
Sarika Agarwal
Division of Plant Physiology, Indian Agricultural Research Institute, New Delhi-110 012, India
R.K. Sairam
Division of Plant Physiology, Indian Agricultural Research Institute, New Delhi-110 012, India
R.C. Meena
Division of Biochemistry, Indian Agricultural Research Institute, New Delhi-110 012, India
Aruna Tyagi
Division of Biochemistry, Indian Agricultural Research Institute, New Delhi-110 012, India
G.C. Srivastava
Division of Plant Physiology, Indian Agricultural Research Institute, New Delhi-110 012, India
Journal of Plant Sciences
Year: 2006 | Volume: 1 | Issue: 1 | Page No.: 86-97







ABSTRACT
Effect of excess and deficient levels of iron and copper were studied in wheat genotypes C 306 and Hira grown in Hoagland solution in green house. Iron deficiency resulted in loss of leaf area and total biomass. Both the genotypes showed similar pattern. In case of excess iron treatments there was slight increase in leaf area in both the genotype, whereas decrease was observed in total biomass in comparison to control plants. Copper deficiency caused loss of leaf area and total biomass in both the genotypes. Excess copper treatment resulted in slight increase in leaf area in C 306 than control, while Hira showed slightly lower value than control. Hydrogen peroxide and TBARS contents (lipid peroxidation) increased both under iron and copper deficiency as well as excess condition at both the stages but the increase was more than double in excess iron/copper plants in both the genotypes and at both the stages. Total SOD activity decreased in copper and iron deficient plants and slightly increased in excess copper and iron plants over deficient levels, though the level of activity was less than control plants. Excess copper and iron levels have positive effect on Cu/Zn-SOD and Fe-SOD activities at 30 DAS, while at 45 DAS the activities were more than deficient nutrient levels, but less than control plants. Mn-SOD activity decreased under excess and deficient levels of copper, however, in excess copper plants activity was slightly higher than copper deficient plants. Excess iron has more inhibitory effect on Mn-SOD activity than deficient levels in both genotypes. Iron and copper deficiency decreased APOX and CAT activities below control levels. In case of APOX activity excess copper resulted in greater response at 30 DAS, while excess iron showed more response at 45 DAS. Excess levels of copper and iron increased CAT activity over control at both stages in 2 genotypes, but the response was greater in case of iron.
PDF Abstract XML References
How to cite this article
Sarika Agarwal, R.K. Sairam, R.C. Meena, Aruna Tyagi and G.C. Srivastava, 2006. Effect of Excess and Deficient Levels of Iron and Copper on Oxidative Stress and Antioxidant Enzymes Activity in Wheat. Journal of Plant Sciences, 1: 86-97.
DOI: 10.3923/jps.2006.86.97
URL: https://scialert.net/abstract/?doi=jps.2006.86.97
DOI: 10.3923/jps.2006.86.97
URL: https://scialert.net/abstract/?doi=jps.2006.86.97
INTRODUCTION
Oxidative stress due to excessive production of reactive oxygen species (ROS) viz., superoxide radical (O2.−), hydrogen peroxide (H2O2) and hydroxyl radical (OH.) is a common consequence of most abiotic and biotic stresses (Polle and Rennenberg, 1983). The toxicity of ROS depends on the presence of a Fenton catalyst such as iron or copper ions, giving rise to extremely reactive OH.-radical in the presence of H2O2 and O2.− (Halliwell and Guteridge, 1989; Chen and Schopfer, 1999). These reactive toxic oxygen species causes damage to DNA, proteins, lipids, chlorophyll and almost every other organic constituent of the living cell (Fridovich, 1986; Liebler et al., 1986; Halliwell, 1987; Davies, 1987; Wise and Naylor, 1987; Imlay and Linn, 1988; Becana et al., 1998). Plants protect cell and subcellular systems from the cytotoxic effects of these reactive oxygen species with antioxidant enzymes such as superoxide dismutase (SOD), ascorbate peroxidase (APOX), glutathione reductase (GR), mono-dehydro ascorbate reductase (MDAR), dehydro ascorbate reductase (DAR), catalase (CAT) and metabolites such as glutathione, ascorbic acid, α-tocopherol and carotenoids (Liebler et al., 1986; Elstner, 1986; Bowler et al., 1992; Moran et al., 1994; Menconi et al., 1995; Alscher et al., 1997).
Micronutrients i.e., iron, copper, zinc and manganese are integral constituents of antioxidant enzymes such as Fe-SOD, Cu/Zn-SOD and Mn-SOD, APOX, CAT and POX. As the intrinsic constituent or metal cofactor iron is actively involved in cellular detoxification reactions catalyzed by catalase, phenolic dependent POX, APOX and Fe-SOD, which scavenge hydrogen peroxide and superoxide and protects cell from oxidative injury. In iron deficient sunflower plants, decrease in APOX and non-specific POX activities has been reported by Ranieri et al. (2001). Various workers have reported varied responses of copper in relation to antioxidant enzyme. Luna et al. (1994) reported an increase in SOD activity and no response for APOX and CAT, due to copper application in oat leaves. Greco et al. (1990) have also reported increase in Cu/Zn-SOD activity and it’s mRNA under excess copper. Bueno and Piqueras (2002) reported a very significant increase in SOD (Cu/Zn-SOD) and APOX by copper application in tobacco cell cultures. Excess copper induced APOX gene and consequent increase in APOX has also been reported (Retkevicius et al., 2003; Hang et al., 2004). Loss in SOD activity by copper induced Fenton reaction has also been reported by various workers (Kurepa et al., 1997; Cassano et al., 1997), while an increase in GR activity has also been reported (Kurepa et al., 1997). The excess levels of iron and copper results in oxidative injury in many plant (De Vos et al., 1992; Mazhoudi et al., 1997; Yamamoto et al., 1997). Excess iron and copper induced reactive species have also been reported (Caro and Puntarulo, 1996; Kurepa et al., 1997).
The present study was therefore, planned to study the effect of iron and copper stress (deficiency and excess) on the regulation of antioxidant enzymes activity and oxidative stress in tolerant and susceptible wheat genotypes.
MATERIALS AND METHODS
Plant Material and General Growing Conditions
An experiment was conducted with two genotypes of wheat, Hira (susceptible to water and heat stress) and C 306 (tolerant to water and heat stress) in the Division of Plant Physiology, Indian Agricultural Research Institute, New Delhi, India. Plants were raised in the National Phytotronic facility of the Institute. The temperature, humidity and light/dark periods were maintained at 25EC, 80 and 16/8 h, respectively. Five types of stock solutions were made viz., complete Hoagland solution and Hoagland solutions with double concentration of iron (+ Fe) and copper (+ Cu) and deficient levels of iron (-Fe) and copper (- Cu). The different iron and copper treatments were maintained as following: Control: complete Hoagland solution (with iron and copper at the rate of 4996 and 127 μg L-1, respectively); (-)Fe (without iron); (+) Fe (9992 μg L-1), (-)Cu (without copper), (+)Cu (254 μg L-1). The plants were grown in acid washed sand medium in pots having four plants each. For three days pots were supplied with only double distilled water till the endosperm was exhausted. After that plants were irrigated with complete Hoagland solution or Hoagland solutions with excess or deficient levels of iron/copper and double distilled water given alternatively. First sampling was done on 30th day after germination and second on 45th day after germination. Observation were recorded on total dry matter and leaf area, activities of superoxide dismutase (SOD), ascorbate peroxidase (APOX), catalase (CAT), hydrogen peroxide (H2O2) content and lipid peroxidation in terms of thiobarbituric acid reactive substances (TBARS) contents.
Assays and Estimations
Hydrogen peroxide was estimated by forming titanium-hydro peroxide complex Rao et al., 1997). One gram leaf material was grinded with liquid nitrogen and the fine powdered material was mixed with 10 mL cooled acetone in a cold room (10°C). Mixture was filtered with Whatman No. 1 filter paper followed by the addition of 4 mL titanium reagent and 5 mL ammonium solution to precipitate the titanium-hydro peroxide complex. Reaction mixture was centrifuged at 10000 g for 10 min in the centrifuge (J2-21, Beckman, Geneva, Switzerland). Precipitate was dissolved in 10 mL 2 M H2SO4 and than recentrifuged. Supernatant was read at 415 nm against reagent blank in UV-visible spectrophotometer (model M 36, Beckman, Ca., USA). Hydrogen peroxide contents were calculated by comparing with a standard curve drawn with known hydrogen peroxide concentrations. The level of lipid peroxidation was measured in terms of TBARS content (Heath and Packer, 1968). Leaf sample (0.5 g) was homogenized in 10 mL 0.1% trichloro-acetic acid (TCA). The homogenate was centrifuged at 15000 g for 15 min. To 1.0 mL aliquot of the supernatant 4.0 mL of 0.5% thiobarbituric acid (TBA) in 20% TCA was added. The mixture was heated at 95°C for 30 min in the laboratory electric oven (Scientific, India) and than cooled in an ice bath. After centrifugation at 10000 g for 10 min the absorbance of the supernatant was recorded at 532 nm. The TBARS content was calculated according to its extinction coefficient 0 = 155 mM-1 cm-1. The values for non-specific absorbance at 600 nm were subtracted.
Enzyme extract for superoxide dismutase, ascorbte peroxidase, glutathione reductase and catalase was prepared by first freezing the weighed amount of leaf samples (1 g) in liquid nitrogen to prevent proteolytic activity followed by grinding with 10 mL extraction buffer (0.1 M phosphate buffer, pH 7.5, containing 0.5 mM EDTA and 1 mM ascorbic acid). Brie was passed through 4 layers of cheesecloth and filtrate was centrifuged for 20 min at 15000 g and the supernatant was used as enzyme.
Superoxide dismutase (SOD) was estimated by recording the decrease in optical density of formazone made by superoxide radical and nitro-blue tetrazolium dye by the enzyme (Dhindsa et al., 1981). Three ml of the reaction mixture contained, 13.33 mM methionine (0.2 mL of 200mM), 75 μM nitro blue tetrazolium chloride (NBT) (0.1 mL of 2.25 mM), 0.1 mM EDTA (0.1 mL of 3mM), 50 mM phosphate buffer (pH 7.8) (1.5 mL of 100 mM), 50 mM sodium carbonate (0.1 mL of 1.5M), 0.05 to 0.1 mL enzyme, 0.9 to 0.95 mL of water. Reaction was started by adding 2 μM riboflavin (0.1 mL) and placing the tubes under two 15 W fluorescent lamps for 15 min. A complete reaction mixture with out enzyme, which gave the maximal colour, served as control. Switching off the light and putting the tubes into dark stopped the reaction. A non-irradiated complete reaction mixture served as a blank. Complete reaction mixture plus KCN 3 mM (0.1 mL of 90 mM solution) was used to inhibit Cu/Zn-SOD and complete reaction mixture plus 3 mM KCN (0.1 mL of 90 mM solution) and 5 mM H2O2 (0.1 mL of 150 mM solution) were used to inhibit both Cu/Zn-SOD and Fe-SOD activities. Separate controls (lacking enzymes) were used for total SOD and inhibitor studies. The absorbance was recorded at 560 nm and one unit of enzyme activity was taken as that amount of enzyme, which reduced the absorbance reading to 50% in comparison with tubes lacking enzyme.
Ascorbate peroxidase (APOX) was assayed by recording the decrease in optical density due to ascorbic acid at 290 nm (Nakano and Asada, 1981). The 3 mL reaction mixture contained: 50 mM potassium phosphate buffer (pH 7.0) (1.5 mL of 100 mM), 0.5 mM ascorbic acid (0.5 mL of 3.0 mM), 0.1 mM EDTA (0.1 mL of 3.0 mM), 0.1 mM H2O2 (0.1mL of 3.0 mM), 0.1 mL enzyme. The reaction was started with the addition of 0.1 mL of hydrogen peroxide. Decrease in absorbance for a period of 30 sec. was measured at 290 nm in an UV-visible spectrophotometer (model M 36, Beckman, Ca., USA). Activity is expressed by calculating the decrease in ascorbic acid content by comparing with a standard curve drawn with known concentrations of ascorbic acid.
Catalase (CAT) was assayed by monitoring the decrease in absorbance due to hydrogen peroxide at 240 nm as described by Aebi (1984). The 3.0 mL reaction mixture consisted of potassium phosphate buffer 50 mM (1.5 mL of 100mM buffer, pH 7.0), hydrogen peroxide 12.5 mM (0.5 mL of 75 mM H2O2), enzyme 50 μL and water to make up the volume to 3.0 mL. Adding H2O2 started reaction and decrease in absorbance was recorded for 1 min. Enzyme activity was computed by calculating the amount of H2O2 decomposed by referring to a standard curve of known concentrations of hydrogen peroxide. Soluble protein was estimated according to the method of Bradford (1970). For leaf area and biomass estimation plants were harvested from 4 pots separately for each treatment at 45 DAS. Leaves were separated and their area was estimated with the help of leaf area meter (LiCOR-1600, USA). For biomass estimation samples were dried at 65°C in a hot air oven till constant weights were achieved.
Results
Copper deficiency resulted in leaf tip burning, while excess copper resulted in increased leaf yellowing and enhanced senescence of lower leaves. Copper deficiency caused very significant reduction in total biomass in both the genotypes, while the loss in leaf area though marginal was significant. Under excess copper treatment, leaf area of C 306 was slightly higher than control, while Hira did not show significant variation compared to control. Excess and deficient levels of iron also showed tip burning necrotic patches and leaf yellowing compared to control. Iron deficiency resulted in loss of leaf area and total biomass (Fig. 1A and B) and both the genotypes showed similar pattern. In case of excess iron treatments slight increase in leaf area was observed in C 306, while total biomass decreased in comparison to control. However, even in case of Hira leaf area and biomass were higher than iron deficient plants (Fig. 1A and B). Data also showed that wheat genotype C 306 was generally superior to Hira.
Under excess and deficient copper levels hydrogen peroxide content increased in C 306 and Hira genotype at 30 and 45 DAS. Hydrogen peroxide content also increased both under iron deficiency as well as excess iron condition at both the stages but the increase was more than double in excess iron plants at 45 DAS (Fig. 2A and B). Lipid peroxidation measured as TBARS content increased under deficiency and excess levels of iron and copper in both the genotypes at the two stages. However, the increase was more under excess levels of iron, copper and excess copper was more toxic than excess iron in both the genotypes at both the stages (Fig. 2C and D). Results also showed lower H2O2 and TBARS contents in C 306 than Hira.
Total SOD activity increased in control plants with age from 30 to 45 DAS. However, excess and deficient levels of iron and copper resulted in decrease in activity with age as well as over respective controls at 45 DAS (Fig. 3A and B).
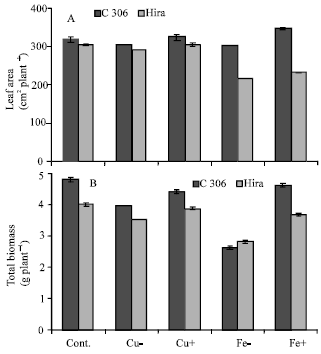 | |
| Fig. 1: | Effect of deficient and excess levels of iron and copper on (A) leaf area and (B) total biomass in two wheat genotypes. Vertical bars indicate±SE of mean. LSD significant at p = 0.05. (Nutrient contents in mg L-1 in different treatments are: Cont. = Cu 127, Fe 4980; Cu - = no Cu; Cu + = 254; Fe- = no Fe; Fe+ = 9960) |
Compared to iron/copper deficient plants, excess iron and copper treatments showed increase in SOD activity in both genotypes at the 2 stages, while in case of C 306 excess copper resulted in increase in activity over control plants at 30 DAS. Cu/Zn-SOD decreased under copper and iron deficiency at the both stages (Fig. 3C and D). At 30 DAS excess copper resulted in increase in activity in C 306 only, while excess iron caused increase in both genotypes. However, at 45 DAS Cu/Zn-SOD activities in excess copper or iron plants was lower than control plants but better than iron/copper deficient plants. Both excess and deficient levels of copper resulted in decrease in Mn-SOD activity, though excess level showed slightly higher activity than copper deficient plant at the 2 stages. Excess and deficient levels of iron also caused decrease in Mn-SOD activity at both stage in the 2 genotypes, however, iron deficient plants showed higher Mn-SOD activity than excess iron plants (Figure 3E and F). Fe-SOD activity, though very low, increased under excess levels of iron and copper and decreased under deficient levels of both metals at both the stages (Fig. 3G and H).
C 306 showed higher overall activity for total and different isoforms of SOD than Hira
Ascorbate peroxidase activity (APOX) increased with age (30 to 45 DAS) under all treatments. APOX activity decreased in deficient levels of iron at 30 DAS, although the activity in excess iron plants was higher than control plants. At 45 DAS very significant increase in APOX activity was observed in excess iron plants over control. Copper deficiency resulted in decrease in APOX activity at the 2 stages, while excess copper caused very significant increase in activity at 30 DAS and marginal increase at 45 DAS compared to control plants in both genotypes (Fig. 4A and B).
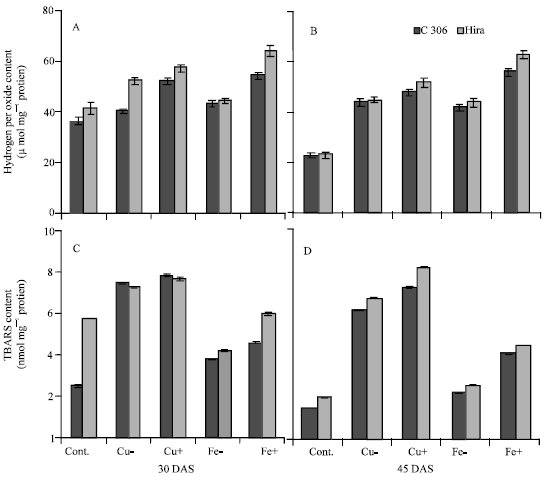 | |
| Fig. 2: | Effect of deficient and excess levels of iron and copper on (A, B) hydrogen peroxide content and (C, D) lipid peroxidation (TBARS content) in two wheat genotypes. Vertical bars indicate±SE of mean. LSD significant at p = 0.05. (Nutrient contents in mg L-1 in different treatments are: Cont. = Cu 127, Fe 4980; Cu- = no Cu; Cu + = 254; Fe- = no Fe; Fe+ = 9960) |
Catalase activity decreased at 45 DAS compared to 30 DAS in all the treatments. Excess copper and iron resulted in increase in activity over control plants, though increase was significantly more in excess iron treated plants than excess copper. Both iron and copper deficiencies resulted in decline in CAT activity; however the inhibitory effect of deficiency was more in case of iron than copper (Fig. 4C and D).
DISCUSSION
Iron and copper are important micronutrients, as these are components of various proteins and enzymes and thereby regulate plant metabolism and growth. Both these metals are also constituents of antioxidant enzymes like Fe-SOD, APOX, CAT and Cu/Zn-SOD and thus have a role in alleviation of oxidative stress. However, reduced forms of both metals favour Fenton reaction (Haber and Weiss, 1934) and thus facilitate generation of highly cytotoxic hydroxyl radical.
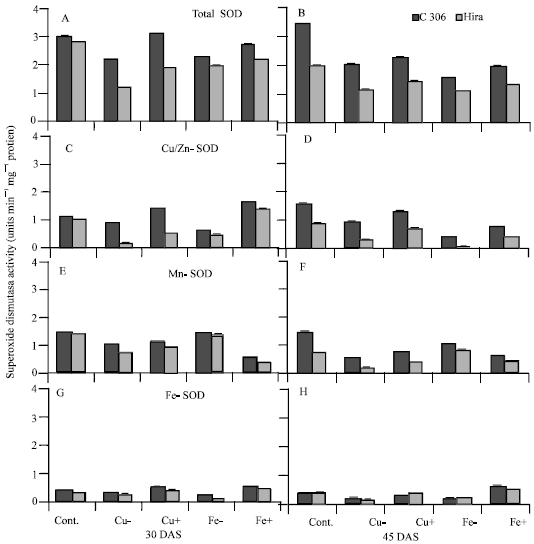 | |
| Fig. 3: | Effect of deficient and excess levels of iron and copper on (A, B) total superoxide dismutase, (C, D) Cu/Zn-SOD, (E, F) Mn-SOD and (G, H) Fe-SOD in two wheat genotypes. Vertical bars indicate"SE of mean. LSD significant at p = 0.05. (Nutrient contents in mg L-1 in different treatments are: Cont. = Cu 127, Fe 4980; Cu- = no Cu; Cu+ = 254; Fe- = no Fe; Fe+ = 9960) |
Hydroxyl radical cause’s lipid peroxidation, pigment bleaching and DNA and protein degradation and thus is the main cause of early senescence, necrotic patches, leaf yellowing and growth retardation. Our study showed that iron and copper deficiency resulted in growth retardation, while excess levels improved growth over deficient levels. Depression in growth parameters under iron deficient conditions and improvement upon iron application have been reported by various workers (Clark and Reinhard, 1991; Hansen et al., 1996; Luo et al., 1997). Copper deficiency also showed decrease in leaf area and total biomass, while excess copper improved both parameters over deficient levels in both genotypes. Growth retardation due to copper deficiency as well as beneficial effect of copper application has also been reported by various workers (Luna et al., 1994; Guster, 1990; Owuoche et al., 1995; Seliga, 1999).
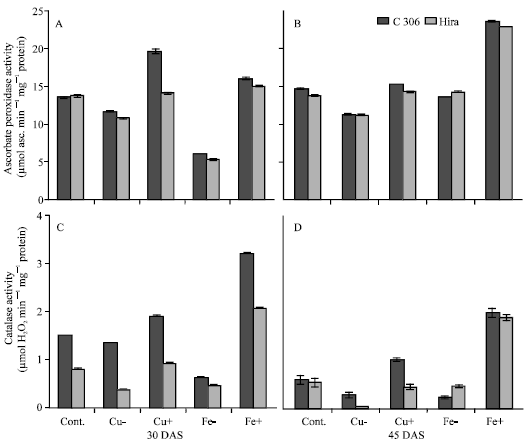 | |
| Fig. 4: | Effect of deficient and excess levels of iron and copper on (A, B) ascorbate peroxidase and (C, D) catalase activity in two wheat genotypes. Vertical bars indicate±SE of mean. LSD significant at p = 0.05. (Nutrient contents in mg L-1 in different treatments are: Cont. = Cu 127, Fe 4980; Cu- = no Cu; Cu+ = 254; Fe- = no Fe; Fe+ = 9960) |
Iron deficiency resulted in decline in activity of SOD and its isozymes, ascorbate peroxidase and catalase. Similar results of inhibition of Fe-SOD activity (Luna et al., 1994), ascorbate peroxidase and catalase (Iturbe-Ormaetxe et al., 1995) have also been reported earlier. It seems iron not only has a direct role as prosthetic group in case of Fe-SOD but also has some regulatory role on other SOD isoforms as its deficiency also inhibited Cu/Zn-SOD activity and a continuous deficiency as observed in 45 days old plants also has negative effect on Mn-SOD as well, resulting in decline in total SOD activity. The response in case of APOX and CAT again could be due to its role in being constituents of these enzymes. Fang et al. (2001) reported a significant increase in APOX and GR activity by iron in rice leaves. A very significant increase in Fe-SOD, APOX and CAT by 100 mM iron in tobacco has been reported by Bueno and Piqueras (2002). Fourcroy et al. (2004) have reported iron regulated expression of cytosolic APOX. However, excess iron also increase Fenton reaction and may cause oxidative loss of defence enzymes (Cassano et al., 1997; Fang et al., 2001), which may be the reason of decrease in Cu/Zn-SOD, Mn-SOD and total SOD activity at 45 DAS, though ascorbate peroxidase and catalase showed very significant increase in activity due to excess iron.
Studies conducted with excess and deficient copper levels revealed different responses for various antioxidant enzymes as well as two wheat cultivars. Copper deficiency in general resulted in decline in SOD activity, its various isozymes, APOX and CAT. A decrease in antioxidant enzyme activity in tobacco plants due to copper deficiency has been reported by Yu et al. (1998). Copper is a component of a major SOD isoform, i.e., Cu/Zn-SOD and its deficiency, therefore, will have direct bearing on total SOD activity. However, a decline in Mn-SOD and Fe-SOD activity in both wheat cultivars showed that copper might be having some regulatory role on the synthesis/activation of other SOD isoforms, other than its role of being metal prosthetic group. Similar conclusions can also be drawn with relation to reduction in APOX and CAT activity under copper deficiency, which accentuated with age.
Excess copper resulted in decrease in total SOD, particularly Mn-SOD at 30 and 45 days in both wheat cultivars, while prolonged treatment (45 DAS) also caused inhibition of Cu/Zn-SOD. These suggest that oxidative damage by cytotoxic levels of copper is experienced both by Mn-SOD and Cu/Zn-SOD, resulting in total-SOD activity lower than control plants. However, a very significant increase was observed in case of APOX and CAT activities in both the cultivars. Various workers have reported varied responses of copper in relation to antioxidant enzyme. Luna et al. (1994) reported an increase in SOD activity and no response for APOX and CAT, due to copper application in oat leaves. Greco et al. (1990) have also reported increase in Cu/Zn-SOD activity and it’s mRNA under excess copper. Similar results were also reported by other workers (Carri et al., 1991 and Chongpraditnum et al., 1992). Bueno and Piqueras (2002) reported a very significant increase in SOD (Cu/Zn-SOD) and APOX by copper application in tobacco cell cultures. Excess copper induced increase in APOX activity due to gene induction has also been reported (Sauser et al., 1997; Retkevicius et al., 2003; Hang et al., 2004). Loss in SOD activity by copper induced Fenton reaction has been reported by various workers (Kurepa et al., 1997; Cassano et al., 1997), while an increase in GR activity has also been reported (Kurepa et al., 1997). The decrease in CAT activity in copper deficient plants in both varieties and improvement in activity over copper deficient plants as well as increase over control plants suggest that copper has regulatory role on CAT either by way of activation or gene induction, as it is not a constituent of CAT enzyme.
Both excess and deficiency of iron resulted in increase in oxidative stress as is clear from increase in hydrogen peroxide (H2O2) content and lipid peroxidation (TBARS content). While increased oxidative stress under complete iron deficiency could be due to loss in ROS scavenging activity due to decrease in APOX, CAT and SOD. A similar effect under excess iron is probably due to an enhanced Fenton activity by excess iron. Increase in oxidative injury by excess concentration of redox active metals such as iron and copper have also been reported (Yamamoto et al., 1997; Ranieri et al., 2001).
Similar to the effect of iron; copper deficiency and excess application resulted in increased oxidative stress (H2O2 and TBARS content). Like iron, copper is also a redox active metal and increased oxidative stress under excess copper could be due to increase in Fenton activity, as also reported by various other workers (De Vos et al., 1992; Mazhoudi et al., 1997). The increase in oxidative stress under copper deficiency is most probably due to decrease in antioxidant activity as evidenced by lower levels of SOD, APOX and CAT. It is evident that inhibitory effect of excess copper induced oxidative stress was more than compensated by increase in antioxidant activity as observed by increase in SOD, APOX and CAT resulting in to vegetative growth at par with control plants.
From the aforesaid discussion it is evident that deficiency of iron and copper result in heightened oxidative stress, which was due to decline in antioxidant enzyme activity and was manifested in reduced plant growth. The excess metal though caused some increase in antioxidant enzymes activity over the deficient metal levels, but the increase in oxidative stress was more than the deficient levels, due to Fe/Cu ion induced Fenton reaction. The over all influence of slight increase in antioxidant activity and higher oxidative stress was reflected in growth, which was better than deficient metal levels.
The effects of iron or copper on antioxidant enzymes could be due to these being components of respective prosthetic groups of the enzymes such as Fe-SOD, APOX and CAT in case of iron and Cu/Zn-SOD in case of copper. However, the positive effects of iron on Cu/Zn-SOD and of copper on APOX and CAT suggest that these metal ions may have some regulatory role either on the activation of these antioxidant enzymes or at the level of gene induction or its translation. However, these aspects require further elaboration. The beneficial effects of control levels of iron and copper over complete deficiency in terms of antioxidant enzymes activity and growth was generally more in tolerant genotype C 306, which also experienced less oxidative stress than Hira. It seems that an inbuilt efficient mechanism exists in tolerant cultivar, which responds to the availability of required metal ions by increasing antioxidant enzyme activity and consequently decrease in oxidative stress.
REFERENCES
- Aebi, H., 1984. Catalase in vitro. In: Methods in Enzymology, Packer, L., Academic Press, Cambridge, Massachusetts, United States, ISBN: 9780121820053, pp: 121-126.
CrossRefDirect Link - Alscher, R.G., J.L. Donahue and C.L. Cramer, 1997. Reactive oxygen species and antioxidants: Relationship in green cells. Physiol. Plant., 100: 224-233.
CrossRefDirect Link - Bowler, C., M.V. Montagu and D. Inze, 1992. Superoxide dismutase and stress tolerance. Annu. Rev. Plant Physiol. Mol. Biol., 43: 83-116.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Caro, A. and S. Puntarulo, 1996. Effect of in vivo iron supplementation on oxygen redical production by soybean roots. Biochim. Biophys. Acta, 1291: 245-251.
PubMed - Chen, S.X. and P. Schopfer, 1999. Hydroxyl-radical production in physiological reactions: A novels function of peroxidase. Eur. J. Biochem., 260: 726-735.
CrossRefPubMedDirect Link - Chongpraditrum, P., S. Mori and M. Chino, 1992. Excess copper induces a cytosolic Cu, Zn-superoxide dismutase in soybean root. Plant Cell Physiol., 33: 239-244.
Direct Link - Davies, K.J.A., 1987. Protein damage and degradation by oxygen radicals. I. general aspects. J. Biol. Chem., 262: 9895-9901.
PubMedDirect Link - De Vos, C.H.R., M.J. Vonk, R. Vooijs and H. Schat, 1992. Glutathione depletion due to copper induced phytocheation synthesis causes oxidative stress in Silene cucubalus. Plant Physoil., 98: 853-858.
Direct Link - Fridovich, I., 1986. Biological effects of superoxide radical. Arch. Biochem. Biophys., 247: 1-11.
CrossRef - Haber, F. and J.J. Weiss, 1934. The catalytic decomposition of hydrogen peroxide by iron salts. Proc. R. Soc. Lond., 147: 332-351.
Direct Link - Halliwell, B. and J.M. Gutteridge, 1984. Oxygen toxicity, oxygen radicals, transition metals and disease. Biochem. J., 219: 1-14.
PubMed - Halliwell, B., 1987. Oxidative damage, lipid peroxidation and antioxidant protection in chloroplasts. Chem. Phys. Lipids, 44: 327-340.
CrossRefDirect Link - Heath, R.L. and L. Packer, 1968. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys., 125: 189-198.
CrossRefPubMedDirect Link - Imlay, J.A. and S. Linn, 1988. DNA damage and oxygen radical toxicity. Science, 240: 1302-1309.
CrossRefDirect Link - Iturbe-Ormaetxe, I., J.F. Moran, C. Arrese-Igor, Y. Gogorcena, R.V. Klucas and M. Becana, 1995. Activated oxygen and antioxidant defences in iron-deficient pea plants. Plant Cell Environ., 18: 421-429.
Direct Link - Luna, C.M., C.A. Gonzalez and V.S. Trippi, 1994. Oxidative damage caused by an excess of copper in oat leaves. Plant Cell Physiol., 35: 11-15.
Direct Link - Luo, A., G. Jing, P. Wu, J. Ni, S. Jiang and Y. Zhang, 1997. Rice genotypes differences in nutrient status under excessive ferric iron conditions. J. Plant Nutr., 20: 1361-1373.
CrossRef - Mazhoudi, S., A. Chaoui, M.H. Ghorbal and E.E. Ferjani, 1997. Response of antioxidant enzymes to excess copper in tomato (Lycopersicon esculentum, Mill). Plant Sci., 127: 129-137.
CrossRefDirect Link - Menconi, M., C.L.M. Sgherri, C. Pinzino and F. Navari-Izzo, 1995. Activated oxygen production and detoxification in wheat plants subjected to a water deficit programme. J. Exp. Bot., 46: 1123-1130.
CrossRef - Nakano, Y. and K. Asada, 1981. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in Spinach chloroplasts. Plant Cell Physiol., 22: 867-880.
CrossRefDirect Link - Sauser, K.R., J.K. Liu and T.Y. Wong, 1997. Identification of a copper sensitive ascorbate peroxidase in the unicellular green alga Selenastrum capricornutum. Biometals, 10: 163-168.
PubMed - Moran, J.F., M. Becana, I. Iturbe-Ormaetxe, S. Frechilla, R.V. Klucas and P. Aparicio-Tejo, 1994. Drought induces oxidative stress in pea plants. Planta, 194: 346-352.
CrossRef
sandeep sharma Reply
Sir I am a PhD student in IARI New Delhi India, Please send me this paper.