
Soad E. Hassan
Department of Parasitology and Animal Diseases, National Research Centre, Dokki, Giza, Egypt
LiveDNA: 20.10583
Sanaa K.A. Abou-El-Dobal
Department of Parasitology and Animal Diseases, National Research Centre, Dokki, Giza, Egypt
Eman H. Abdel-Rahman
Department of Parasitology and Animal Diseases, National Research Centre, Dokki, Giza, Egypt
LiveDNA: 20.1117
Research Journal of Parasitology
Year: 2016 | Volume: 11 | Issue: 3 | Page No.: 67-72







ABSTRACT
Background and Objective: Trichostrongylus colubriformis is a common nematode which infect livestock and can cause accidental infection in humans. In the present study, affinity purification process succeeded in the isolation of a single specific fraction identified by experimentally infected rabbit sera. Methodology: The isolated fraction detected T. colubriformis antibodies in 68 (77.3%) out of 88 examined sheep serum samples. By SDS-PAGE, the isolated fraction showed simple electrophoretic profile and resolved into three bands of molecular weights 74, 85 and 97 kDa. While eight bands of molecular weights ranged from 25.7-189 kDa were detected in crude adult worm extract. In immunoblot assay, the three bands of isolated fraction were reacted with infected rabbit serum compared with four bands of molecular weights 41, 74, 85 and 97 kDa in crude extract. Results: Concerning, humeral immune responses of infected rabbits, the level of IgG antibodies increased to reach (0.526 OD±0.0027) in the first-week post infection. Then decreased to reach (0.34 OD±0.02) at the end of the experiment using ELISA. A significant expression of IL4 was observed and reached the highest level (225±0.012) at second-week post infection. While, IFN-ɣ showed less and nearly constant expression through out the time of the experiment. Conclusion: The partially immuno-affinity purified fraction of T. colubriformis adult antigen proved high diagnostic efficacy in diagnosis of trichostrongylosis in sheep. In addition, T. colubriformis infection stimulates rabbits cellular and humeral immune responses IL4, IFN-γ and IgG which, possibly create some level of protection to re infection.
PDF Abstract XML References Citation
Received: May 05, 2016;
Accepted: May 20, 2016;
Published: September 15, 2016
How to cite this article
Soad E. Hassan, Sanaa K.A. Abou-El-Dobal and Eman H. Abdel-Rahman, 2016. Diagnosis of Sheep Trichostrongylosis Based on Imune Response Profile in Experimental Rabbit Infection. Research Journal of Parasitology, 11: 67-72.
URL: https://scialert.net/abstract/?doi=jp.2016.67.72
URL: https://scialert.net/abstract/?doi=jp.2016.67.72
INTRODUCTION
Gastrointestinal nematodes are one of the most restrictions on ruminant production in the world. It causes a reduction in nutrient availability to the host through both reducing optional feed intake and/or reductions in the efficiency of utilization of absorbed nutrients (Dynes et al., 1998). This lead to weight loss, anemia, edema and reduction in the resistance to other diseases. Annual treatment costs due to these parasites have been reached to $46 and $103 million in South Africa (Waller and Chandrawathani, 2005) and in India (McLeod, 2004), respectively. Haemonchus contortus and Trichostrongylus colubriformis are the most important gastrointestinal nematodes of sheep (Rocha et al., 2008). Because, H. contortus is the most pathogenic parasite it had been focused in scientific research in tropical and sub tropical countries. In contrast little attention has been concerned T. colubriformis which proved a reduction in animal performance caused by the damages in the intestinal mucosa, and by immunopathological changes associated with the infection (Cardia et al., 2011). This loss associated with T. colubriformis infection must be oriented the scientific study to including this neglected nematode. The diagnosis of this gastrointestinal nematode mostly depends on the detection of eggs which observed in feces after the prepatent period. So, a trustworthy serological assays such as ELISA which enables the detection of host immune responses at early phase of infection is very important (Lone et al., 2012).
Hosts have developed a large variety of immune components and functions to recognize and target different parasite life stages and their products (Murphy et al., 2011). In turn, parasites display different mechanisms to evade or modify the immune response of the host to persevere and survive in the host (Schmid-Hempel, 2009). Generally, helminth infections lead to polarization of the immune system towards a Th2 response which is responsible for the production of IL-4, IL-5, IL-13 in addition to IgE and eosinophilia (Anthony et al., 2007; Allen and Maizels, 2011). In a few exceptions, infection with some tissues helminths directed the immune system towards a Th1 response in particular during specific tissue phase of the infection process (Mulcahy et al., 2005; Zhang et al., 2005; Fabre et al., 2009). These cells are responsible for the production of IFN-γ and TNF-α and enhance the activation of macrophages which are responsible for the production of antibodies (Abou-El-Dobal et al., 2015). Mean while, mixed Th1/Th2 response was found toward Trichostrongylus retortacformis in experimetally infected rabbits (Murphy et al., 2013). So, infected rabbits proved increase in the level of IFN-γ, IL-4, IgG, IgA and eosinophilia.
The current study aims to evaluate T. colubriformis partially purified fraction in the diagnosis of natural sheep trichostrongylosis after following up its effect on the level of IgG in experimentally infected rabbit sera at different weeks post infection. Rabbit IL-4 and IFN-γ levels will also be determined.
MATERIALS AND METHODS
Animals: Adult 10 male New Zealand parasite-free rabbits weighing 3-5 kg were fed ad libitum and maintained at 25°C in a conditioned vivarium in the animal house of the Veterinary Division, National Research Centre. Adult 88 males and females out-bred sheep weighing 40-50 kg were supplied by local slaughterhouse. All procedures related to animal experimentation met the International Guiding Principles for Biomedical Research involving animals as issued by the International Organizations of Medical Sciences.
Parasites: Infective larvae of T. colubriformis field strain were obtained from fecal cultures of sheep harboring T. colubriformis infection (Langrova and Jankovska, 2004). A single dose of 5000 freshly cultured infective larvae was administered orally for each rabbit. Twenty days after infection, infected rabbits were slaughtered and adult worms of T. colubriformis were harvested from small intestine using Baerman technique (Baerman and Wetzal, 1953). Larvae and worms of T. colubriformis were identified according to Soulsby (1986).
Preparation of somatic worm antigen: Adult worms of T. colubriformis were washed several times in saline chopped and homogenized in PBS pH 7.2. Particulate materials were removed by centrifugation at 14000 rpm for 30 min the supernatant was decanted and used as somatic adult antigen according to the method of Murphy et al. (2011). Protein concentration was determined by the method of Lowry et al. (1951).
Antibody-sepharose 4B affinity column chromatography: Rabbits experimentally infected sera were dialyzed against 100 mM NaHCO3 buffer pH 8.3 containing 500 mM NaCl and 0.02% NaNO3 and coupled to cyanogen bromide-activated sepharose-4B (CNBr-sepharose 4B) at the ratio of 2 mg mL–1 swollen beads by strictly following the manufacturer instructions. Trichostrongylus colubriformis mature worm extract was applied to the column. The column was washed with 0.015 M PBS containing 0.02% NaNO3 pH 8 and bound materials were eluted with 50 mM glycine-500 Mm NaCl 0.02% w/v NaNO3 pH 2.3. The protein content of isolated fraction was measured according to the method described by Lowry et al. (1951).
SDS-polyacrylamide gel electrophoresis (SDS-PAGE): The SDS-PAGE was performed in polyacrylamide gels according to Laemmli (1970). Both crude antigen and bound fraction were separately mixed with sample buffer containing 5% 2-mercaptoethanol before loading to the gel. After separation, the gel was fixed in 50% methanol and stained with silver stain according to the method of Wray et al. (1981). Relative molecular weights of bands were calculated using marker supplied by Fermentas International INC, Canada.
Immunoblot: The assay was utilized to identify the immunoreactive components recognized in the crude and purified antigens using T. colubriformis rabbits experimentally infected sera. After electrophoresis, protein components of the two antigens were immunoblotted onto nitrocellulose membrane according to the method of Towbin et al. (1979). The nitrocellulose membrane was incubated with positive rabbit sera (diluted at 1:50) and anti-rabbits IgG (whole molecule) peroxidase conjugate (diluted 1:3000) as a second antibody. The ECL Western blotting detection reagents (Amersham, UK) were utilized to visualize the immunoreactive bands.
Enzyme linked immunosorbent assay: The ELISA was adopted to evaluate the success of the purification process by determination of the antigenic activities of the eluted fraction compared with crude extract in a detection of IgG level in experimentally infected rabbits at different weeks post infection according to Engvall and Perlmann (1971). In brief, plates were coated, separately with 5 μg mL–1 isolated fraction and crude extract in carbonate buffer. Positive and negative rabbit serum samples diluted at 1:100 was added to the coated plates separately. Anti-rabbit IgG horse radish peroxidase labeled-conjugate (1:1000) and ortho-phenylenediamine (OPD) 1 mg mL–1 substrate buffer (Sigma) were used. Also, ELISA was adopted to the diagnosis of trichostrongylosis among sheep using Trichostrongylus colubriformis affinity purified fraction. The optimum antigen concentration, antibody and conjugate dilutions were determined by checker board titration. Plates were read spectrophotometrically at 450 nm and the cut off values of Optical Densities (OD) were calculated according to the method of Woo et al. (2001).
Evaluation of cytokines level: Serum levels of IL-4 and IFN-γ were measured with double-antibody sandwich ELISA kit (Glory Science Co., Ltd, Del, Rio, Tx 78840, USA). The concentration was calculated from the standard curve that was performed in the same assay.
Statistical analysis: Data are expressed as Mean±SD. Comparison between the mean values of different parameters in the studied groups was performed using 1-way ANOVA test.
RESULTS
Electrophoretic profile of Trichostrongylus colubriformis extracts: By SDS-PAGE 10% slab gel under reducing condition, the isolated fraction proved simple electrophoretic profile and resolved into three bands of molecular weights 74, 85 and 97 kDa. While the crude extract resolved into 8 bands of molecular weights ranged from 25.7-189 kDa (Fig. 1).
Immunogenic components of Trichostrongylus colubriformis extracts: By immunoblot assay and rabbit positive sera the isolated fraction proved three immunogenic bands of molecular weights 74, 85 and 97 kDa, while four bands of molecular weights 41, 74, 85 and 97 kDa were detected in crude extract (Fig. 2).
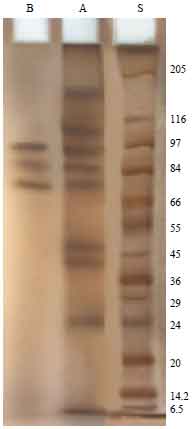 | |
| Fig. 1: | ELectrophoretic profile of T. colubriformis extract. Lane A: Crude antigen, lane B: Isolated fraction and lane S: Molecular weight standards |
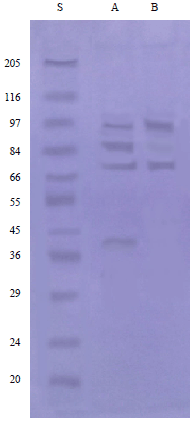 | |
| Fig. 2: | Immunoblotting assay showing reactivity of T. colubriformis extract with positive rabbit serum. Lane A: Crude antigen, lane B: Isolated fraction and lane S: Molecular weight standards |
Immune response of rabbits experimentally infected with Trichostrongylus colubriformis
Humoral response: The IgG response due to T. colubriformis infection was detected by ELISA using isolated fraction compared with crude extract. As depicted in Fig. 3, the isolated fraction has the most diagnostic potential. The level of antibodies increased to reach (0.526 OD±0.0027) in the 1st week post infection. Following this activity period, there is a little or no change in immune response from the 2nd week to 5th week post infection. The response decreased in 6 week post infection and reached a plateau until 8 week then decreased to reach (0.34 OD±0.02) at the end of the experiment.
Cellular response: Level of IL-4 increased in rabbit sera with the infection course and reached the highest level (225±0.012) at 2nd week post infection. Then slightly decreased to reach (128.750.013) at the end of the experiment. While IFN-γ showed constant expression over the time of the experiment although there was significant higher level in infected animals compared to control ones (Fig. 4).
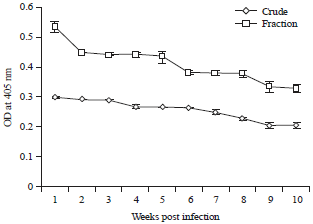 | |
| Fig. 3: | Mean level of IgG antibodies in experimentally infected rabbit sera at different weeks post infection |
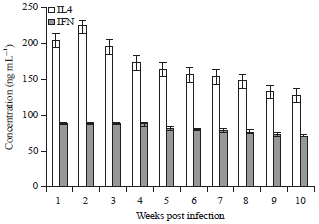 | |
| Fig. 4: | Mean level of IL-4 and IFN-γ in experimentally infected rabbit sera at different weeks post infection |
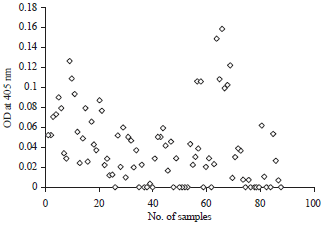 | |
| Fig. 5: | Scatter graph represents the efficacy of T. colubriformis isolated fraction in the diagnosis of Trichostrongylosis in sheep |
Diagnostic potential of Trichostrongylus colubriformis isolated fraction: As shown in Fig. 5, the isolated fraction succeeded in detection of T. colubriformis antibodies in 68 (77.3%) out of 88 examined sheep serum samples.
DISCUSSION
In the present study, rabbits were selected as lab animals based on the previous study suggested similarity between rabbits and ruminants immune response toward T. colubriformis infection (Bezubik et al., 1988; Langrova and Jankovska, 2004). These in addition to low price and easy manipulation in experimental infection. In current study examined immune response of rabbits to experimental infection with gastrointestinal T. colubriformis. The infection stimulated rabbits immune system and directed it toward Th2 activation more than Th1. So, infected animals exhibited strong anti-inflammatory cytokine (IL-4) which reached its highest level at secondary week post infection, while IFN-γ proved less and nearly constant expression level during the experiment. The current result was coincided with Anthony et al. (2007) and Allen and Maizels (2011), who proved that helminths infection mainly directed immune system toward a Th2 response. These cells characterized by high production of IL-4, IL-5, IL-13 in addition to IgE and eosinophilia. Also, these results were confirmed by Abou-El-Dobal et al. (2015) in experimental infection of rabbits with Fasciola gigantica, where IL-4 showed higher level compared to IFN-γ.
In previous study to follow up the immune response of rabbits experimentally infected with T. retortaeformis, Th1 was firstly activated and animal proved an initial strong inflammatory response (IFN-γ) followed by increase in IL-4 expression (Murphy et al., 2011, 2013). They contributed this firstly strong inflammatory response to bacterial invasion into the small intestine mucosa following movement of larvae into the tissue. Actually, this relative increase in IL-4 in presence of IFN-γ indicated that both immune phenotypes can work and this coincided with that reported in the present study.
In the current study a single dose of 5000 T. colubriformis infective larvae not only induced cellular immune response but also humeral immune response represented by high level of IgG antibodies which detected at first week post infection and remained high compared to the control animals throughout the experiment . Comparable humeral immune response was previously reported in rabbits experimentally infected with T. retortaeformis (Murphy et al., 2011). The high level of IgG in infected rabbits, throughout the experiment, directed toward isolated fraction proved success of both experimental infection in rabbits and purification process of crude extract. So, the isolated fraction was utilized in the diagnosis of trichostrongylosis in sheep, it succeeded in the detection of antibodies in 68 (77.3%) out of 88 examined sheep serum samples. A comparable infection percentage (79.8%) was detected in goats in Malaysia although they used PCR (Tan et al., 2014), this confirmed the sensitivity of utilized antigen and technique. While, less prevalence of Trichostrongylus (7.33%) was detected in Pakistan by ELISA using somatic crude antigen (Razzaq et al., 2013). The difference in infection percentage may be due to difference in utilized antigen, where they used crude antigen but in the current study partially purified isolated fraction was utilized.
By SDS-PAGE, the isolated fraction exhibited simple electrophoretic profile and resolved into 3 bands of molecular weights 74, 85 and 97 kDa compared with 8 bands of molecular weights ranged from 25.5-189 kDa were detected in the crude extract. To our knowledge, there no electrophoretic profile concerned adult T. colubriformis but in the related nematode, Haemonchus contortus, four bands of different molecular weights 66, 40, 33 and 26 kDa were detected (Tak et al., 2015).
Three bands of molecular weights 74, 85 and 97 kDa were detected in the isolated fraction by immunoblot with rabbit sera experimentally infected with T. colubriformis, while four bands of molecular weights 41, 74, 85 and 97 kDa were detected in crude extract. In the previous study, a 31 kDa allergen was detected in T. colubriformis third larval stage with IgE purified from infected sheep serum (Shaw et al., 2003).
CONCLUSION
Trichostrongylus colubriformis infection stimulates rabbits immune system and appear to elicit an unequivocal Th2 based immune response. Also, the present study introduced partially purified fraction that can be successfully utilized in the diagnosis of trichostrongylosis in sheep after proving success in rabbits. It deserves further purification to use in the diagnosis of this parasite in farm animals on large scale.
REFERENCES
- Abou-El-Dobal S.K.A., S.E. Hassan, N.I. Toaleb, A.G. Hegazi and E.H. Abdel-Rahman, 2015. Induction of protective cellular and humoral responses against fasciolosis in rabbits using immunoaffinity fraction of Fasciola gigantica excretory secretory products. J. Clin. Cell. Immunol.
CrossRefDirect Link - Allen, J.E. and R.M. Maizels, 2011. Diversity and dialogue in immunity to helminths. Nat. Rev. Immunol., 11: 375-388.
CrossRefDirect Link - Anthony, R.M., L.I. Rutitzky, J.F. Urban Jr., M.J. Stadecker and W.C. Gause, 2007. Protective immune mechanisms in helminth infection. Nat. Rev. Immunol., 7: 975-987.
CrossRefDirect Link - Cardia, D.F.F., R.A. Rocha-Oliveira, M.H. Tsunemi and A.F.T. Amarante, 2011. Immune response and performance of growing Santa Ines lambs to artificial Trichostrongylus colubriformis infections. Vet. Parasitol., 182: 248-258.
CrossRefDirect Link - Dynes, R.A., D.P. Poppi, G.K. Barrell and A.R. Sykes, 1998. Elevation of feed intake in parasite-infected lambs by central administration of a cholecystokinin receptor antagonist. Br. J. Nutr., 79: 47-54.
CrossRefDirect Link - Engvall, E. and P. Perlmann, 1971. Enzyme-Linked Immunosorbent Assay (ELISA) quantitative assay of immunoglobulin G. Immunochemistry, 8: 871-874.
CrossRefPubMedDirect Link - Fabre, V., D.P. Beiting, S.K. Bliss, N.G. Gebreselassie and L.F. Gagliardo et al., 2009. Eosinophil deficiency compromises parasite survival in chronic nematode infection. J. Immunol., 182: 1577-1583.
Direct Link - Schmid-Hempel, P., 2009. Immune defence, parasite evasion strategies and their relevance for macroscopic phenomena such as virulence. Philos. Trans. R. Soc. London B: Biol. Sci., 364: 85-98.
CrossRefDirect Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Langrova, I. and I. Jankovska, 2004. Arrested development of Trichostrongylus colubriformis in experimentally infected rabbits. Effect of decreasing photoperiod, low temperature and desiccation. Helminthologia, 41: 85-90.
Direct Link - Lone, B.A., M.Z. Chishti, F. Ahmad, H. Tak and J. Hassan, 2012. Immunodiagnosis of Haemonchus contortus infection in sheep by indirect Enzyme Linked Immunosorbent Assay (ELISA). Iran. J. Vet. Res., 13: 49-53.
Direct Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Mulcahy, G., S. O'Neill, J. Fanning, E. McCarthy and M. Sekiya, 2005. Tissue migration by parasitic helminths-an immunoevasive strategy? Trends Parasitol., 21: 273-277.
CrossRefDirect Link - Murphy, L., N. Nalpas, M. Stear and I.M. Cattadori, 2011. Explaining patterns of infection in free-living populations using laboratory immune experiments. Parasite Immunol., 33: 287-302.
CrossRefDirect Link - Murphy, L., A.K. Pathak and I.M. Cattadori, 2013. A co-infection with two gastrointestinal nematodes alters host immune responses and only partially parasite dynamics. Parasite Immunol., 35: 421-432.
CrossRefDirect Link - Razzaq, A., K. Ashraf, A. Maqbool, M. A. Khan, M. Islam and H. Khan, 2013. Epidemiology, serodiagnosis and therapeutic studies on ovine nematodes at district Loralai, Balochistan, Pakistan. J. Anim. Plant Sci., 23: 1559-1565.
Direct Link - Rocha, R.A., K.D.S. Bresciani, T.F.M. Barros, L.H. Fernandes, M.B. Silva and A.F.T. Amarante, 2008. Sheep and cattle grazing alternately: Nematode parasitism and pasture decontamination. Small Rumin. Res., 75: 135-143.
CrossRefDirect Link - Shaw, R.J., M.M. McNeill, D.R. Maass, W.R. Hein and T.K. Barber et al., 2003. Identification and characterisation of an aspartyl protease inhibitor homologue as a major allergen of Trichostrongylus colubriformis. Int. J. Parasitol., 33: 1233-1243.
CrossRefDirect Link - Tak, I.U.R., M.Z. Chishti and F. Ahmad, 2015. Protein profiling of Haemonchus contortus found in sheep of Kashmir valley. J. Parasitic Dis., 39: 639-644.
CrossRefDirect Link - Tan, T.K., C. Panchadcharam, V.L. Low, S.C. Lee, R. Ngui, R.S.K. Sharma and Y.A.L. Lim, 2014. Co-infection of Haemonchus contortus and Trichostrongylus spp. among livestock in Malaysia as revealed by amplification and sequencing of the internal transcribed spacer II DNA region. Vet. Res., Vol. 10.
CrossRefDirect Link - Towbin, H., T. Staehelin and J. Gordon, 1979. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA., 76: 4350-4354.
PubMedDirect Link - Waller, P.J. and P. Chandrawathani, 2005. Haemonchus contortus: Parasite problem No. 1 from tropics-polar circle. Problems and prospects for control based on epidemiology. Trop. Biomed., 22: 131-137.
Direct Link - Woo, P.C.Y., A.S.P. Leung, S.K.P. Lau and K.T.K. Chong, 2001. Use of recombinant mitogillin for serodiagnosis of Aspergillus fumigatus-associated diseases. J. Clin. Microbiol., 39: 4598-4600.
CrossRefDirect Link - Wray, W., T. Boulikas, V.P. Wray and R. Hancock, 1981. Silver staining of proteins in polyacrylamide gels. Anal. Biochem., 118: 197-203.
CrossRefDirect Link - Zhang, W.Y., E. Moreau, J.C. Hope, C.J. Howard, W.Y. Huang and A. Chauvin, 2005. Fasciola hepatica and Fasciola gigantica: Comparison of cellular response to experimental infection in sheep. Exp. Parasitol., 111: 154-159.
CrossRefDirect Link