
S. Johri
Division of Biotechnology and Division of Pharmacology, Regional Research Laboratory, Canal Road, Jammu-Tawi, 180 001, J and K, India
U. Jamwal
Division of Biotechnology and Division of Pharmacology, Regional Research Laboratory, Canal Road, Jammu-Tawi, 180 001, J and K, India
P. Kaiser
Division of Biotechnology and Division of Pharmacology, Regional Research Laboratory, Canal Road, Jammu-Tawi, 180 001, J and K, India
S. A. Tasduq
Division of Biotechnology and Division of Pharmacology, Regional Research Laboratory, Canal Road, Jammu-Tawi, 180 001, J and K, India
S. Rasool
Division of Biotechnology and Division of Pharmacology, Regional Research Laboratory, Canal Road, Jammu-Tawi, 180 001, J and K, India
V. Verma
Division of Biotechnology and Division of Pharmacology, Regional Research Laboratory, Canal Road, Jammu-Tawi, 180 001, J and K, India
G. N. Qazi
Division of Biotechnology and Division of Pharmacology, Regional Research Laboratory, Canal Road, Jammu-Tawi, 180 001, J and K, India
Journal of Plant Sciences
Year: 2006 | Volume: 1 | Issue: 1 | Page No.: 72-79







ABSTRACT
This study reports the isolation of a robust class 111 peroxidase (EC.1.11.1.7) from roots of Withania somnifera (WSPOD), which has the ability to oxidize indole-3-acetic acid (IAA) to free radicals like indolyl, skatolyl and peroxyl radicals which are highly cytotoxic species. WSPOD is not toxic when incubated independently in the assay system at equivalent concentrations. WSPOD, a monomer glycoprotein of 34 kDa and pI 5.2 is a highly thermotolerant enzyme with optimum pH ~6.0 and stable in the pH range of 3-9. Km and Vmax for IAA was calculated to be 0.17 mmol and 0.66 μmol min-1, respectively. The enzyme did not require H2O2 for the activity but needed 2,4-DCP and manganese ions as cofactors.
PDF Abstract XML References
How to cite this article
S. Johri, U. Jamwal, P. Kaiser, S. A. Tasduq, S. Rasool, V. Verma and G. N. Qazi, 2006. A Novel Peroxidase from Withania somnifera. Journal of Plant Sciences, 1: 72-79.
DOI: 10.3923/jps.2006.72.79
URL: https://scialert.net/abstract/?doi=jps.2006.72.79
DOI: 10.3923/jps.2006.72.79
URL: https://scialert.net/abstract/?doi=jps.2006.72.79
INTRODUCTION
Indole-3-acetic acid (IAA), a plant growth hormone and some of its derivatives, upon oxidation by Horseradish Peroxidase (HRP) produce lipid peroxidation in lipid vesicles, damage to nucleic acids and cytotoxicity (Candeias et al., 1995, 1996; Pires de Melo et al., 1997; Folkes et al., 1999). Recent experimental evidence has suggested that IAA/HRP can be delivered systemically to be activated only in the tumour and thus could be a basis for the targeted cancer therapy (Folkes et al., 1998; Greco et al., 2000; Folkes and Wardman, 2001; Wardman, 2002). Horseradish Peroxidase (HRP) is generally preferred for carrying out oxidation of IAA, because of its having better substrate specificity, thermostability, no requirement for cofactors, yield and economic feasibility. In the present communication we report application of another highly robust heme peroxidase in the oxidative activation of IAA isolated from the roots of Withania somnifera. The purified enzyme when used in combination with IAA exhibited almost similar activity towards lipid peroxidation in rat microsomes, damage to the DNA and cytotoxicity as reported for HRP.
Materials and Methods
Germ plasm of different accessions of W. somnifera collected from the different parts of India is being maintained in our institute. Authenticated samples of the roots of W. somnifera were collected from 3-month old plants. FPLC apparatus and the columns used were obtained from M/S Amersham Biosciences. Ready Strip was obtained from BioRad. All other chemicals were of high purity reagent grade.
Isolation of Peroxidase from Withania somnifera Roots
Small pieces of roots (35 g fresh weight) from 3-month old plants were pulverized and homogenized in a Tris-HCl buffer (20 mM, pH 7.5). The homogenate was allowed to pass through four layers of cheesecloth and then through 0.45 μM filter (Millipore). The cell free extract (CFE) was freeze dried and stored at-20°C. Freeze dried powder was reconstituted in 50 mL of 5 mM Tris-HCl buffer (pH 7.5) and centrifuged at 15,000xg for 15 min at 4°C. The supernatant was filtered through ultra membrane (Amicon) with ~10 kDa cut off. The retentate was collected and applied on a DEAE-Sepharose column (2x10 cm) equilibrated in 5 mM Tris-HCl buffer (pH 7.5). The column was washed with 5 bed volumes of equilibration buffer and the proteins eluted with a linear gradient of 0-0.5 M NaCl in equilibration buffer. Fractions of 2 mL were collected and peroxidase activity was determined in each fraction in a 96-well microtiter plate assay performed in presence of 80 mM guaiacol, 100 mM sodium acetate buffer (pH 5.5) and 80 mM hydrogen peroxide. Appearance of dark reddish brown color indicated the presence of activity in the fractions. Fractions with peroxidase activity were pooled and after the addition of ammonium sulpfate (final concentration ~1.7 M), loaded onto a phenyl-Sepharose column (1.6x15 cm) pre-equilibrated with 20 mM Tris-HCl, buffer (pH 7.5). The column was washed with 5 bed volumes of equilibration buffer and the proteins were eluted with a linear gradient (1.5 mL min-l) of equilibration buffer (20 mM Tris-HCl, pH 7.5). All steps in the purification were performed at 25°C. The active fraction was applied to a Mono Q HR 5/5 (1 mL) column equilibrated with 10 mM Tris-HCl buffer (pH 7.5) and eluted with a gradient of 0-0.5 M NaCl in equilibration buffer at a flow rate of 0.75 mL min-1. Fractions (0.5 mL) were collected and assayed for the protein and heme content. Protein concentration was determined by the method of Bradford (1976) with BSA as the standard. Heme content was monitored in a spectrophotometer at 403 nm. Reinheitzahl (RZ) value was determined as a ratio of OD403/OD280 and taken as an index of heme purity. Zymogram was obtained on 8% native-PAGE with protein sample (5-20 fmol of purified WSPOD) dissolved in loading buffer without SDS and thiol-reducing agents. Proteins were separated at 10 V cm-1.
Characterization of Purified WSPOD
Determination of molecular weight was done by SDS-PAGE by the method of Laemmli (1970) using pre-stained molecular weight markers in the range of 18 kDa-216 kDa; (Pierce). ReadyStrip (IPG strip) of 7 cm with pI range of 3-6 was used for the determination of pI using protein IEF cell. The gel strip was rehydrated with rehydration buffer (available with the kit) along with the sample (0.5 pmole of purified WSPOD) or with 10 μL of pI standards (low range, BIORAD) overnight. The samples were focused for 8000 V-h with rapid ramping at 20°C according to the manufacturer’s instructions (BIORAD). The gels were equilibrated for 30 min in 50 mM sodium acetate buffer, pH 5.5 and stained for peroxidase activity with 80 mM guaiacol and 80 mM H2O2 in fresh sodium acetate buffer. The migration of activity band and standards was measured and the pI determined by interpolation.
For determining the thermal stability, 0.3 μM of the purified peroxidase was dissolved in 5 mM acetate buffer (pH 5.5) and incubated at different temperatures 30-80°C. An aliquot of 0.05 mL was withdrawn at 60 min intervals from each tube, cooled by immersing in ice and assayed for residual peroxidase activity. Optimum pH and the pH stability of purified peroxidase was determined by assaying the activity using the guaiacol as the substrate in the pH range of 2-10 obtained by using glycine-HCl (pH 2-3), acetate (pH 4-5), phosphate (pH 6-7), Tris-HCl (pH 8) and glycine-NaOH (pH 9-10) buffers at 25°C.
Biochemical Assays
The oxidation of IAA was determined by measuring the residual IAA in reaction mixture using Salkowski reagent (Huang and Haard, 1977). The reaction mixture consisted of 1 mM IAA (0.3 mL), 1 mM 2, 4-DCP (0.3 mL), 1 mM MnCl2 (0.3 mL) in 0.2 mL of 0.1 M acetate buffer pH 5.5 to the final volume 1.5 mL. The reaction was started by the addition of enzyme extract (10 μl). After 40-50 min of incubation at 30°C, 2.0 mL of Salkowski reagent (1.5 mL of 500 mM FeCl3, 50 mL of water and 30 mL of concentrated sulfuric acid) were added. Absorbance at 525 nm was determined after the samples were stored for 30 min in the dark. Absorbance was converted to μmol of residual IAA using a standard curve for IAA.
Kinetic measurements were performed in the IAA concentration range of 0.01-2 mM and 0.04 μM WSPOD in 0.1 M acetate buffer pH 5.5. The standard error determined from 3 repetitions was <4%. Km and Vmax values were obtained from double-reciprocal plots and turnover numbers were calculated by dividing Vmax by the enzyme concentration.
In vitro Lipid Peroxidation
In vitro lipid peroxidation was determined in microsomes obtained from rat liver as per the procedure described by Tasaduq et al. (2003). Liver microsomes (5 mg protein mL-1 in 0.15 M NaCl, pH 7) were incubated for 20 min at 37°C in absence (control) and presence of 100 μM FeSO4+50 μM H2O2. In identically set up incubations, IAA with and without WSPOD were added. Reaction was terminated by addition of 2.0 mL TCA (5% w/v) and LPO content determined as nmol MDA formed mg-1 microsomal protein.
MTT Reduction Assay
MTT reduction assay was performed in hepatocytes prepared by collagenase perfusion technique (Minotti and Aust, 1987). Hepatocytes (2x106 cells) were incubated William’s medium E contained in 60 mm per collagen coated plates in a CO2 incubator. Plates were washed after 3 hr with ice cold PBS to remove the unattached cells. Fresh medium having 200 μL mL-1 of MTT (60 μg mL-1) dye was added to the cultures in presence/absence of WSPOD and IAA and incubations were allowed to proceed for a further 2 h. Medium was centrifuged (100xg for 5 min) and the supernatant was collected. The resulting formazan crystals were dissolved in DMSO and absorbance of the resulting formazan read at 565 nm.
Plasmid Band Shift Assay
Peroxidase-induced binding of indoles to DNA was tested by a plasmid band shift assay (Folkes et al., 1999). Plasmid (pUC19) DNA was isolated from Escherichia coli strain JM110 using a Qiagen Kit (Mega plasmid). Plasmid DNA (1 mg mL-1) was incubated with 2 mM IAA with or without WSPOD (10 μg mL-1) for 1 h at 37°C. The samples were then loaded on 1.0% agarose gels for electrophoresis in TBE buffer (89.2 mM Tris base, 89 mM boric acid, 2 mM EDTA, pH 8) for 16 h at 16 V, using mini DNA cell (Bio-Rad) with a model 200/2.0 power supply. The DNA was stained with ethidium bromide (0.5 μg mL-1) and photographed under UV light.
Results and Discussion
Chromatographic separation by FPLC using DEAE-Sepharose, phenyl-Sepharose and Mono-Q columns yielded 0.18 mg protein with a final peroxidase to protein enrichment of ~284 fold (Fig. 1a-d Table 1) and ~55% recovery.
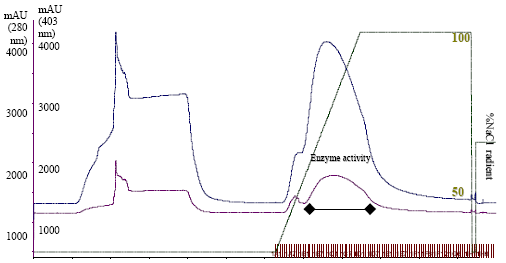 | |
| Fig. 1a: | FPLC of CFE of Withania somnifera roots on DEAE-Sepharose column. |
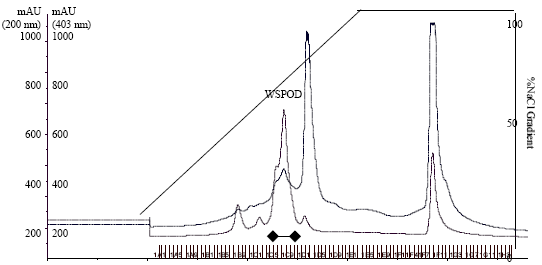 | |
| Fig. 1b: | FPLC of partially purified WSPOD on phenyl-Sepharose column. |
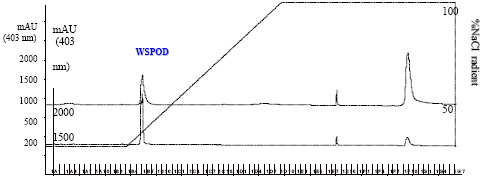 | |
| Fig. 1c: | FPLC of partially purified WSPOD on Mono Q column (flow rate 1 mL min-1) |
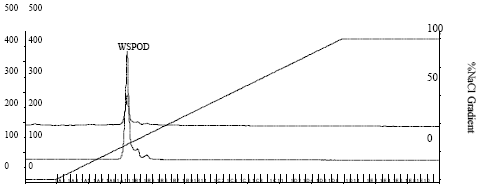 | |
| Fig. 1d: | FPLC of purified active fraction from on Mono Q column on Mono Q column (flow rate 0.5 mL min-1) |
| Table 1: | Purification of root peroxidase WSPOD from Withania somnifera |
 | |
RZ: purity index of the heme content was 3.1 as determined by the absorption ratio of A403 and A280. WSPOD was optimally active at pH 6.0 in sodium acetate buffer with guaiacol as substrate and exhibited stability in the pH range of 3 to 9.
Average molecular mass of purified peroxidase was ~36 kDa as shown by SDS-PAGE (Fig. 2) and has a pI value of 5.2. Treatment with 2-mercaptoethanol did not yield additional bands, indicating that WSPOD is a monomer protein. The purified peroxidase exhibited thermal stability upto 80°C for 1h with the retention of 40-60% of the original activity. After heating at 60°C and separation on SDS gel system, peroxidase maintained its ability to oxidize guaiacol and to develop brown band on the gel (Fig. 2). The thermal stability of WSPOD was comparable with peroxidase reported from palm but greater than those reported from coconut, strawberry and cotton (Deepa and Arumughan, 2002; Mujer et al., 1983; Civello et al., 1995; Triplett and Melton, 1992).
| Table 2: | Effect of WSPOD on IAA oxidation |
 | |
| Values are mean±SE from three independent experiments | |
| Table 3: | Effect of IAA/WSPOD on lipid peroxidation in rat liver microsomes |
 | |
| Values are mean±SE from three independent experiments | |
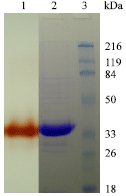 | |
| Fig. 2: | 10% SDS-PAGE of Lane 1: Purified WSPOD (Activity staining); Lane 2: Purified WSPOD (Protein staining); Lane 3: Protein MW markers |
Catalytic properties of the Withania somnifera peroxidase with regard to oxidation of IAA were investigated. The homogenous enzyme was capable of oxidising IAA in the absence of H2O2 (Table 2) but required divalent manganese ion and 2,4-dichlorophenol as cofactors. These results are not in conformity with the reports on the degradation of IAA by HRP and tobacco peroxidase for which the requirement of cofactors was reported to be obligatory (Gazaryan and Lagrimini, 1996).
Kinetic curve of IAA oxidation by purified WSPOD had a characteristic sigmoid shape and followed Michaelis-Menten kinetics in the range of 0.015-0.5 mmol at 0.04 μmol concentration.
The values of Km and Vmax of the purified WSPOD for IAA were 0.17 mmol and 0.66 μmol min-1 with turnover number of 37.5 sec-1 (Fig. 3). Km of purified WSPOD for IAA was observed to be lower than that reported for tobacco peroxidase and comparable with HRP (Gazaryan and Lagrimini, 1996).
Incubation of microsomes with IAA/WSPOD increased the LPO levels in a concentration dependent manner by 0.06 to 4.8 fold (Table 3) and decreased the hepatocyte viability (Table 4). IAA/WSPOD caused single-and double-strand breaks as shown by an increase in circular and linear DNA.
| Table 4: | Cytotoxic effect of IAA/WSPOD in rat hepatocytes |
 | |
| Values are mean±SE from three independent experiments | |
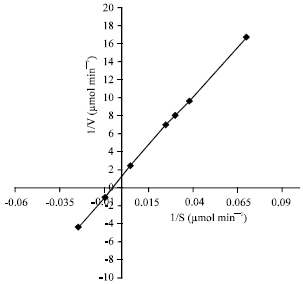 | |
| Fig. 3: | kM of purified WSPOD for IAA |
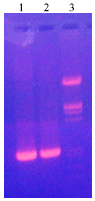 | |
| Fig. 4: | DNA adduct formation Lane 1: pUC19; Lane 2: pUC19+IAA; Lane 3: pUC19+IAA+WSPOD |
Increased lipid peroxidation in microsomes with decreased hepatocyte viability and the formation of DNA adducts is due to the formation of highly reactive indolyl, skatolyl and peroxyl radicals by one-electron oxidation of IAA by WSPOD (Fig. 4). Subsequent products include 3-methylene-2-oxindole, which is highly reactive towards cellular nucleophiles such as thiols and DNA (Wardman, 2002).
In conclusion, present results demonstrate significant cytotoxicity of IAA in combination with a robust heme peroxidase from Withania somifera not requiring added hydrogen peroxide cofactor. These results are in conformity with the cytotoxicity induced by IAA in combination with HRP which is considered a model prodrug/enzyme combination for targeted cancer therapy (Wardman, 2002).
REFERENCES
- Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link