
A. Soltani
Department of Agronomy and Plant Breeding,
Gorgan University of Agricultural Sciences, P.O. Box 386, Gorgan, Iran
M. Gholipoor
Department of Agronomy and Plant Breeding, Shahrood University ofTechno1ogy, Shahrood, Iran
K. Ghassemi-Golezani
Departrnent of Agronomy and Plant Breeding, Tabriz University, Tabriz, Iran
Journal of Plant Sciences
Year: 2007 | Volume: 2 | Issue: 1 | Page No.: 89-95







ABSTRACT
In this study, a simple theoretical framework was extended to account for the effects of temperature and atmospheric CO2 concentration on RUE. A general test of the model showed that simulated and observed daily biomass productions under average daily temperatures ranged from 11 to 28°C are similar. The calculated RUEs for 13-23°C were similar to measured ones and percentage increase in RUE for CO2 concentration of 640 ppm relative to 330 ppm was in agreement with the measured percentage increase in biomass production. By using the framework, RUE response functions to average daily temperature and CO2 concentration were calculated for chickpea. RUE of chickpea at temperatures lower than 3°C and higher than 36°C is zero. RUE rapidly increased (9.5%°C-1) with increasing temperature from 3 to 14°C. With further increase in temperature to 22°C, RUE slowly (1.4%°C-1) decreased and temperature increase between 22 to 36°C resulted in sharp decrease (7.4%°C-1) in RUE. Response of RUE to CO2 concentration was curvilinear. At low concentrations of CO2 (60 to 400 ppm), RUE was especially sensitive to increases in CO2 concentration, but increases in CO2 greater than 700 ppm were predicted to result in only small increases in RUE. The functions obtained can be used in simulation studies of chickpea crop response to projected climate change.
PDF Abstract XML References
How to cite this article
A. Soltani, M. Gholipoor and K. Ghassemi-Golezani, 2007. Analysis of Temperature and Atmospheric CO2 Effects on Radiation Use Efficiency in Chickpea (Cicer arietinum L.). Journal of Plant Sciences, 2: 89-95.
DOI: 10.3923/jps.2007.89.95
URL: https://scialert.net/abstract/?doi=jps.2007.89.95
DOI: 10.3923/jps.2007.89.95
URL: https://scialert.net/abstract/?doi=jps.2007.89.95
INTRODUCTION
Chickpea (Cicer arietinum L.) is one of the major pulse crops in arid and semiarid environments of West Asia and North Africa (WANA) region and has considerable importance as food, feed and fodder. Due to the increasing need for legumes, chickpea is no longer considered a subsistence crop (Saxena et al., 1996). Projected global climate change may have major influences on cropping aspects of this crop in the region. In order to ensure a balanced growth and development in agriculture in the region, a comprehensive assessment of the vulnerability of chickpea cropping due to projected climate change is required.
Radiation use efficiency (RUE, g MJ-1) is the amount of biomass accumulated for each unit of total solar radiation intercepted by leaf canopy (Choudhury, 2000). Recently, a simple mechanistically based model was developed for chickpea in which the daily biomass production is calculated using RUE (Soltani et al., 1999). In this model, the effect of supra-optimal temperatures and atmospheric CO2 concentrations were not included and the effect of low temperature was only considered. The effect of temperature and CO2 on RUE is especially important in climate change studies using RUE based models. However, there is no report on the response functions of RUE to temperature and CO2 concentration in chickpea.
Sinclair and Horie (1989), Sinclair (1991), Sinclair et al. (1992) and Hammer and Wright (1994) developed and extended a framework to analyze the effects of radiation, leaf nitrogen, fraction diffuse radiation and leaf light-saturated CO2 assimilation rate on RUE. This framework provides a simple and effective method to analyses the variation of RUE between species and under different conditions. The framework, however, does not account for the effects of temperature and CO2 concentration.
Our objectives of this study were: (1) to extend and test the theoretical framework to model the effects of average daily temperature and atmospheric CO2 concentration on RUE and (2) to use the extended framework to produce the response functions of chickpea RUE to average daily temperature and atmospheric CO2.
MATERIALS AND METHODS
The Theoretical Framework Used
Crop radiation use efficiency on any day is calculated as the ratio of crop biomass produced that day to total solar radiation intercepted by the canopy that day. Crop biomass increment is determined from canopy gross CO2 assimilation rate, daily rate of maintenance respiration costs and the assimilate requirement for dry matter production (Goudriaan and van Laar, 1994; Choudhury, 2000). Growth respiration and the efficiency of conversion of glucose to plant tissue was computed using the procedure of Penning de Vries et al. (1989). The maintenance coefficients for chickpea were considered as 0.03 for leaves, 0.015 for stems, 0.015 for roots and 0.01 for storage organs based on Penning de Vries et al. (1989) and Goudriaan and van Laar (1994). When organs dry matter are not available, maintenance respiration is calculated as a fraction of the daily total hexose production. The fraction has a value of 0.15-0.2 (Sinclair and Horie, 1989).
Canopy gross assimilation is calculated by integrating the instantaneous assimilation rates of sun leaves and shade leaves over the day light periods. The instantaneous assimilation rates are calculated from the product of leaf area index of sunlit or shade leaves and the assimilation rate per unit leaf area (P). P is given by an asymptotic exponential equation of radiation flux density incident on either sun or shade leaves (KJ m-2 sec-1) (Boote and Loomis, 1991):
| (1) |
where Pmax (mg CO2 m-2 sec-1) is the assimilation rate per unit leaf area asymptote to leaf potential for CO2 assimilation rate per unit leaf area corrected for temperature and CO2 concentration and QEc (mg CO2 kJ-1) is the light use efficiency at low light corrected for temperature and atmospheric CO2.
The leaf area index of sun or shade leaves is calculated by considering the sun angle elevation, the proportion of intercepted radiation and shadow projection function (Boote and Loomis, 1991). The radiation flux density, incident on either sun or shade leaves is calculated from the incident direct and diffuse radiation, the proportion of incident radiation intercepted by the leaf canopy and the leaf area index of sun or shade leaves in a manner similar to Sinclair et al. (1992). The direct and diffuse components of the total incident radiation at any time are calculated in a manner similar to that of Goudriaan and van Laar (1994).
Pmaxc and QEc, required in Eq. 1 are calculated by:
| (2) |
| (3) |
where Pmax is leaf light-saturated CO2 assimilation rate (mg CO2 m-2 sec-1), Ptmp is a correction factor (between 0 to 1) of Pmax for temperature,![]() is a correction factor of Pmax for atmospheric CO2 concentration, QE is the light use efficiency at low light which was set at 5 mg CO2 KJ-1 (Sinclair and Horie, 1989) and
is a correction factor of Pmax for atmospheric CO2 concentration, QE is the light use efficiency at low light which was set at 5 mg CO2 KJ-1 (Sinclair and Horie, 1989) and ![]() is a correction factor of QE for CO2 concentrations.
is a correction factor of QE for CO2 concentrations.
The correction of the Pmax and QE for the effect of atmospheric CO2 is modeled using Farquhar and von Cammerer (1982) procedure for limiting RuBP as used by Boote and Pickering (1994) and Yu et al. (2003):
| (4) |
| (5) |
where CiPmax is internal CO2 concentration for the computation of ![]() with a Ci/Ca ratio of 0.7, CiQE is internal CO2 concentration for the computation of
with a Ci/Ca ratio of 0.7, CiQE is internal CO2 concentration for the computation of ![]() with a Ci/Ca ratio of 1 and Γ* is CO2 compensation point in absence of dark respiration. The factors 6.5382 and 5.7768 scale
with a Ci/Ca ratio of 1 and Γ* is CO2 compensation point in absence of dark respiration. The factors 6.5382 and 5.7768 scale![]() and
and![]() to 1 at 25°C and 350 ppm CO2, respectively. CiPmax and CiQE are calculated as:
to 1 at 25°C and 350 ppm CO2, respectively. CiPmax and CiQE are calculated as:
| (6) |
| (7) |
where CO2 is atmospheric CO2 concentration (ppm). The Γ* depends on the specificity factor (calculated according to Evans and Farquhar, 1991) and the O2 concentration.
Photosynthesis response to temperature is modeled as Goudriaan and van Laar (1994). Considering a lack of data for chickpea, the Pmax response function to temperature was borrowed from Penning de Veris et al. (1989) based on a fababean (a cool season legume crop like chickpea) photosynthesis response to temperature; Ptmp increases from 0 at 3°C to relative rate of 1 between 14°C and 22.5°C, declines with rising temperature above 22.5°C and reaches to 0 at 35°C. The average temperature during daytime is used to determine the relative response of Pmax to temperature (Ptmp). QE was related to temperature using the function for C3 plants cited in Penning de Veris et al. (1989).
A detailed description of the model equations and source codes of the model are obtainable from the authors.
Test of the Model
To conduct a general test of the model, Pmax was firstly estimated as 0.85 mg CO2 m-2 sec-1 for chickpea cultivars Jam and Kaka using crop growth rate data of K. Ghassemi-Golezani (unpubl. data). For testing of the model, crop growth rate data of Movahhedi (1996) was used. To do this, average daily crop growth rates (g m-2 day-1) were calculated using weekly samples of crop dry matter. In each case meteorological observations of maximum and minimum temperatures and sunshine hours were available and LAI for each day between samplings was estimated using linear interpolation. Then average simulated crop growth rates for different intervals were calculated and compared with observed ones.
For specific test of the model, calculated RUEs at temperatures between 13 and 23°C were compared to the measured values by Soltani et al. (2006) for the same average temperatures. Similarly, percentage increase in RUE calculated for CO2 concentrations of 640 ppm relative to 330 ppm was compared to percentage increase in biomass production measured by Khana-Chopra and Sinha (1987) in three chickpea cultivars at the same CO2 concentrations.
| Table 1: | Typical characteristics of a chickpea crop between flowering and beginning seed growth in Tabriz, NW Iran (K. Ghassemi-Golezani, unpubl. data of 2 years growth analysis of 9 chickpea cultivars at the University of Tabriz, Tabriz, Iran; Movahedi, 1996) |
 | |
| *Estimated using a root/shoot ratio of 0.2 (Gregory, 1988) | |
Simulation for Chickpea
The model was run for chickpea crops at different latitudes and days of year with varying LAI and crop weight. Results showed that RUE is changed little with variation of latitude, day of year and LAI (data not shown), thus a typical chickpea crop between flowering and beginning seed growth stages in Tabriz (38°N), NW Iran, with characteristics presented in Table 1 was considered. In this stage which occurs at day of year 177 (with a range of 170 to 190), vegetative organs are dominant. Long-term (1965-1997) averages of temperature and solar radiation at this time are 25°C and 27.7 MJ m-2 day-1, respectively. The latitude was set at 38°N, which corresponds to the chickpea growing area in NW Iran and the location of the crop presented in Table 1. To generate the response functions of RUE, daily average temperatures from 0 to 40°C and ambient CO2 concentrations from 20 to 1200 ppm were inputted to the model.
RESULTS AND DISCUSSION
Figure 1 shows a plot of simulated versus observed crop growth rates. The crop growth rate data were observed under (and simulated for) average daily temperatures ranged from 11 to 28°C. Good agreement between simulated and observed crop growth rates was obtained.
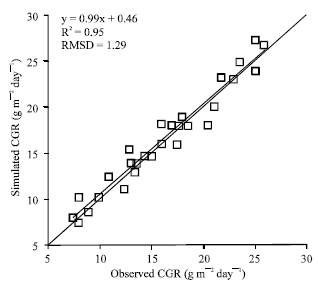 | |
| Fig. 1: | Simulated versus observed Crop Growth Rates (CGR) for chickpea cultivars Jam and Kaka. The solid line is 1:1 line and the thick line is the regression line |
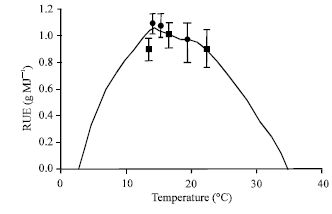 | |
| Fig. 2: | Simulated curve of RUE response to temperature and experimental RUE values. Squares are RUE values from Exp. 1 and circles from Exp. 2 of Soltani et al. (2006) |
This test show the ability of the model to simulate daily biomass production under a range of temperatures. Of course, it should be noted that under field conditions temperature is normally associated positively with solar radiation and negatively with vapor pressure deficit.
The calculated RUEs for temperatures of 13 to 23°C were similar to that measured by Soltani et al. (2006) in chickpea under the same temperature averages (Fig. 2). Soltani et al. (2006) measured RUE in 2 experiments each with 4 plant densities and 3 sowing dates at Gorgan in the west part of the Caspian Sea Coast of Iran. There was no significant difference between sowing densities, but sowing dates differed significantly. Variability of RUE between sowing dates was related to temperature. Khana-Chopra and Sinha (1987) found, in three chickpea genotypes, that with increase in CO2 concentration from 330 to 640 ppm, biomass production increases by 20 to 33%. The model predicts a 25% increase in RUE for the same concentrations, midway between the reported values. From these tests we concluded that model performance is acceptable and can be used to produce response of RUE to temperature and CO2 concentration.
With mean daily temperature of 25°C and radiation of 27.7 MJ m-2 day-1 the crop presented in Table 1 produced 23.8 g DM m-2. RUE was 0.95 g MJ-1 that was obtained with a Pmax of 0.85 mg CO2 m-2 sec-1. This value of RUE is also equal to that reported for chickpea by Hughes et al. (1987).
Figure 2 shows the dependence of RUE to average daily temperature. At temperatures lower than 3°C and higher than 36°C RUE is zero. RUE is sharply increased (9.5%°C-1) with increasing temperature from 3 to 14°C, where it has maximum of its value. With further increase in temperature to 22°C, RUE is slowly decreased (1.4%°C-1). Rising temperature from 22 to 36°C is resulted in the sharp decrease (7.4%°C-1) of RUE. This sharp decrease is a result of increasing respiration and declining photosynthesis with increase in temperature.
Figure 3 shows response to atmospheric CO2 of RUE in chickpea. At CO2 concentrations lower than 60 ppm, RUE is zero, a similar value presented by Wolfe (1994) for net photosynthesis response to CO2. With increasing CO2, RUE is increased by a diminishing rate and saturated at CO2 greater than 700 ppm. This response also is very similar to that reported for C3 plants (net photosynthesis) by Wolfe (1994). Figure 3 shows that a doubling atmospheric CO2 (from 350 to 700 ppm) increases RUE of chickpea by 23%. Kimball et al. (2002) summarizing data from experimental studies, found a 25% increase in growth rate with a doubling of CO2 concentration. Melkonian et al. (1998), using three formulations for estimating the impact of elevated CO2 on daily net canopy carbon assimilation, found that with doubling CO2 concentration 15-48% increase in net carbon assimilation is occurred.
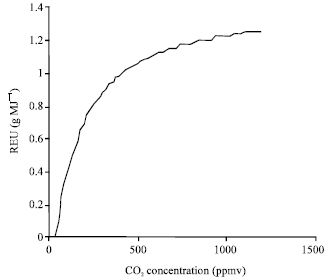 | |
| Fig. 3: | Simulated Radiation Use Efficiency (RUE) of chickpea as a function of atmospheric CO2 concentration |
Overall, by integrating quantitative relationships that capture the physiological responses, a simple, suitable theoretical framework was extended to allow the effects of temperature and CO2 on RUE. By using the framework, response functions of RUE to temperature and atmospheric CO2 concentration were calculated for chickpea. The RUE response to temperature and CO2 concentration is consistent with the results obtained for some other C3 crops. These functions can be used for describing the response of RUE to temperature and CO2 concentration in climate change studies.
REFERENCES
- Choudhury, B., 2000. A sensitivity analysis of the radiation use efficiency for gross photosynthesis and net carbon assimilation by wheat. Agric. For. Meteorol., 101: 217-234.
Direct Link - Hughes, G., J.D.H. Keating, P.J.M. Cooper and N.F. Dee, 1987. Solar radiation interception and utilization by chickpea (Cicer arietinum) crops in Northern Syria. J. Agric. Sci., 108: 419-424.
CrossRef - Kimball, B.A., K. Kobayashi and M. Bindi, 2002. Responses of agricultural crops to free air CO2 enrichment. Adv. Agron., 77: 293-368.
CrossRef - Melkonian, J., S.J. Riha and D.S. Wilks, 1998. Simulation of elevated CO2 effects on daily net canopy carbon assimilation and crop yield. Agric. Syst., 58: 87-106.
CrossRefDirect Link - Saxena, N.P., M.C. Saxena, C. Johansen, S.M. Virmani and H. Harris, 1996. Future Research Priorities for Chickpea in Wana and Sat. In: Adaptation of Chickpea in the West Asia and North Africa Region, Saxena, N.P., M.C. Saxena, C. Johansen, S.M. Virmani and H. Harris (Eds.). ICARDA, Aleppo, Syria, pp: 257-263.
- Sinclair, T.R. and T. Horie, 1989. Leaf nitrogen, photosynthesis and crop radiation use efficiency: A review. Crop Sci., 29: 90-98.
Direct Link - Soltani, A., K. Ghassemi-Golezani, F.R. Khooie and M. Moghaddam, 1999. A simple model for chickpea growth and yield. Field Crops Res., 62: 213-224.
CrossRefDirect Link - Soltani, A., M.J. Robertson, A. Rahemi-Karizaki, J. Poorreza and H. Zarei, 2006. Modeling biomass accumulation and partitioning in chickpea (Cicer arietinum L.). J. Agron. Crop. Sci., 192: 379-379.
Direct Link - Yu, G., T. Kobayashi, J. Zhuang, Q. Wang and L. Qu, 2003. A coupled model of photosynthesis-transpiration based on the stomatal behavior for maize grown in the field. Plant Soil, 249: 401-416.
Direct Link