
M. Sridevi
Department of Microbiology, Acharya Nagarjillill University, Nagarjnna Nagar, Guntur (Dt), Andhra Pradesh-522 510, India
K. V. Mallaiah
Department of Microbiology, Acharya Nagarjillill University, Nagarjnna Nagar, Guntur (Dt), Andhra Pradesh-522 510, India
N. C.S. Yadav
Department of Microbiology, Acharya Nagarjillill University, Nagarjnna Nagar, Guntur (Dt), Andhra Pradesh-522 510, India
Journal of Plant Sciences
Year: 2007 | Volume: 2 | Issue: 6 | Page No.: 635-639







ABSTRACT
Rhizobium species isolated from Crotalaria species (C. junceaC. laburnifoliaC. retusa and C. verrucosa) were tested for their ability to solubilize tricalcium phosphate (TCP). Among these four species, Rhizobium sp. isolated from Crotalaria retusa and C. verrucosa showed zone of TCP solubilization on Pikovskaya`s agar medium. Highest solubilization index (2.70) was observed in Rhizobium sp. from C. retusa. Effect of different carbon and nitrogen sources was also studied. Among the carbon sources, glucose was found to be best carbon source for TCP solubilization. Presence of highest concentration (3%) of glucose enhanced TCP solubilization. Among the nitrogen sources, ammonium sulphate was the best promoter for the process. Addition of low levels (0.5µg mL-1) of EDTA increased TCP solubilization.
PDF Abstract XML References
How to cite this article
M. Sridevi, K. V. Mallaiah and N. C.S. Yadav, 2007. Phosphate Solubilization by Rhizobium Isolates from Crotalaria Species. Journal of Plant Sciences, 2: 635-639.
DOI: 10.3923/jps.2007.635.639
URL: https://scialert.net/abstract/?doi=jps.2007.635.639
DOI: 10.3923/jps.2007.635.639
URL: https://scialert.net/abstract/?doi=jps.2007.635.639
INTRODUCTION
Plant Growth Promoting Rhizobacteria (PGPR) are involved with host plants in mutual interaction. They promote plant growth by the production of phytohormones, solubilization of insoluble phosphates and biocontrol of host plant diseases or improvement of plant nutritional status (Deshwal et al., 2003). Rhizobia are the first group of bacteria, which are attributed to the ability of PGPR to solubilize insoluble phosphates (Johri et al., 2003). So far only a few reports on phosphate solubilizing ability of Rhizobium strains (Halder et al., 1990, 1991; Halder and Chakrabarty, 1993; Rivas et al., 2006; Daimon et al., 2006).
Crotalaria is one of the largest genera of Fabaceae with more than 500 species commonly occurring in diverse climatological situations (Allen and Allen, 1981). Some Crotalaria species are of great agronomic interest, since they are used as used as green manure to improve soil fertility or control nematode populations in infested soils (Sy et al., 2001). Because of its importance the present study was taken up to investigate phosphate solubilizing ability of Rhizobium isolates from commonly occurring Crotalaria species (C. juncea L., C. laburnifolia L., C. retusa L. and C. verrucosa L.) and the ability of these isolates to solubilize tricalcium phosphate (TCP) in the presence of different carbon sources, nitrogen sources and cell affecting agents was also studied.
MATERIALS AND METHODS
Organism and Phosphate Solubilization
Rhizobium species isolated from root nodules of C. juncea, C. laburnifolia, C. retusa and C. verrucosa on Yeast Extract Mannitol Agar (YEMA) medium. The identification of the isolates as Rhizobium was confirmed by plant infection test (Vincent, 1970). The phosphate solubilizing ability of the Rhizobium species was tested on Pikovskaya’s agar (Pikovskaya, 1948) containing tricalcium phosphate (TCP) as insoluble phosphate source. The formed halo zone surrounding the colony revealed phosphate solubilization and was expressed in solubilization index (Arun and Sridhar, 2005). The Rhizobium species, which formed zone of solubilization on agar medium, was further tested in Pikovskaya’s broth having initial pH 7.0.
After incubation the final pH and phosphorus (P) content of the medium were also estimated in culture supernatant (Subba Rao, 1993).
Effect of Carbon and Nitrogen Sources
Effect of different carbon sources (1%) on phosphate solubilization was tested by replacing glucose with 14 carbon compounds (mannitol, galactose, fructose, rhamnose, arabinose, trehalose, inositol, raffinose, sucrose, maltose, mannose, dulcitol, ribose and xylose) sterilized separately and added aseptically to the medium before sterilization. The effect of different concentrations (1.0, 1.5, 2.0, 2.5 and 3.0%) of most effective carbon source was also studied. Effect of different nitrogen sources (0.1%) on TCP solubilization was studied by replacing ammonium sulphate with five different nitrogen sources (sodium nitrate, potassium nitrate, calcium nitrate, asparagine and urea).
Effect of EDTA
The chelating effect of EDTA on TCP solubilization was tested by adding different concentrations (0.5, 1.0, 2.0, 2.5 and 3.0 μg mL-1) of EDTA to the medium.
Statistical Analysis
The data were statistically analyzed by correlation coefficient and ANOVA (two way classification technique), using Statistica software.
RESULTS AND DISCUSSION
The Rhizobium species isolated from C. juncea, C. laburnifolia, C. retusa and C. verrucosa were fast growers as they produced acid in YEM broth and colony diameter was greater than 2.0 mm. Among these species, the Rhizobium species from C. retusa and C. verrucosa formed clear zones around the colonies on Pikovskaya’s agar medium after 3 days of incubation and it gradually increased up to 9 days. The colony diameter is almost similar throughout the incubation period. Solubilization Index (SI) of these species on solid media ranged between 2.4 and 2.7 (Table 1). Maximum SI was observed in Rhizobium sp. from C. retusa (2.7). In liquid medium maximum solubilization was also recorded in this species (840 μg mL-1).
| Table 1: | Solubilization of tricalcium phosphate by Rhizobium species from Crotalaria retusa and C. verrucosa |
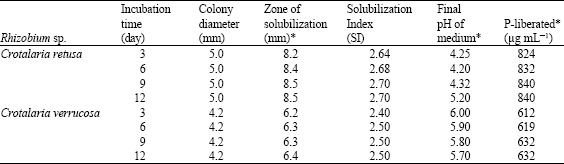 | |
| Solubilization Index (SI) = Diameter of the colony+Halo zone/Diameter of the colony; *: Correlation coefficient (r); Between pH and P liberated = -1; Between zone of solubilization and P liberated = 1 | |
A fall in pH accompanied phosphate solubilization, due to production of organic acids was observed up to 9 days, but there is sudden increase in pH after 9 days. This may be due to utilization of organic acids produced during phosphate solubilization by the strains as reported earlier in Pseudomonas (Dave and Patel, 1999). The data were statistically analyzed using correlation coefficient and it was found that there is positive correlation between zone of solubilization on agar medium and liberated P in broth and negative correlation between liberated P and final pH of the medium.
Effect of different carbon sources (1%) on phosphate solubilizing activity by the Rhizobium sp. revealed that the maximum solubilization occurred in glucose containing medium (Table 2). That the glucose was the best carbon source for phosphate solubilization was also reported earlier in Bradyrhizobium sp. isolated from Cicer arietinum (Halder et al., 1991). Maximum decrease in pH was recorded in glucose containing medium. In other carbon sources little decrease in pH and no correlation between acidic pH and P liberated were observed. The relative efficiency of Rhizobium spp. on different carbon sources could be due to the organic acids secreted by the isolates rather than the total acidity (Dave and Patel, 1999). The data on effect of carbon sources on TCP solubilization by Rhizobium spp. were statistically analyzed using analysis of variance (two way classification technique) and it was found that variations due to both carbon sources and Rhizobium spp. were found to be significant and the Rhizobium spp. differ significantly with different carbon sources.
As the glucose at 1% concentration (as in Pikovskaya’s medium) supported maximum solubilization of TCP, effect of different concentrations of glucose was studied. Rhizobium sp. from C. retusa and C. verrucosa showed maximum solubilization at 3.0% concentration of glucose (Table 3). Linear increase in TCP solubilization was observed as the concentration of glucose increased. This may be due to greater acidity produced at 3.0% concentration of glucose, which is an important factor in phosphate solubilization. The variation in the effect of different concentrations of glucose is found to be statistically significant.
| Table 2: | Effect of different carbon sources on TCP solubilization |
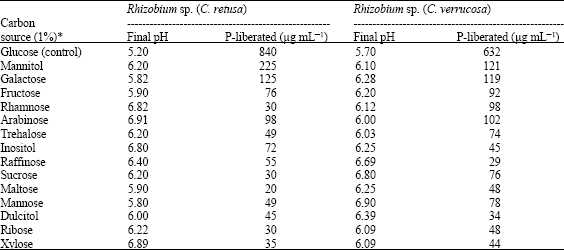 | |
| *: Significant at 1% | |
| Table 3: | Effect of different concentrations of glucose on TCP solubilization |
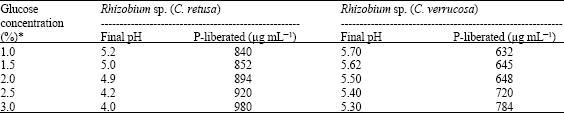 | |
| *: Significant at 1% | |
| Table 4: | Effect of nitrogen sources on TCP solubilization |
 | |
| *: Significant at 1% | |
| Table 5: | Effect of EDTA on TCP solubilization |
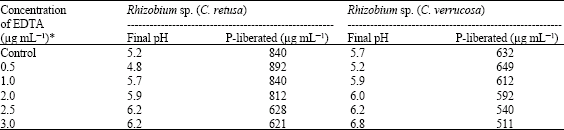 | |
| *: Significant at 1% | |
Effect of different nitrogen sources on TCP solubilization revealed that maximum solubilization of TCP occurred in ammonium sulphate containing medium (Table 4) and reduced the pH of the medium. Further it was observed that the inorganic nitrogen sources supported better solubilization of TCP than organic nitrogen sources. This could be due to the production of inorganic acids by proton exchange mechanism in presence of NH4+ cause accelerated phosphate solubilization (Halder et al., 1991). Statistical analysis showed that the effect of different nitrogen sources on TCP solubilization was also found to be significant.
The effect of different concentrations of EDTA (0.5, 1.0, 2.0, 2.5 and 3.0 μg mL-1) revealed that the addition of 0.5 μg mL-1 EDTA to the standard medium results in significant increase of TCP solubilization, but increasing concentration of EDTA had little effect on TCP solubilization (Table 5). This could be due to the chelation of Ca2+ produced during TCP solubilization as reported in Bradyrhizobium sp. (Halder et al., 1991). The effect of EDTA on TCP solubilization was found to be statistically significant.
Only a few reports on phosphate solubilization have been reported earlier (Halder et al., 1990; 1991; Halder and Chakrabarty, 1993; Rivas et al., 2006; Daimon et al., 2006) and studies were confined to only limited number of hosts and a single species of Rhizobium from a single host. From this it may concluded that, in addition to symbiotic nitrogen fixation, some species of Rhizobium can also involved in phosphate solubilization. Furthermore, the capacity of phosphate solubilization by the Rhizobium species can be exploited as PGPR.
REFERENCES
- Arun, A.B. and K.R. Sridhar, 2005. Growth tolerance of rhizobia isolated from Sand Dune legumes of Southwest coast of India. Eng. Life Sci., 5: 134-138.
Direct Link - Daimon, H., K. Nobuta, M. Ohe, J. Harada and Y. Nakayama, 2006. Tricalcium phosphate solubilization by root nodule bacteria of Sesbania cannabina and Crotalaria juncea. Plant Prod. Sci., 9: 388-389.
Direct Link - Deshwal, V.K., R.C. Dubey and D.K. Maheshwari, 2003. Isolation of plant-growth promoting strains of Bradyrhizobium (Arachis sp.) with biocontrol potential against Macrophomina phaseolina causing charcoal rot of peanut. Curr. Sci., 84: 443-448.
Direct Link - Halder, A.K. and P.K. Chakrabarty, 1993. Solubilization of inorganic phosphate by Rhizobium. Folia Microbiol., 38: 325-330.
Direct Link - Johri, B.N., A. Sharma and J.S. Virdi, 2003. Rhizobacterial diversity in India and its influence on soil and plant health. Adv. Biochem. Eng. Biotechnol., 84: 49-89.
Direct Link - Rivas, R., A. Peix, P.F. Mateos, M.E. Trujillo, E. Martinez-Molina and E. Velazquez, 2006. Biodiversity of populations of phosphate solubilizing rhizobia that nodulates chickpea in different Spanish soils. Plant Soil, 287: 23-33.
CrossRefDirect Link - Sy, A., E. Giraud, P. Jourand, N. Garcia and A. Willems et al., 2001. Methylotrophic Methylobacterium bacteria nodulate and fix nitrogen in symbiosis with legumes. J. Bacteriol., 183: 214-220.
CrossRefDirect Link