
Ercan Ceyhan
Department of Field Crops, Agriculture Faculty,
Selyuk University, Campus, Konya, Turkey
Journal of Plant Sciences
Year: 2006 | Volume: 1 | Issue: 2 | Page No.: 138-143







ABSTRACT
Four pea lines as female parents (Carina, Sprinter, Cosmos, Manuel) were crossed with three male tester parents (B1, B6 and B12) and 12 F1 hybrids and 12 F2 populations were evaluated in Konya, during 2000-2001 and 2001-2002 growing seasons. General and specific combiners suitable for use in breeding programmes were identified from a linextester cross in peas. Although several genetic studies have been conducted, there is not a general consensus as the made gene action controlling the expression of winter hardiness in pea. The inheritance of cold hardiness was controlled mainly by non- additive gene action. Sprinter and B12 were good general combiners for cold hardiness. The best specific combiners were SprinterxB1 for cold hardiness. The average level of heterosis and heterobeltiosis for winter hardiness in F2 population was low that in F1 hybrids. Broad sense heritability was high due to the major role of environmental factors in expression of winter hardiness in pea. Thus, delay selection was suggested for breeding of winter resistance.
PDF Abstract XML References
How to cite this article
Ercan Ceyhan, 2006. Genetic Analysis of Cold Hardiness in Peas (Pisum sativum L.). Journal of Plant Sciences, 1: 138-143.
DOI: 10.3923/jps.2006.138.143
URL: https://scialert.net/abstract/?doi=jps.2006.138.143
DOI: 10.3923/jps.2006.138.143
URL: https://scialert.net/abstract/?doi=jps.2006.138.143
P>
INTRODUCTION
Pea seeds are rich in protein and most mineral elements. Pea varieties are sown in the spring and their yields are very variable, being very dependent on the temperature and water supply at the end of their growth cycle. Autumn-sown varieties may have higher and more stable yields than spring-sown ones: they also enter the flowering period earlier and so avoid stresses due to the high temperatures and poor water supply which are frequently encountered in late spring. Hovewer, autumn sowing is only possible if the varieties are winter hard (Bourion et al., 2003). Winter hardiness is one of the major a biotic stresses in pea. Winter hardiness is a complex quantitative trait conditioned by the plant genotype and the environment in which is grown. Current varieties grown in World are considered as cold tolerant. Winterhardiness has been sought by North American and French scientists (Auld et al., 1983; Eteve, 1985) and although some level tolerance to cold has been obtained it remains a challenge (Murray and Swensen, 1991). The level of winter hardiness does not match that of the winter cereals.
Most winter pea breeding programs have used winter Pisum fulvum pea because of their good resistance to freezing: such genotypes can survive temperatures of -6°C (no damage to leaves; 100% survival) (Balackova et al., 1986). Although several genetic studies have been conducted, there is not a general consensus on the made of gene action controlling the expression of cold hardiness in pea. Genetic studies on winter hardiness in pea have rivaled that it is quantitative in nature and governed by intermediate dominance (Cousin et al., 1985) and additive genes (Auld et al., 1983). Liesenfeld et al. (1986) reported that as few as three or four genes or linkage groups could be responsible for winter hardiness. They also observed that winter survival is associated with genes which produce pigmented hilum and pigmented seed coat.
Cultivation of pea in Turkey is very limited mainly in Western Anatolia region, compared to beans, chickpea and lentils, which constitute approximately 95% of grain legumes production. This research aims to investigate to inheritance of cold hardiness in pea as a good source of plant protein. If winter tolerant or resistant varieties can be selected and grown successfully, pea can be involved in crop rotation in Turkey.
MATERIALS AND METHODS
Seven pea lines, Carina, Sprinter, Cosmos, Manuel representing the main cultivars used in dried and canning pea breeding and B1, B6 and B12 (obtained by Dr. Ahmet TAMKOÇ) representing a wide range in winter survival ability were chosen as parental material. The four pea cultivars (lines) and tree lines (tester) were made by Line x Tester. The seven parents were established under the field condition during 2000 summer growing season and crossed in 12 combinations including LinexTester. The 12 F1 and F2 populations with the seven parents were planted in a randomized complete blocks design with tree replications at the experimental site of Faculty of Agriculture, Selcuk University, in Konya, during 2000-2001 and 2001-2002 growing seasons. While the plots consist of one rows in F1 populations, F2 populations consist of three rows, each row containing five plants.
Cold hardiness was examined in F1 and F2 populations. All plant material tested for cold hardiness was grown under field conditions. F1 populations were scored according to five categories (1= extremely cold sensitive, with 100% plants killed; 5 = most cold tolerant, with less than 25% plants killed) (Guye et al., 1987), as F2 populations were survival on the each plot. Data were collected on plants in the each plot.
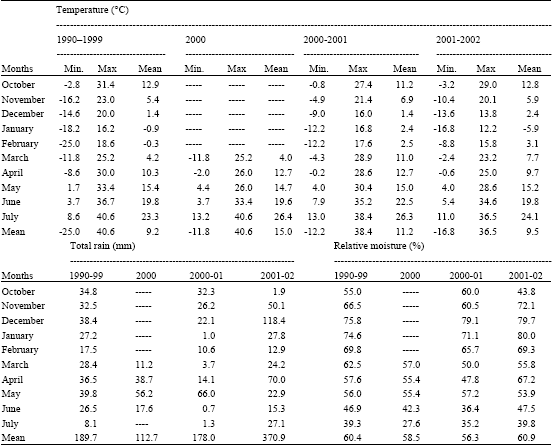
In both experimental years, monthly minimum, maximum and mean temperature values were higher than observed for 10-year mean (Table 1). Similarly, total rainfall values were higher than observed for the 10-year mean in both grown seasons. First grown season was lower than 10-year mean value of 60.4%. However, 10-year was higher than second grown seasons.
Regarding statistical analysis, parents and F2 population data were analyzed together as suggested by Sing and Chaudhary (1979). The combining ability analysis was done following Kempthore (1957). Broad and narrow sense heritabilities were calculated for each character by using the Falconer’s (1982) methods. Heterosis over mid-parent (HMP) or the better parent (HBP) of the observed characters were also calculated.
RESULTS AND DISCUSSION
Growth Habit and Winter Hardiness
Cold hardiness was lower at F2 populations than at F1 hybrids. At a given sub-zero temperature in the ranged 0 to -16.8°C the freezining injury observed in the field. At below -10°C, damage first appears on the edge of the leaflets, later on the whole leaflets, the main stem. The young tissues are more resistant. The B1, B6 and B12 lines had not injury at -16.8°C on January in 2002. The F1 and some F2 populations had more resistant than the female. Frost occurring on January of relatively minimum temperatures destroyed in plants from F2 populations and the female. However, a period of frost in 2000-2001 grown seasons of F1 hybrids had not cause any injury. The air temperatures gradually drop during the winter plants starts to redevelop cold resistance. Very high levels of cold hardiness are obtained by B1, B6 and B12 lines among the parents. The highest level of cold hardiness was obtained by SprinterxB12 crosses among the hybrids. This region, air minimum temperatures ranged from 0 to -25.0°C (Table 1) while short periods at -13.0°C air temperatures have little effect on cold hardiness pea hybrids, long period of -13.0°C cause severe damage.
| Table 1: | Monthly minimum, maximum and mean temperature, relative moisture and total rainfall during the 2000, 2000-2001, 2001-2002 and 10- years average growing seasons |
 | |
| Table 2: | Mean squares related to cold hardiness in line x tester analysis |
 | |
| **: p<0.01 | |
| Table 3: | Estimates of components of genetic variance for cold hardiness in pea |
 | |
| σ2GCA: General Combining ability. σ2SCA: Specific Combining ability. σ2D: Additive variance. σ2H: Dominant Variance. H2: Broad sense heritability. h2: Narrow sense heritability | |
Balackova et al. (1986) obtained that among the Pisum fulvum cultivars, a temperature of -6°C caused no damage to leaves (100% survival). Resistance in the cultivars of Pisum sativum ranged 5 to 100%; it was highest in four conventional lines and five short-stemmed lines. In France, air temperatures -23°C and below are considered lethal to overwintering pea, while air temperatures of -6 to -14°C are not injurious to fully hardened pea (Eteve, 1985). Our some hybrids survived exposure to air temperatures of -16.8 0C without snow cover in field. The findings of this study agree with reported by above literatures.
The analysis of variance of F2 populations indicated that variation among crosses was significant for winter hardiness (Table 2). Statically analysis of the results showed a highly significant difference (p<0.01) in winter hardiness of hybrids, lines, tester and lineHtester in 2001-2002 grown seasons.
Table 4 presets the data from a screening analysis of parents, F1 and F2 population cold tolerance and score in the field. The survival percentage ranged from 6.62 to 100.00% among the parents and the survival percentage of hybrids was 100.00%. The percentage survival of F2 populations which were found to be cold tolerant and of the most susceptible lines is given in Table 4. The most tolerant was B1, B6, B12 lines and F1 hybrids. In the F2 populations, the survival percentage ranged from 31.10 to 100.00%. The highest cold tolerant was obtained to SprinterxB12 hybrids.
General Combining Ability (GCA) and Specific Combining Ability (SCA)
The estimations of the variances of general combining ability (σ2GCA), specific combining ability (σ2SCA) additive and dominance are presented in Table 3. The differences of σ 2GCA are mainly due to the additive variance in the population whilst the differences of σ2SCA are attributable to the non-additive genetic variance (Falconer, 1982). The σ2SCA variance was higher than σ 2GCA variance for winter hardiness. This results show that non additive gene effects were important in the inheritance for winter hardiness, it revealed over dominance effect on winter hardiness.
There has been some of the ambiguity in the literature with respect to the inheritance of winter hardiness. Cousion et al. (1985) observed intermediate dominance, while Auld et al. (1983) observed additive genes. In contrast, this result confirms nature of dominance and non-additive genes for winter hardiness. Liesenfeld et al. (1986) obtained three or four genes or linkage groups for winter hardiness. This study showed that the over-dominant effects and non-additive effects were important in the inheritance of winter hardiness.
General and specific combining ability for cold hardiness varied significantly among parents and hybrids (Table 4). Sprinter showed highly significant positive GCA for cold hardiness (7.591), while Carina had significant negative GCA for cold hardiness. B12 line had the largest significant positive GCA for cold hardiness at F2 populations. B1 line had significant negative GCA for cold hardiness. Considering SCA effects, CarinaxB12 and CarinaxB1 had the highest (11.853**) and the lowest (-19.259**) SCA effects for cold hardiness at F2 populations. SprinterxB1, SprinterxB12, BoleroxB1, ManuelxB6 and CarinaxB6 had positive SCA effects for cold hardiness; while others F2 populations had negative SCA effects.
GCA for males was greater than for females in cold hardiness, reflecting the greater diversity among the male parents. Among the parents, Sprinter cultivar and B6 line were chosen as source of cold hardiness for this breeding programme. In this study, two crosses showed significant positive SCA for cold hardiness and two crosses showed a significant negative SCA effect in F2 population, indicating non-additive gene action. Of the 12 F2 progenies analyzed, SprinterxB1, SprinterxB12, ManuelxB6, CarinaxB6 and CarinaxB12 confirmed their cold hardiness (Table 4). All selected progenies will be advanced in the next generation. The results obtained to in this breeding programme are encouraging. Selected cold hardiness breeding lines for grain yield is better than the best control used in the programme under the field (not show here).
| Table 4: | General and specific combining ability and heterosis (%) values over mid-parent (HMP) and better-parent (HBP) related to cold hardiness in pea |
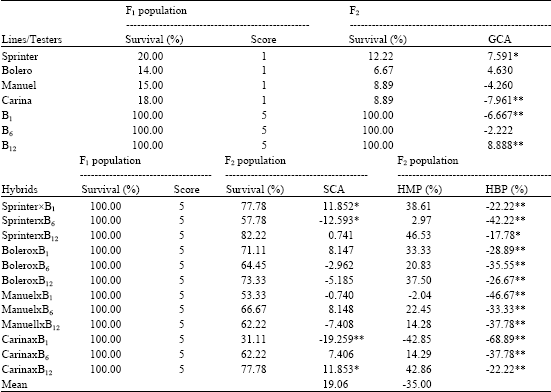 | |
| *: p<0.05. ** : p<0.01 | |
Heterosis over Mid-parent (HMP) or the Better Parent (HBP)
The average values of HMP (19.06%) were found positive in cold hardiness. In contrast, the average values of HBP (% -35.00) were found negative in cold hardiness. These values ranged from -42.88 and -68.89% (HMP and HBP, respectively), for CarinaxB1 cross, to 46.53 and -17.78%, for SprinterxB12 cross. Most of the values of HMP for cold hardiness were positive and insignificant, while the values of HBP for cold hardiness were all negative and significant (Table 4).
Table 4 gives the values of broad and narrow sense heritability for the cold hardiness. Broad and narrow sense heritability of cold hardiness was 0.85 and 0.01. Cold hardiness had the greatest heritability for broad sense heritability. Broad sense heritability was high due to the major role of environmental factors in expression of this trait. The narrow sense heritability of cold hardiness was relatively high suggesting that selection of cold hardiness in early generations would not be possible.
In conclusion, these results show that pea hybrids will be used successfully to improve cold hardiness. From the results of the breeding programme, we were able to improve some cold hardiness in pea cultivars. The hybrids were characterized by higher cold hardiness than the parents. In addition, new germplasm for useful morphological and agronomic traits were improved which will be used for combining different desirable traits in breeding programmes.
ACKNOWLEDGEMENT
The data of this study were accepted as Ph.D. Thesis of Dr. Ercan Ceyhan in 26.12.2003 by Graduate School of Natural and Applied Science.
REFERENCES
- Auld, D.L., K.J. Adams, J.B. Swensen and G.A. Murray, 1983. Diallel analyses of winter hardiness in peas. Crop Sci., 23: 763-766.
Direct Link - Guye, M.G., L. Vigh and J.M. Wilson, 1987. Recovery after chilling an assessment of chill tolerance in Phaseolus ssp. J. Exp. Bot., 38: 691-701.
CrossRef