
Nishi Mathur
Department of Botany, Microbial Biotechnology and Biofertilizer Laboratory, J.N.V. University, Jodhpur (Rajasthan)-342 001, India
Joginder Singh Sachendra Bohra
Department of Botany, Microbial Biotechnology and Biofertilizer Laboratory, J.N.V. University, Jodhpur (Rajasthan)-342 001, India
Afshan Quaizi
Department of Botany, Microbial Biotechnology and Biofertilizer Laboratory, J.N.V. University, Jodhpur (Rajasthan)-342 001, India
Anil Vyas
Department of Botany, Microbial Biotechnology and Biofertilizer Laboratory, J.N.V. University, Jodhpur (Rajasthan)-342 001, India
Journal of Plant Sciences
Year: 2007 | Volume: 2 | Issue: 2 | Page No.: 127-140







ABSTRACT
Phytoremediation as a means of cleaning up polluted soils has gained popularity during the last decade due to its convenience and low costs of installation and maintenance. When the target pollutant is biodegradable, this technology exploits the stimulating effect that roots have on microbial processes and physical/chemical modifications in the rhizosphere. Among the microorganisms that affect rhizosphere processes, symbiotic fungi forming mycorrhizas induce a series of changes in plant physiology, nutrient availability and microbial composition that may determine the outcome of a phytoremediation attempt. Beyond the rhizosphere, mycorrhizal hyphae act as the roots of the roots and may thus extend the rhizosphere into the bulk soil by creating a new interface of soil-plant interactions: the hyphosphere. Isolation of the indigenous and presumably stress-adapted AM fungi can be a potential biotechnological tool for inoculation of plants for successful restoration of degraded ecosystems. This review highlights the potential of AM fungi for enhancing phytoremediation of heavy metal contaminated soils. A great deal of research indicates that plants have the genetic potential to remove many toxic metals from the soil. Despite this potential, phytoremediation is yet to become a commercially available technology. Progress in the field is hindered by a lack of understanding of complex interactions in the rhizosphere and plant-based mechanism which allow metal translocation and accumulation in plant.
PDF Abstract XML References Citation
How to cite this article
Nishi Mathur, Joginder Singh Sachendra Bohra, Afshan Quaizi and Anil Vyas, 2007. Arbuscular Mycorrhizal Fungi: A Potential Tool for Phytoremediation. Journal of Plant Sciences, 2: 127-140.
DOI: 10.3923/jps.2007.127.140
URL: https://scialert.net/abstract/?doi=jps.2007.127.140
DOI: 10.3923/jps.2007.127.140
URL: https://scialert.net/abstract/?doi=jps.2007.127.140
INTRODUCTION
Phytoremediation, the use of plants for environmental restoration, is an emerging cleanup technology. To exploit plant potential to remediate soil and water contaminated with a variety of compounds, several technological subsets have been proposed. Phytoextraction is the use of higher plants to remove inorganic contaminants, primarily metals, from polluted soil (Lasat, 2002). In the approach, plants capable of accumulating high levels of metals are grown in contaminated soil. At maturity, metal-enriched aboveground biomass is harvested and a fraction of soil metal contamination removed. Plants have a natural propensity to take up metals.
Phytoremediation utilizes biological organisms for phytoextaraction or removal of plant biomass containing concentrated levels of heavy metals taken up from polluted soils. Phytoremediation is an alternative to conventional chemical and methods of treating contaminated soils.
Essential heavy metals include zinc (Zn), Copper (Cu), manganese (Mn), Cobalt (Co), but no biological functions in plants are currently known for cadmium (Cd), lead (Pb), mercury (Hg), or chromium (Cr). Chromium is a heavy metal with risk to human health. Its presence in the agricultural soils can be attributed to the use of organic wastes as fertilizer and the use of wastewater for irrigation. Sites for wood-treatment/preservation, steel processing, aluminum alloys, electroplating and microbial growth inhibition such as cooling towers of power plants, may be contaminated with Cr.
| Table 1: | Summary of AM fungi-heavy metal interactions and application in phytoremediation |
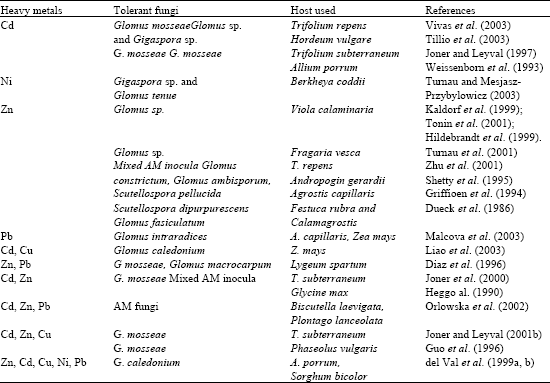 | |
Heavy metals are not degraded biologically in soil. In soils they occurs as free metal ions, exchangeable metal ions, soluble metal complexes (sequestered to ligands), organically bound metals, precipitated or insoluble compounds such as oxides, carbonate and hydroxides, or they may form part of the silicate structure (indigenous soil content). Mycorrhizal fungi have greatest impact on elements with narrow diffusion zones around plant roots, including heavy metals and phosphorus (P). Mycorrhiza enhances the uptake of Cu, Zn, Ni, Cd, Pb and other metals (Table 1).
Mycorrhiza increase nutrient and water uptake, alleviate cultural and environmental stresses and enhances disease resistance and plant health. An important arbuscular mycorrhizal genus is Glomus, which colonize a variety of hot species, including sunflower.
Mycorrhizal fungi are a direct link between soil and roots and consequently of great importance in phytoremediation-potentially enhancing heavy metals availability and plant tolerance (Gaur and Adholeya, 2004). Little is known of the ability of mycorrhizal fungi to enhance plant tolerance and phytoaccumulation of Cr.
ROLE OF MYCORRHIZAE IN PHYTOREMEDIATION
Since heavy metal uptake and tolerance depend on both plant and soil factors including soil microbes, we require information on interactions between plant root and their symbionts such as Arbuscular Mycorrhizal (AM) fungi and nitrogen- fixing microbes. It is the generally held view that the majority of plants growing under natural conditions have mycorrhizae (Smith and Reed, 1997; Mathur and Vyas, 1997). Mycorrhizal colonization of roots results in an increase in root surface area for nutrient acquisition. The extramatrical fungal hyphae can extend several cm into the soil and uptake large amounts of nutrients, including heavy metals, to the host root. The effectiveness of AM root colonization in terms of nutrient acquisition differs markedly between AM fungi and host plant genotype (Bohra et al., 2005; Mathur and Vyal, 2004; Ahiabor and Hirata, 1995; Marschner, 1995).
Mycorrhizae have also been reported in plants growing on heavy metal contaminated sites (Pawloska and Charvat, 2004; Shetty et al., 1995; Chaudhry et al., 1980) indicating that these fungi have evolved a HM- tolerance and that they may play a role in the phytoremediation of the site. Noyd et al. (1996) reported that AM fungal infectivity of native prairie grasses increased over three seasons on a coarse taconite iron ore tailing plots which helped to establish a sustainable native grass community that will meet reclamation goals. The reported symbiotic associations in the plants colonizing heavy metal contaminated soils further suggests a selective advantage for these plants as pioneering species on such sites and that they may be largely responsible for the successful colonization of such habitats.
Various authors have reported isolating spores of arbuscular mycorrhizal fungal texa such as Glomus and Gigaspora associated with most of the plants growing in heavy metal polluted habitats (Chaudhry et al., 1980; Raman and Sambandan, 1998; Raman et al., 1993). Raman et al. (1993) identified Glomus and Gigaspora sp. in the mycorrhizospheres of fourteen plant species colonising a magnesite mine spoil in India. Whereas Weissenhorn and Leyval, 1995) isolated only Glomus mosseae and Dueck et al. (1986) isolated Glomus fasciculatum alone from the heavy metal polluted soils. Pawlowska et al. (1996) surveyed a calamine spoil mound rich in Cd, Pb and Zn in Poland and recovered spores of Glomus aggregatum, G. fasciculatum and Entrophospora spp. from the mycorrhizospheres of the plants growing on spoil. Galli et al. (1994) suggested that mycorrhizae can playa crucial role in protecting plant roots from heavy metals. The efficiency of protection, however, differs between distinct isolates of mycorrhizal fungi and different heavy metals. Joner and Leyval (1997) reported that extra-radical hyphae of AM fungus Glomus mosseae can transport Cd from soil to subterranean clover plants growing in compartmented pots, but that transfer from fungus to plant is restricted due to fungal immobilization. The authors also reported no restriction of fungal hyphal growth into soil with high extractable Cd levels. Our preliminary studies have also showed very little, if any, translocation of Zn absorbed by mycorrhizal maize seedlings grown in contaminated soil, to the shoots. Turnau (1998) studied the localization of heavy metals within the fungal mycelium and mycorrhizal roots of Euphorbia cyparissias from Zn contaminated wastes and found higher concentrations of Zn as crystaloids deposited within the fungal mycelium and cortical cells of mycorrhizal roots. Studies by various researchers (Galli et al., 1994; Leyval et al., 1995; Hetrick et al., 1994) have shown that mycorrhizal fungal ecotypes from heavy metal contaminated sites seem to be more tolerant to heavy metals (and have developed resistance) than reference strains from uncontaminated soils.
Galli et al. (1995) reported that although there was an increase in the contents of cystein, gamma EC and GSH in the mycorrhizal maize roots grown in quart sand with added Cu, no differences in Cu uptake were detected between non-mycorrhizal and mycorrhizal plants. These results do not support the idea that AM fungi protects maize from Cu-toxicity. Mycorrhizae are also known to produce growth- stimulating substances for plants, thus encouraging mineral nutrition and increased growth and biomass necessary for phytoremediation to become commercially viable strategy for decontamination of polluted soils.
For arbuscular mycorrhizae the results are conflicting. Some reports indicate higher concentrations of heavy metals in plants due to AM, even resulting in toxic levels in plants (Weissenhorn and Leyval, 1995; Joner and Leyval, 1997; Killham and Firestone, 1986) (Table 2).
whereas others have found a reduced plant concentrations of, e.g. Zn and Cu in mycorrhizal plants (Schuepp et al., 1987; El-Kherbawy, 1989; Heggo, 1990). Diaz et al. (1996) studied influence of Zn and Pb uptake by Lygeum spartum and Anthyllis cytisoides plants inoculated with Glomus mosseae and G. macrocarpum AM fungi in soils with different levels of these metals. The authors showed that, at low doses, mycorrhizal plants had equal or higher Zn or Pb concentrations than non-mycorrhizal controls; at higher doses, however, metal concentrations in the plants inoculated with G. mosseae were lower than those found in the corresponding controls, while the plants inoculated with G. macrocarpum showed similar or even higher levels than the controls.
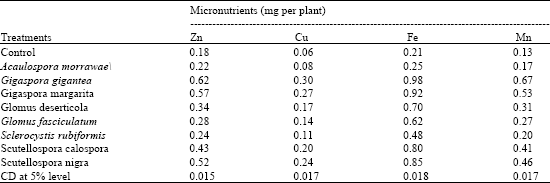
| Table 2: | Influence of different AM fungi on micronutrient uptake in Anogeissus latifolia |
 | |
In addition to the damaging effects on plants, the effect of heavy metals on the soil microorganisms and soil microbial activity also need to be considered. The impact of heavy metals in the field on Rhizobium leguminosarum bv. Trifolii and AM were estimated by various workers (Mench et al., 1994; Weissenhorn and Leyval, 1995). A negative effect of Zn on the indigenous rhizobial population was suggested by Mench et al. (1994). On the contrary, no adverse effect was found on spore number and mycorrhizal colonization of maize (Weissenhorn et al., 1992). Various soil factors such as the clay contents and mobility of heavy metals effect plants and soil biota. As metal uptake by plant roots depends on soil and their associated symbionts, it is important to monitor metal mobility and availability to plant and its symbionts when assessing the effect of soil contamination on plant uptake and related phytotoxic effects.
The prospect of symbionts existing in heavy metal contaminated soils has important implications for phytoremediation. Mycorrhizal associations increase the absorptive surface area of the plant due to extra-matrical fungal hyphae exploring rhizospheres beyond the root hair zone, which in turn enhance water and mineral uptake. The protection and enhanced capability of greater uptake of minerals result in greater biomass production, a pre-requisite for successful remediation. The potentials of phytoremediation of contaminated soil can be enhanced by inoculating hyperaccumulator plants with mycorrhizal fungi most appropriate for contaminated site.
POTENTIAL OF AMF FOR PHYTOREMEDIATION OF HEAVY METAL POLLUTED SOILS
Soil microorganisms are known to playa key role in the mobilization and immobilization of metal cations, thereby changing their availability to plants (Birch and Bachofen 1990). AMF are among the most common soil microorganisms and constitute an important functional component of the soil-plant system occurring in almost all habitats and climates (Barea et al., 1997), including disturbed soils (Mc Gonigle and Miller, 1996; Brundrett et al., 1996). Degraded soils do, however, suffer from changes in diversity and abundance of AM fungal populations (Koomen et al., 1990; Jasper et al., 1991; Loth, 1996).
More specifically, it has been shown that AMF can be affected by heavy metal toxicity, but in many cases mycotrophic plants growing in soils contaminated with heavy metals are colonized by AMF (Leyval et al., 1997). Many reports concerning this have quantified spores and estimated root colonization in situ. Others have gone further and described metal tolerant AMF in heavy metal polluted soils (Gildon and Tinker 1983; Weissenhorn et al., 1995; del Val 1999a; Hildebrandt et al., 1999).
In the last few years research interest has focused on the diversity and tolerance of AMF in heavy metal contaminated soils trying to understand the basis underlying adaptation and tolerance of AMF to heavy metals in soils, since this could facilitate the management of these soil microorganisms, for restoration/bioremediation programs. Vandenkoornhuyse (1998) showed that AMF species diversity associated with maize plants in a long-term field experiment did not differ between three plots that had received different levels of heavy metals-containing sewage sludge. However, the number of spores of each species was lower in the soil with the highest concentration of heavy metals. Using the same long-term field experiment and the same plant variety, but more acidic soils, del Val et al. (1999b) found a reduction of number, but also of diversity of AMF spores in the soil receiving the highest rate of sludge.
On a highly polluted soil in Indian thar desert where only adapted plants can grow, Anogeissus latifolia roots were collected along a gradient of heavy metal concentration. Up to three different Glomus species were identified inside Anogeissus roots, which differed along the gradient of metals. The contribution of these AMF to plant tolerance to heavy metals or heavy metal accumulation by plants has not been established (Singh et al., unpublished data). Four Glomus species were also found in the rhizosphere of another metal tolerant plant, Viola calaminaria, growing on a soil highly contaminated with heavy metals (20,961 and 41 mg kg-1 Zn and Cd, respectively) (Tonin et al., 2001). Only one of these fungi colonized clove roots growing in pots supplemented with Cd and Zn salts. This Glomus sp. Increased Cd and Zn concentrations in clover roots, but not in shoots and did not affect plant growth. On the contrary, a Glomus isolate from the rhizosphere of Viola calaminaria increased the growth of maize and lucerne in, heavy metal polluted soils and reduced Zn concentration in roots and shoots (Hildebrandt et al., 1999; Kaldorf et al., 1999).
Although AMF have been recovered from numerous metal enriched habitats, their role in plant interaction with toxic metals is not well understood. At high metal concentrations reports show variations in metal accumulation and inter-plant translocation depending on the fungi, host-plant, root density, soil characteristics, metals and their availability (EI-Kerbawy et al., 1989; Leyval et al., 1997) (Fig. 1) Large variations have also been found between AM fungal species due to differences in hyphal growth outside the rhizosphere (Joner and Leyval 1997). Metal tolerant AMF isolates can decrease metal concentration in shoots or in roots, or decrease translocation from root to shoots (Diaz et al., 1996; Joner and Leyval, 1997). The latter (could be due to the high metal sorption capacity of these fungi, which could 'filter' metal ions during uptake (Joner et al., 2000). The high concentrations of heavy metals in the intracellular hyphae of a heavy metal tolerant AMF colonizing maize roots (Kaldorf et al., 1999) and in phosphate rich material in hyphal vacuoles of mycorrhizal roots of Pteridium aquilinum (Turnau et al., 1993) strengthen the hypothesis of a sequestration of metals by AMF structures.
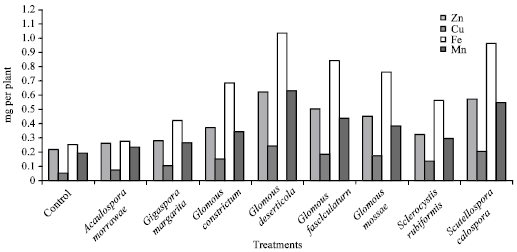 | |
| Fig. 1: | Influence of different AM fungi on micronutrient uptake in Salvadora persica |
However, the competivity of such metal tolerant AMF in the field is often unknown and should be investigated. Further, the potential benefit of a consortium of AMF, which corresponds to the situation in the roots, to improve phytoremediation, should be considered.
Phytoextraction studies often use hyper accumulators (plants accumulating high concentrations of heavy metals, e.g., 1% Zn in their dry matter), which are in most cases non-mycotrophic plants belonging to the Brassicaceae. One objective is to use plants with high concentrations of heavy metals in shoots, which may limit the potential use of AM plants. However, many of these hyperaccumulating plants, such Thlaspi caerulescens, are small and grow slowly, which may limits phytoextraction rates. Other accumulators producing a higher biomass, such as sunflower and willow, are now receiving attention and these are mycorrhizal plants. Highly productive crops associated with metal-tolerant AMF may therefore be considered for decontamination of slightly contaminated soils (Ernst, 2000).
ROLE OF AMF IN PAH-POLLUTED SOILS
Polycyclic Aromatic Hydrocarbons (PAH) are hydrophobic organic molecules consisting of two or more fused benzene rings. A selection of 16 PAH are commonly quantified for characterization and monitoring of these pollutants. While in fact 200-300 PAH compounds and their derivatives are commonly extracted and may putatively be identified in polluted soil samples. The concern for PAH pollution derives from their ubiquitous distribution, their recalcitrance towards degradation and their proven or suspected mutagenic properties I (WHO, 1983). The origin may partially be natural (organic residues after tire), or anthropogenic (mainly processing and incomplete combustion of fossil fuels). Thus, oil spills and industrial sites. e.g., for coke distillation etc. commonly give rise to extreme pollution event!; for which physical, chemical and biological remediation strategies are employed for clean-up. These include, among others, bio-venting, land-farming, bio-augmentation and phytoremediation. The latter is only applicable when pollution levels and physical conditions of the polluted matrix permit the establishment of plants, but offers a cost-effective and efficient treatment that simultaneously restores an ecosystem, limits erosion and improves the esthetical impression of a polluted site (Cunningham et al., 1996; Schwab and Banks, 1994; Wilson and Jones, 1993).
Arbuscular mycorrhiza fungi may playa role in two aspects of bioremediation of PAH: The establishment of a plant cover on polluted soil and modification of PAH degradation rates or pathways.
Improved plant establishment on fallowed, degraded or polluted soils, waste materials or soil-waste mixtures in the presence of AMF is well known (Reddell and Milnes, 1992; Shetty et al., 1994; Thompson, 1994; Leyval and Binet, 1998). The major mechanism behind the success of AM under such conditions is an improvement of plant nutrient acquisition, with additional benefits due to improved water relations, pollutant tolerance and sequestration. The impact of the latter factors are often difficult to distinguish due to the confounding effect of plant nutrition on plant size, but, in the case of enhanced survival in the absence of competitors/predators, the role of AM may be clear. Identification of the symbiotic features that have permitted continued subsistence is however difficult and such investigations are still in their infancy. One mechanism that may be involved is the oxidation of the contaminant by activated oxygen species and concomitant enhancement of oxidoreductases to protect the plant from oxidative stress. Indeed, enhanced levels of hydrogen peroxide in AM roots (Salzer et al., 1999) as well as enhanced levels of peroxidase activity in mycorrhizal roots and the rhizosphere (Criquet et al., 2000) has been demonstrated.
One peculiarity of PAH polluted soil that may be overcome by AM plants is the hydrophobicity and resulting limitations in uptake of water and water- dissolved inorganic nutrients (Leyval and Binet, 1998). Again confounding effects of improved mineral nutrition prevents the distinction of mycorrhizal effects on uptake of water and mineral nutrients, but even if the effect is limit- ed to mineral uptake it is of no less importance.
Effects of AMF on PAH degradation in the rhizosphere may be direct or indirect. As PAH are not absorbed by plants (Binet et al., 2000b; Schwab and Banks, 1994) and are metabolized intracellularly, all degrading activity would take place in soil or inside soil organisms other than AM. Furthermore, AMF have poor saprophytic capacities, so the only probable direct effect of AMF on PAH degradation would be through enhanced production of extracellular peroxidases. Indirect effects would be due to changes in the microbial community e.g. due to stronger competition for mineral nutrients, direct antagonistic or synergistic effects of AMF or changes in root exudation patterns, phenomena that are well documented.
Binet et al. (2000b) found no effect of AM on the dissipation on light compounds like anthracene (3 rings) in spiked soil on the short term (<40 days). Continued phytoremediation treatments of spiked soil resulted in an almost complete (98%) dissipation of anthracene after only 56 days in the rhizosphere of clover/ryegrass (Joner et al., unpublished data). Heavier compounds (4-6 rings) are more persistent and often rely on degradation by co-metabolism rather than direct metabolism (Cutright and Lee, 1991; Perry, 1979). The rhizospheric effect on dissipation of these compounds is thus more pronounced than for the lighter PAH (Binet et al., 2000a) as the root exudates may drive co-metabolism. Mycorrhizal effects on PAH dissipation was in this case accompanied by a modification in biomass partitioning (but not total plant biomass) between two co-occurring plant species (clover and ryegrass) (Joner and Leyval, 2001b), as well as changes in soil microbial community structure. The effect of colonization by mycorrhizal fungi on the competitive ability of clover on PAH-polluted soil (Joner and Leyval, 2001) is perhaps similar to that observed in non- polluted soil (Bolan et al., 1987), i.e., a phenomenon linked to the improved nutrient acquisition of the mycotrophic legume. The modification of the soil microbial community may on the other hand either be due to inherent qualitative (Germida et al., 1998) or quantitative (Griffiths et al., 1999) differences in root exudation between the two plant species coupled with a proportional change in plant biomass (indirectly mediated by the mycorrhizal effect on competitive ability/plant size of the two plants), or more direct effects of AMF, i.e., alterations in root exudation as a consequence of AMF colonization (Graham et al., 1981; Laheurte et al., 1990) and direct antagonistic/synergistic effects between the AMF hyphae and the soil microtlora (Linderman, 1988, 1991). The exploitation of root-free soil by AMF hyphae has a potential for modifying microbial composition (Olsson et al., 1996; Ravnskov et al., 1999) and activity. Ephemeral hyphae serving as a source of C outside.
POTENTIAL USE OF AMF IN BIOASSAYS FOR SOIL POLLUTION
Arbuscular mycorrhizas are not only an aid to ecosystem remediation. They should also be considered as a key indicator for soil pollution or soil quality because:
| • | mycorrhizal fungi are ubiquitous microorganisms, |
| • | many plants are highly dependant on mycorrhizas for their growth, |
| • | they provide a direct link between soil and roots and |
| • | are involved in the transfer of elements including pollutants from soil to plants. |
Furthermore, AMF can be affected by pollutants in soil and can be more sensitive to pollutants than plants (Weissenhom and Leyval, 1995).
The toxicity of compounds such as xenobiotics, PAH and heavy metals on AMF has been studied using techniques based on estimation of spore germination (Weissenhom and Leyval, 1996), mycorrhizal colonisation of roots in pot cultures using nested PCR (Jacquot et al., 1999), mycorrhizal infectivity (Leyval et al., 1995) and mycorrhizal colonization of Ri T-DNA transformed roots (Wan et al., 1998). AMF spore germination in soil can be used as an early indication of the toxicity towards the ecosystem, while mycorrhizal colonisation of roots indicates toxicity at a later stage of the association between symbiotic fungi and roots. AMF can be affected by pollutants but also by soil properties such as pH and phosphorus content (Leyval et al., 1995). The lack of specificity towards heavy metals or other pollutants is often the case for other indicator organisms such as earthworms, algae, fish and plants, which are used for the assessment of ecological risk due to toxic substances. AMF could be included as addition- al indicator organisms in the existing battery of bioassays. There is a need for a standardized technique using arbuscular mycorrhizas as a bioassay, which should be made easier since commercial AMF inoculum is now available. The technology of phytoextraction utilizes hiperaccumulating plants in order to extract heavy metals from the environment. Most hiperaccumulating plants belong to the families Brassicaceae (pl. Thlaspi, Alyssum), Compositae, Euphorbiaceae, Fabaceae, Liliaceae, Scrophulariaceae, Poaceae, Violaceae, which occur also in the native Hungarian flora. Nearly all of them are not agricultural plants, thus their economic utilization is problematic (also often because of the low biomass, the complicated harvesting and propagation). Another problem is the mentioned selectivity of accumulation. Until now there are some new innovations in order to solve these problems, e.g., the heavy metal uptake can be enhanced among larger biomass-producing, but not hiperaccumulating plants with chelating soil treatment like with EDTA (Huang et al., 1997).
In recultivation processes the adaptation mechanism of hiperaccumulating plants is a competitive advantage to be a pioneer and on a longer scale it alters the environment so that weaker or non-tolerant species can distribute. Hence, hiperaccumulating plants offer an environment-friendly way of land decontamination and controlled agro-ecosystem formation. Such succession can be accelerated through the co-application of hiperaccumulating plants and selected metal-tolerant microbes.
Several publications report on the role of ecto- and endomycorrhizal fungi in the recolonization of heavy metal contaminated soils by pioneer plants. Mycorrhizal fungi offer a better nutrient supply, higher metal tolerance for the macrophyte symbiotic partner (Gemma and Koske, 1990; Vörös et al., 1998). The more abundant and more ancient mycorrhizal type is the arbuscular one (AM), which offers a better soil exploitation, higher nutrient and water uptake, thus results in a higher biomass production (Marschner, 1997). AM fungi are among the most abundant soil fungi.
The effect of AM fungi on the heavy metal uptake of the host plant depends on the physical and chemical properties of the contaminated soil (Wang and Chao, 1992; Thompson, 1990; Killham and Fireston, 1986; El-Kherbawy et al., 1989), on the heaviness and duration of the contamination load, on the plant (Griffionen and Ernst, 1989; Kucey and Janzen, 1987) and fungal species (Gildon and Tinker, 1981), thus, the efficiency of the symbiosis. The published data demonstrate that by selected coupling of compatible symbiotic partners plant metal uptake can be altered parallel to higher plant vitality.
The scientific and economic establishment and the necessity of the planned research, research-innovation is proved by our previous experimental results and the country's environmental problems.
Inorganic contaminants like heavy metals are not biodegradable thus possible management like phytoremediation aims in the decreasing of soil heavy metal content (by extraction), parallel to the prevention of food chain contamination.
Based on toxicity, level of contamination and ecological conditions heavily contaminated (lethal for most organisms) and contaminated (tolerable) sites are differentiated. Level of contamination changes in time and this dynamics is affected by the successive order of the applied phytoremediation methods.
According to these we plan the development of phytoremediation methods (and method-combinations) that could be applied together in space and successively in time:
Cultivation of Heavy Metal Accumulating Plants (On Tolerable Contaminated and Heavily Contaminated Sites)
An important aim in the application of hiperaccumulating plants to maximize the level and speed of metal uptake. Important factor is the allocation of uptake metals into aboveground organs that can be easier harvested and further incinerated. The rate of the heavy metal extraction can be increased by the choice of the plant species, through intraspecific selection, by the application of uptake-enhancing additives (Deram et al., 2000), by increasing plant biomass (through agricultural techniques), by increasing plant life cycles that are active in heavy metal uptake (e.g., rotation of species with different temperature optima).
Cultivation of Heavy Metal Accumulating Plants That Are Able to Form Mycorrhizal Partnership (On Heavily Contaminated Sites)
The choice of plants based on their ability to form symbiosis with AM fungi. Coupling with proper fungal partner has positive effect on water and nutrient uptake and thus on biomass production. Coupling of such plant and fungal species in which symbiotic fungal partner enhance metal uptake in order to extract higher amount of contaminant.
Cultivation of Non-Hiperaccumulating but Mycorrhizal Plants That Produce Large Amount of Biomass (On Tolerable Contaminated Sites)
In the choice of the proper plant and fungal partners it is important to have a positive mycorrhizal effect on metal uptake (controlled mycorrhization with heavy metal uptake enhancing AM fungi).
Cultivation of non-accumulating, but highly mycorrhizal plants (on weakly contaminated sites).
The last step of a phytoremediation technology is the one when metal contamination has decreased or on a weakly contaminated site the heavy metal entrance to food chains should be prevented (e.g., grazing wild animals). At this step a negative AM-fungal effect on metal uptake is beneficial, in order to decrease the amount of metals entering the food chain.
REFERENCES
- Malcova, R., M. Voslitka and M. Gryndler, 2003. Effects of inoculation with Glomus intraradices on lead uptake by Zea mays L. and Agrostis capillaris L. Applied Soil Ecol., 23: 55-67.
Direct Link - Mathur, N. and N. Vyas, 1997. Vesicular arbuscular mycorrhizae in Indian thar desert. J. Indian Bot. Soc., 76: 125-126.
Direct Link - McGonigle, T.P. and M.H. Miller, 1996. Development or fungi below ground in association with plants growing in disturbed and undisturbed soil. Soil Biol. Biochem., 28: 263-269.
Direct Link - Mench, M.J., V.L. Didier, M. Loffler, A. Gomez and P. Masson, 1994. A mimicked in-situ remediation study of metal contaminated clay soil with emphasis on cadmium and lead. J. Environ. Qual., 28: 58-63.
Direct Link - Noyd, R.K., F.L. Pfleger and M.R. Norland, 1996. Field responses to added organic matter, arbuscular mycorrhizal fungi and fertilizer in reclamation of torbonite iron ore tailing. Plant Soil 179: 89-97.
Direct Link - Olsson, P.A., K. Baath, I. Jakosen and B. Soderstrom, 1996. Soil bacteria respond to respond to presence of roots but not to mycelium of arbuscular mycorrhizal fungi. Soil Biol. Biochem., 28: 463-470.
Direct Link - Orlowska, E., S. Zubek, A. Jurkiewicz, G. Szarek-Ukaszewska and K. Tumau, 2002. Influence of restoration on arbuscular mycorrhiza of Biscutella laevigata L. (Brassicaceae) and Plantago lanceolata L. (Plantaginaceae) from calamine spoil mounds. Mycorrhiza, 12: 153-159.
Direct Link - Pawlowska, T.E., J. Blaszkowski and A. Ruhling, 1997. The mycorrhizal status of plants colonizing a calamine spoil mound in Southern Poland. Mycorrhiza, 6: 499-505.
CrossRefDirect Link - Pawlowska, T.E. and I. Charvat, 2004. Heavy-metal stress and development patterns of arbuscular mycorrhizal fungi. Applied Environ. Microbiol., 70: 6643-6649.
Direct Link - Raman, N. and S. Sambandan, 1998. Distribution of V AM fungi in tannery effluent polluted soils of Tamil Nadu, India. Bull. Environ. Contamin. Toxicol., 60: 142-150.
Direct Link - Raman, N., N. Nagarajan, S. Gopinathan and K. Sambandan, 1993. Mycorrhiza1 status of plant species colonizing a magnesite mine spoil in India. Biol. Fertil. Soil, 16: 76-78.
Direct Link - Ravnskov, S., O. Nybroe and I. Jakobsen, 1999. Influence or an arbuscular mycorrhizal fungus on Pseudomonas f1uorescens D F57 in rhizosphere and hyphosphere soil. New Phytol., 142: 113-122.
Direct Link - Reddell, P. and A.R. Milnes, 1992. Mycorrhizas and other specialised nutrient-acquisition strategies. Their occurrence in woodland plants from Kakadu and their role in rehabilitation of waste rock dumps at a local uranium mine. Aust. J. Bot., 70: 223-242.
Direct Link - Salzer, P., H. Corbiere and T. Boller, 1999. Hydrogen peroxide accumulation in Medicago truncatula roots colonized by the arbuscular mycorrhiza-forming fungus Glomus intraradices. Plama, 208: 319-325.
Direct Link - Shetty, K.G., B.A.D. Hetrick, D.A.H. Figge and A.P. Schwab, 1994. Effects of mycorrhizae and other soil microbes on revegetation of heavy metal contaminated mine spoil. Environ. Pollut., 86: 181-188.
CrossRefDirect Link - Shetty, K.G., B.A. Hetrick and A.P. Schwab, 1995. Effects of mycorrhizae and fertilizer amendments on zinc tolerance of plants. Environ. Pollut., 88: 307-314.
Direct Link - Thompson, J.P., 1990. Soil sterilization methods to show VA-mycorrhizae aid P and Zn nutrition of wheat in vertisols. Soil Biol. Biochem., 22: 229-240.
Direct Link - Thompson, J.P., 1994. Inoculation with vesicular-arbuscular mycorrhizal fungi from cropped soil over- comes long-fallow disorder of linseed (Linum usitatissimum L.) by improving P and Zn uptake. Soil Biol. Biochem., 26: 1133-1143.
Direct Link - Tonin, C., P. Vandenkoornhuyse, E.J. Loner, I. Straczek and C. Leyval, 2001. Assessment of arbuscular mycorrhizal fungi diversity in the rhizosphere of Viola calaminaria and effect of these fungi on heavy metal uptake by clover. Mycorrhiza, 10: 161-168.
CrossRefDirect Link - Tullio, M., F. Pierandrei, A. Salemo and E. Rea, 2003. Tolerance to cadmium of vesicular arbuscular mycorrhizae spores isolated from a cadmium-polluted and unpolluted soil. Biol. Fertil. Soils, 37: 211-214.
Direct Link - Tumau, K., 1998. Heavy metal content and localization in mycorrhizal Euphorbia cyparissias from zinc wastes in Southern Poland. Acta Soci. Bot. Poloniae, 67: 105-113.
Direct Link - Tumau, K., P. Ryszka, V. Gianinazzi-Pearson and D. van Tuinen, 2001. Identification of arbuscular mycorrhizal fungi in soils and roots of plants colonizing zinc wastes in Southern Poland. Mycorrhiza, 10: 169-174.
Direct Link - Turnau, K. and J. Mesjasz-Przybylowicz, 2003. Arbuscular mycorrhiza of Berkheya coddii and other Ni-hyperaccumulating members of Asteraceae from ultramafic soils in South Africa. Mycorrhiza, 13: 185-190.
CrossRefDirect Link - Vivas, A., A. Marulanda, M. Gomez, J.M. Barea and R. Azcon, 2003. Physiological characteristics (SDH and ALP activities) of arbuscular mycorrhizal colonization as affected by Bacillus thuringien- sis inoculation under two phosphorus levels. Soil Biol. Biochem., 35: 987-996.
Direct Link - Wan, M.T., J.E. Rahe and R.G. Watts, 1998. A new technique for determining the sublethal toxicity of pesticides to the vesicular-arbuscular fungus Glomus intraradices. Environ. Toxicol. Chem., 17: 1421-1428.
Direct Link - Wang, Y. and C.C. Chao, 1992. Effects of vesicular-arbuscular mycorrhizae and heavy metals on the growth of soybean and phosphate and heavy metal uptake by soybean in major soil groups of Taiwan. J. Agric. Assoc. China. New Ser., 157: 247-256.
Direct Link - Weissenhom, I., C. Leyval and I. Berthelin, 1992. Bioavailablity of heavy metals and abundance of arbuscular mycorrhiza in soil polluted by atmospheric deposition from a smelter. Biol. Fertil. Soils, 19: 22-28.
Direct Link - Weissenhorn, I., C. Leyval and J. Berthelin, 1993. Cd-tolerant Arbuscular Mycorrhizal (AM) fungi from heavy-metal polluted soils. Plant Soil, 157: 247-256.
Direct Link - Weissenhorn, I. and C. Leyval, 1995. Root colonization of maize by a Cd-sensitive and a Cd-tolerant Glomus mosseae and cadmium uptake in sand culture. Plant Soil 175: 233-238.
Direct Link - Weissenhorn, I. and C. Leyval, 1996. Spore germination of arbuscular-mycorrhizal (AM) fungi in soils differing in heavy metal content and other physicochemical properties. Eur. J. Soil Biol., 32: 165-172.
Direct Link - Wilson, S.C. and K.C. Jones, 1993. Bioremediation of soil contaminated with Polynuclear Aromatic Hydrocarbons (PAHs): A review. Environ. Pollut., 81: 229-249.
CrossRefPubMedDirect Link - Zhu, Y.G., P. Christie and A.S. Laidlaw, 2001. Uptake of Zn by arbuscular mycorrhizal white clover from Zn-contaminated soil. Chemosphere, 42: 193-199.
Direct Link