart from fruits treated with 10-5 M NAA + CNO or with 10-6 M NAA no significant difference in the water content in the peel could be observed between treated and control bananas at stage 7 of ripening of control fruits. At this stage, the water contents in the peel of bananas treated with 10-5 M NAA + CNO or with 10-6 M NAA were significantly higher than the water content in the peel of untreated fruits.
The water contents in the pulp of both treated and control banana fruits generally increased from stage 1 to stage 7 and extra-stage of ripening (Table 2). In most of the cases significant increases in the water content in the pulp were observed only between stage 1 and stage 3 of ripeness. At stages 3, 5 and 7 of ripening of control, the water contents in the pulp of treated fruits were lower than that observed in untreated bananas. It was only in some cases that these effects of treatments on the water content in the pulp were significant. Thus, at stage 3 of ripening of control bananas, the water content in the pulp of fruits treated with NAA solution at 10-5 M was significantly lower than the water content in the pulp of control fruits; at stage 7 of ripeness of untreated fruits the water contents in the pulp of bananas treated with CNO, 10-5 M 2, 4-D, 10-5 M 2, 4-D + CNO, 10-6 M 2, 4-D, 10-5 M NAA and 10-5 M NAA + PKO were significantly lower than that observed in the pulp of control fruits.
DISCUSSION
Treatments of banana fruits with NAA or 2, 4-D had two concentration-dependent opposed effects on ripening process in banana fruits. The mechanisms of these contrary effects of exogenously applied auxin on banana ripening are not exactly known. In vivo, auxin and ethylene may interact mutually in the regulation of growth and development processes like senescence (Naqvi, 2002). Fowler and Morgan (1972) reported that eight hours after the application of 2, 4-dichlorophenoxyacetic acid and indoleacetic acid (IAA), the initial levels of ethylene were increased in cotton plants and that these magnitudes were maintained for a 72 h-period of observation. Modifications of peroxidase and IAA oxidase activities in auxin treated cotton plants definitely occurred after the elevation of internal ethylene levels. Obviously, at least in vegetative plant parts, high endogenous level of auxin stimulate the biosynthesis of ethylene. On the other hand, when the internal ethylene concentration reaches a certain threshold, it induces an elevation of the activities of peroxidase and related enzymes like auxin oxidase (IAA oxidase) that leads to the feedback destruction of auxin (Gaspar et al., 2000). However, it remains unclear how low concentrations of auxin could inhibit the ethylene biosynthesis and induce a delay in the ripening of banana fruits.
Application of coconut oil or palm kernel oil had synergistic effects with treatments with 2, 4-D or NAA at concentration ≤10-5 M whereas these oils did not influence the effects of the two auxins at concentrations ≥10-4 M on banana ripening. Since the fruits were first treated by dipping for 24 h in auxin solutions before being anointed with oils, 2, 4-D or NAA solutions at concentrations ≥10-4 M may have initiated so strong ripening stimulating effects that could no longer be overcome by oil treatments. Changes in membrane lipids of the banana peel as a result of treatments with oils may have led to alterations in the ethylene signaling pathways. In this context, a relationship has been established between ethylene signaling, membrane sterol and meristem function in Arabidopsis thaliana (L.) heynth (Souter et al., 2004).
The water content in the peel or pulp generally decreased or increased respectively with increasing stages of ripening of control as well as treated fruits. Those treatments that accelerated the ripening process stimulated changes in the water content whereas treatment that prolonged the shelf-life of banana fruits induced delays in alterations in the water content. During ripening of fleshy fruits the peel losses water either through transpiration or diffusion into the pulp by osmosis (John and Marchal, 1995). The water content usually increases in the pulp due to transformation of starch into reducing sugars and consequently to an increase of osmotic potential that induces the movement of water from the peel to the pulp (Ngalani et al., 1999; Cordenunsi and Lajolo, 1995). The water-producing reactions like respiration also contribute to the increase in the water content in the pulp (Wills et al., 1998). The mechanisms whereby treatments with auxin solutions and/or oils affected changes in the water content in the peel and pulp of banana fruits are not exactly known. It is well established that oil represents an excellent barrier against water (Badwin, 2001). Alterations in water content in the peel and pulp as a result of treatments of fruits with auxin solutions and/or oils may be an indirect effect of changes in other metabolic activities that usually accompanied the ripening process in banana fruit.
REFERENCES
- Abel, S., M.D. Nguyen, W. Chow and A. Theologis, 1995. ACS4, a primary indoleacetic acid-responsive gene encoding 1-amino-cyclopropane-1-carboxylate synthase in Arabidopsis thaliana. Structural characterization, expression in Escherichia coli and expression characteristics in response to auxin. J. Biol. Chem., 270: 19093-19099.
PubMed - Aghofack-Nguemezi, J. and V. Tatchago, 2006. Changes in glycerolipid and sterol contents in leaves of Borago officinalis L. plants as related to growth and development. Pak. J. Biol. Sci., 9: 686-691.
CrossRefDirect Link - Aghofack-Nguemezi, J., L.B. Tonfack, S. Kaptue, S. Schoch and U. Graefe, 2006. Effects of coconut oil and palm kernel oil treatments on ripening process in banana fruits. Cameroon J. Biol. Sci., 2: 16-21.
CrossRefDirect Link - Aghofack-Nguemezi, J. and B. Dassie, 2007. Effects of salts and edible oils on calcium and water contents in ripening banana fruits. J. Plant Sci., 2: 302-309.
CrossRefDirect Link - Cordenunsi, B.R. and F.M. Lajolo, 1995. Starch breakdown during ripening. Sucrose synthase and sucrose phosphate synthase. J. Agric. Food Chem., 43: 347-351.
CrossRef - Gaspar, T., C. Kevers, J.F. Hausman, O. Faivre-Rampant, N. Boyer, J. Dommes, C. Penel and H. Greppin, 2000. Integrating Phytohormone Metabolism and Action with Primary Biochemical Pathways. I. Interrelationships Between Auxins, Cytokinins, Ethylene and Polyamines in Growth and Development Processes. In: Intergrated Plant Systems, Greppin, H., C. Penel, W.J. Broughton and R. Strasser (Eds.). University of Geneva Press, Geneva, pp: 163-191.
- Lelievre, J.M., A. Latache, B. Jones, M. Bouzayem and J.C. Pech, 1997. Ethylene and fruit ripening. Physiol. Plant, 101: 727-739.
CrossRefDirect Link - Liang, X., S. Abel, J.A. Keller, N.F. Shen and A. Theologis, 1992. The 1-aminocyclopropane-1-carboxylate synthase gene family of Arabidopsis thaliana. Proc. Natl. Acad. Sci., 89: 11046-11050.
Direct Link - Naqvi, S.S.M., 2001. Plant Growth Hormones: Growth Promoters and Inhibitors. In: Handbook of Plant Crop Physiology, Pessarakli, M. (Ed.), CRC Press, New York, ISBN: 9780429208096, pp: 523-548.
CrossRefDirect Link - Souter, M.A., M.L. Pullen, J.F. Topping, X. Zhang and K. Lindsey, 2004. Rescue of defective auxin-mediated gene expression and root meristem function by inhibition of ethylene signaling in sterol biosynthesis mutants of Arabidopsis. Planta, 219: 773-783.
Direct Link - Torres-Franklin, M.L., A. Gigon, D.F. de Melo, Y. Zuily-Fodil and A.T. Pham-Thi, 2007. Drought stress and rehydratation affect the balance between MGDG and DGDG synthesis in cowpea leaves. Physiol. Plant, 131: 201-210.
Direct Link - White, P.J., 2002. Recent advances in fruit development and ripening: An overview. J. Exp. Bot., 53: 1995-2000.
Direct Link - Whitehead, C.S. and C.A. Bosse, 1991. The effect of ethylene and short-chain saturated fatty acids on ethylene sensitivity and binding in ripening bananas. J. Plant Physiol., 137: 358-362.
CrossRefDirect Link