
Sobhy Abdel- Shafy
Department of Parasitology and Animal Diseases, Veterinary Research Division, National Research Center, Giza, Egypt
Rafaat M. Shaapan
Department of Zoonotic Diseases, Veterinary Research Division, National Research Center, Giza, Egypt
Khaled A. Abdelrahman
Department of Parasitology and Animal Diseases, Veterinary Research Division, National Research Center, Giza, Egypt
Amira H. El- Namaky
Department of Parasitology and Animal Diseases, Veterinary Research Division, National Research Center, Giza, Egypt
Faten A.M. Abo- Aziza
Department of Parasitology and Animal Diseases, Veterinary Research Division, National Research Center, Giza, Egypt
Hala A. Abou Zeina
Department of Parasitology and Animal Diseases, Veterinary Research Division, National Research Center, Giza, Egypt
Research Journal of Parasitology
Year: 2015 | Volume: 10 | Issue: 4 | Page No.: 142-150







ABSTRACT
The brown dog tick Rhipicephalus sanguineus is a three-host tick that feeds primarily on dog and occasionally on other hosts, including human. Toxoplasmosis is generally considered the most important disease that causing abortion of both pregnant women and different female animals throughout the world. Therefore, the aim of this study was to determine the ability of the brown dog tick R. sanguineus to acquire Toxoplasma infection through feeding its larvae on experimentally infected rabbits with T. gondii. The R. sanguineous larvae were feed on rabbits experimentally infected with locally isolated virulent Toxoplasma gondii strain. Nymphs moulted from these larvae were investigated to detect the presence of T. gondii specific B1 gene DNA using PCR at 193 bp. Histological examination for liver, lung and heart of experimentally infected rabbits was performed to confirm the infection of animals with T. gondii. The histopathological examination of infected rabbit tissues (heart, lung liver) revealed infiltration of T. gondii tachyzoites and polymorph nuclear inflammatory cells in addition the presence of different tissue alterations and degeneration other than that in normally histological tissues of non-infected rabbits. The nymphs of R. sanguineus were free from T. gondii. Further investigations are needed on the other developmental stages of R. sanguineus to ensure the ability of this tick species in transmission of T. gondii.
PDF Abstract XML References Citation
Received: August 18, 2015;
Accepted: October 01, 2015;
Published: October 29, 2015
How to cite this article
Sobhy Abdel- Shafy, Rafaat M. Shaapan, Khaled A. Abdelrahman, Amira H. El- Namaky, Faten A.M. Abo- Aziza and Hala A. Abou Zeina, 2015. Detection of Toxoplasma gondii (Apicomplexa: Sarcocystidae) in the Brown Dog Tick Rhipicephalus sanguineus (Acari: Ixodidae) Fed on Infected Rabbits. Research Journal of Parasitology, 10: 142-150.
URL: https://scialert.net/abstract/?doi=jp.2015.142.150
URL: https://scialert.net/abstract/?doi=jp.2015.142.150
INTRODUCTION
Rhipicephalus sanguineus, commonly known as the brown dog tick, is a three-host tick that feeds primarily on dogs. As other ixodid ticks, R. sanguineus undergo four developmental stages: Egg, larva, nymph and adult. Immature stages can be found on rodents and other small mammals. Adults usually parasitize larger animals, including humans. Adult females of R. sanguineus feed on the host for 5-21 days, drop off and lay eggs in sheltered places. After incubation, small larvae hatch from the eggs and immediately begin to look for a host. Larvae feed for 3-10 days, before drop off the host to molt into nymphs. The larva molting period ranges from 5-15 days. Nymphs resemble adults in form and feed for 3-11 days before drop off the host to develop into adults. Unfed R. sanguineus larvae can survive for approximately eight months, whereas, unfed nymphs and adults can survive for 6 and 19 months. R. sanguineus ticks are widely distributed around the world and they are known vectors of pathogens, such as Babesia canis, Ehrlichia canis and Rickettsia conorii (Dantas-Torres, 2008).
Toxoplasma gondii is a single-celled parasite of all warm-blooded hosts that is distributed worldwide. The organism can infect a wide range of animal species and man as an intermediate hosts. Cats are the definitive hosts which are the main reservoirs of the infection because they are the only hosts that can excrete the resistant stage (oocysts) of the parasite in the feces. Humans become infected by eating undercooked meat from infected animals or ingestion of food and water contaminated with oocysts. The infection causes mental retardation and loss of vision in children and abortion in pregnant women and livestock (Sreekumar et al., 2005; Shaapan, 2015). Once the T. gondii tachyzoites infective stages were invade host they multiply by endodyogony in host tissue and might form pseudo cysts filled with crescent or banana shaped centrally located nucleus and when cysts disintegrated it spread by host blood lead to parasitemia and invade other cells. The tachyzoites might be engulfing by phagocytic cells widely distributed throughout the body (Shaapan et al., 2008; Dubey, 2010).
A little known literature studied the presence of Toxoplasma gondii in a few species of tick such as Ornithodoros spp., Ixodes spp. and Amblyomma spp. and their ability to transmit T. gondii. Jagow and Hoffmann (1970) found that Toxoplasma lived in nymph and adult of Ornithodoros moubata up till 10 and 2 days, respectively. They added that transmission was unsuccessful either through sucking, or with the next stage of the same ticks or through the F1-generation of the ticks infected with Toxoplasma. Moreover, Castellani (1970) studied the transmission of Toxoplasma gondii by ticks of the Ornithodorus genus. However, Gidel and Provost (1965) isolated Toxoplasma gondii from naturally infected Ixodidae of the genus Amblyomma. In Egypt, a unique study conducted by Rifaat et al. (1963), who studied natural Toxoplasma infection sought in bloodsucking arthropods at Aburawash, Giza. In experiments on the possibility of natural infection of tick females, nymphs and larvae of Ixodes ricinus during sucking on animals having toxoplasmosis, a positive result was obtained only in nymphs (Derylo et al., 1977). They had obtained a positive result only with nymphs when they carried out experiments to determine the possibility of nymphs and females of I. ricinus transmitting the invasion by intermittent sucking.
Recently, Polymerase Chain Reaction (PCR) is used in detection of Toxoplasma infection in ticks. Sroka et al. (2002) detected the presence of Toxoplasma gondii DNA PCR test in 2 out of 92 Ixodes ricinus ticks (2.8%) collected in the woodlands of eastern Poland. Therefore, they suggested that ticks of this species may be involved in the spread of toxoplasmosis under natural conditions. Sroka et al. (2008) examined a batch of 259 Ixodes ricinus ticks collected by flagging in woodlands of North-Western Poland (Szczecin area) for the presence of Toxoplasma gondii DNA with nested PCR. They revealed that 12.7% of I. ricinus ticks were infected with T. gondii. They homogenized and inoculated into mice another batch of 340 I. ricinus ticks from Szczecin area. Subsequently, the biological material obtained from inoculated mice was also examined with nested PCR for the presence of Toxoplasma gondii DNA. Samples from brains and other organs of 60 inoculated mice showed 44 positive results for T. gondii DNA. Sroka et al. (2009) examined a total of 715 Ixodes ricinus ticks from Eastern Poland for the presence of Toxoplasma gondii, using PCR and nested-PCR methods. They recorded 12.6% tick infection with T. gondii.
Although, the public health importance of T. gondii in worldwide and there is a lack of data on its spreading through ticks. Therefore, the aim of this study was to determine the ability of the brown dog tick R. sanguineus to acquire Toxoplasma infection through feeding its larvae on rabbits experimentally infected with Toxoplasma gondii.
MATERIALS AND METHODS
Toxoplasma gondii strain: Local virulent T. gondii strain was isolated from tissues of Egyptian slaughtered sheep and maintained by intra peritoneal propagation in mice according to the procedures described by Shaapan and Ghazy (2007). The T. gondii strain was used for experimental infection of rabbits and as control strain in PCR detection.
Rhipicephalus sanguineus tick: The brown dog tick Rhipicephalus sanguineus were collected from dogs in Abo Rawash village, Giza governorate during the summer season 2014. Male and female adult ticks were identified according the keys of Hoogstraal (1956) and Estrada-Pena et al. (2004). The fully engorged females were chosen and incubated at 27±2°C and 75% relative humidity in order to lay eggs that maintained in the same incubation conditions until they hatched to larvae. One week post hatching, larvae were used in the experiment.
Experimental design: Nine healthy New-Zeeland white rabbits were used in the experimental infection with T. gondii. The range of weight was 2.0-2.5 kg per rabbit. The rabbits were divided into three groups, 3 rabbits of each and kept under insect free condition.
First group: This group was experimentally infected by subcutaneous inoculation with the tachyzoites of the T. gondii strain using method of Sharma and Dubey (1981) the tachyzoites were freshly obtained from pooled peritoneal exudates recovered from previously inoculated mice 2-3 days earlier and counted using haemocytometer to determine their count per mL. The dose of infection for each rabbit was 2×106 tachyzoites. This first group of rabbit was infested by R. sanguineus larvae according the methodology of Abdel-Shafy et al. (2008) at the day rabbits injected with T. gondii. Larvae were placed inside a feed capsule consisting of a plastic tube (2.5 cm of diameter and 3 cm of height) glued on the shaved backs of rabbits (2 capsules for each). Wooden collar used on rabbits to prevent grooming (Szabo and Bechara, 1997). A total of 200 larvae (100 for each capsule) were fed on each rabbit. Larvae were initially checked at 72 day post infestation to observe their feeding and dropping.
Second group: This group was experimentally infected by T. gondii strain by the way described in the first group and maintained without tick infestation, so considered as positive control group.
Third group: This group was considered as negative control group which maintained without T. gondii infection and R. sanguineus infestation. Four days post injection with T. gondii or tick infestation, larvae dropped and incubated at the same conditions mentioned above until they moulted to nymphs. One week post moulting, nymphs were preserved in ethanol for molecular investigation.
Molecular examination
Extraction and isolation of DNA: Extraction and isolation of DNA from R. sanguineus nymph in which DNA was purified by using a QIAamp blood kit (Qiagen Inc.) according to the manufacturer’s protocol. Briefly, the tissues were lysed and digested with 1.8 mg of Qiagen’s protease per ml for 2 h at 70°C, after which the samples were applied to DNA purification spin columns. The columns were washed twice and the DNA was eluted from the columns with 50 μL TE buffer. While, the extraction and isolation of DNA from ticks was the same procedure except increasing the lysis duration to be overnight lysis. A 5 μL aliquot of the DNA sample was used as the template in PCR.
Detection of Toxoplasma gondii by PCR: Toxoplasma gondii DNA was initially confirmed by PCR amplification of the repetitive and conserved gene B1 (Burg et al., 1989; Ponce and Gomez, 2003). Positive control was DNA from RH strain and negative control was distilled water in presence of primers. Two oligonucleotides were synthesized on (MWV) DNA synthesizer. Oligonucleotide primers used to initiate DNA amplification were complementary to segments of the Bi gene of T. gondii (5'-GGAACTGCATCCGTTCATGAG-3' and 5'-TCQTfAAAGCGTTCGTGGTC-3'). The amplification reactions were performed with the thermostable DNA polymerase (Taq; Fermentus). Briefly, 2.5 U of Taq polymerase was used in a 100 μL reaction volume with 10 mM Tris (pH 8.3) -2.5 mM MgCl2-100 mM deoxynucleoside triphosphates (Pharmacia)-each oligo at 1.0 mM. Samples were amplified for 35 cycles in an automated PCR machine. Each cycle consisted of 1 min of denaturation at 94°C, 1 min at the annealing temperature of 55°C and 50 sec of extension at 72°C. The final extension step continued for an additional 5 min.
Histological examination: All control and infected rabbits were slaughtered at 7-10 days post infection then dissected to remove the internal organs such as liver, lung and heart. Histopathological examination was conducted according to the methodology of Drury and Wallington (1980) and Bancroft et al. (1996). Tissue specimens were taken from liver, lung, kidney and heart of each rabbit in all groups immediately after sacrifice at the end of the experiment. These tissues were then fixed in 10% formol-saline. The specimens were dehydrated, cleared and embedded in paraffin blocks. Paraffin sections of 5 μm thickness were prepared, stained with hematoxylin and eosin and examined microscopically.
RESULTS
Detection of Toxoplasma gondii in Rhipicephalus sanguineus nymphs: Toxoplasma specific B1 gene PCR amplicon was not detected on 2% agarose gel stained with ethidium bromide from R. sanguineous larvae and from rabbit organs-infected by T. gondii.
Detection of Toxoplasma gondii in rabbit organs: Histopathological studies of heart, lung and liver of non-infected and T. gondii experimentally infected rabbits through microscopical examination of paraffin embedded sections stained with hematoxylin and eosin revealed that normal histological picture heart, lung and liver tissue specimens from non-infected rabbits shown in Fig. 1a, 2a and 3a, respectively. On the other hand, infected rabbit heart revealed presence of myocytes and multi-focal interstitial myocarditis with infiltration of lymphocytes, macrophages and few neutrophils with free tachyzoites were seen in cardiac myocytes bordering the lesions and among inflammatory cells (Fig. 1b).
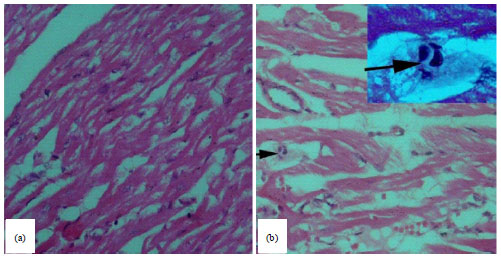 | |
| Fig. 1(a-b): | (a) No-infected and (b) Infected Toxoplasma gondii heart tissues with infiltrated Toxoplasma gondii tachyzoites (black arrow) |
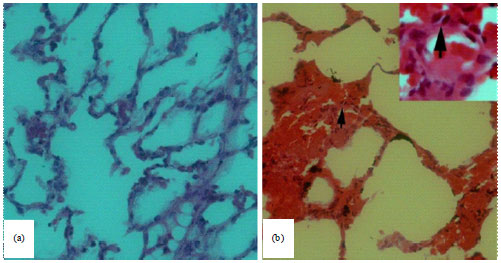 | |
| Fig. 2(a-b): | (a) No-infected and (b) Infected Toxoplasma gondii lung tissues with infiltrated Toxoplasma gondii tachyzoites (black arrow) |
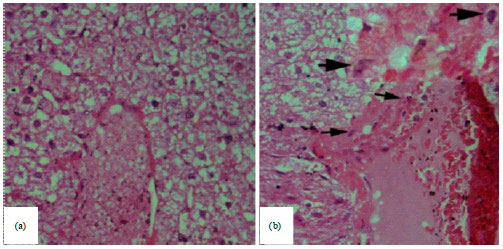 | |
| Fig. 3(a-b): | (a) No-infected and (b) Infected Toxoplasma gondii liver tissues with infiltrated Toxoplasma gondii tachyzoites (black arrow) |
Infiltration of large numbers of mononuclear cells among myocytes and foci of extravasated erythrocytes between the bronchioles were easily recognized, in addition few scattered T. gondii tachyzoites in infected rabbit lung tissue (Fig. 2b). Scattered infiltration of eosinophils singly or in small clusters and focal areas of mild degenerative changes in the form of hydropic degeneration and large number of T. gondii were also seen in infected rabbit liver tissue (Fig. 3b).
DISCUSSION
Toxoplasma gondii is an obligatory intracellular parasitic protozoan transmitted by ingestion of uncooked infected meat with tachyzoite and contaminated water or food with oocysts (Dubey, 2008). Oral transmission by consumption of raw meat is regarded as the only route of primary infection. However, this route hardly explains the common occurrence of T. gondii in a variety of hosts, such as domestic mammals and birds, which are unlikely to contract primary infection orally through meat consumption. Thus, some other transmission routes of transmission T. gondii have been considered by skin lesions and transmission by arthropods (Woke et al., 1953). Previous studies have raised the possibility of transmission by blood-sucking arthropods, in particular, ticks (Woke et al., 1953; Derylo et al., 1977; Sroka et al., 2002, 2009). Sroka et al. (2009) isolated Toxoplasma strains from naturally infected Ixodes recinus ticks suggesting that this tick species may have a role in the spread of toxoplasmosis under natural conditions. Although transmission of T. gondii by ticks is not major route for human infection, cases of human toxoplasmosis associated with tick bites have been described (Singh et al., 1965). Therefore, this study was conducted to determine the ability of R. sanguineus to acquire Toxoplasma infection through feeding its larvae on rabbits experimentally infected with T. gondii. Then, the experimentally infected larvae were allowed to moult into nymphs that were investigated by using PCR for the presence of Toxoplasma DNA. Additionally, histological examinations were performed on liver, lung and heart of infected rabbits to confirm the Toxoplasma infection.
The results of the present study clarified that the infection of rabbits with T. gondii succeeded due to the tachyzoites were observed in the cross sections of all tested organs (liver, lung and heart). Lesions found by the histopathological examination of the different tissues (heart, lung and liver) of the experimentally infected rabbits in our study found were consistent to the ones demonstrated by other authors who studied lesions in animals and man (Morales et al., 1996; Esteban-Redondo et al., 1999). Tissues harboring T. gondii tachyzoites in this study most frequently were the heart and liver tissue, but organisms were detected less frequently in the lung this in agreement with results obtained by Prickett et al. (1985), Da Silva and Langoni (2001) and Shaapan and Ghazy (2007).
The brown dog tick R. sanguineus is three-host tick and it has a chance to spread a pathogen through a wide range of hosts in a lower time comparing with other ixodid tick species infested domestic animals in Egypt. The main concept of this study was to evaluate the ability of R. sanguineus larvae to acquire Toxoplasma infection and preserved the tachyzoite alive inside their body until they moult to nymphs. To achieve this goal, infestation by larvae and infection with Toxoplasma were done together at the same time because the feeding period of larvae is not more than four days. The nymph and adult stages of R. sanguineus will be evaluated for acquiring infection with Toxoplasma in forthcoming research. Results of the present research revealed that nymph moulted from larvae fed on infected rabbits with Toxoplasma were negative when the nymph specimens were investigated by PCR. In spite of this negative finding this study is considered the first research work that evaluated the larvae of one ixodid tick species namely R. sanguineus for acquiring Toxoplasma infection. The negative result may attribute to the inoculation dose of T. gondii was low that led to the parasitemia in blood was very low. This negative finding may indicate to R. sanguineus larvae has not ability to acquire T. gondii infection. To confirm this finding, more doses of Toxoplasma should be evaluated.
The negative Toxoplasma in nymphs moulted from larvae that fed on infected rabbits with Toxoplasma means that the larvae have not ability to acquire Toxoplasma infection. This result agrees with that recorded before by Derylo et al. (1977), who found that the nymphs of Ixodes ricinus are most susceptible to infection with T. gondii and they reported that the role of ticks in transmitting of toxoplasmosis should be considered in further investigations. They also added that Toxoplasma artificially (intracoelomatically) introduced into the organism of I. ricinus females can penetrate and multiply in hypoderma, salivary glands, peritracheal connective tissue and muscular tissue. Moreover, Jagow and Hoffmann (1970) found that Toxoplasma lived in Nymphs and adults of Ornithodoros moubata up till 10 and 2 days, respectively. They also found transmission was unsuccessful either through sucking or with the next stage of the same ticks or through the F1-generation of the ticks infected with Toxoplasma. Whereas, Gidel and Provost (1965) isolated Toxoplasma gondii from to the genus Amblyomma parasiting a bovine in Centre African Republic by inoculation into rabbits and guinea pigs and proved pathogenic for mice.
The PCR techniques are most helpful tool to understand the role of ticks in transmitting Toxoplasma. Recently, a few researches were conducted using PCR on the ticks as vectors of toxoplasmosis (Sroka et al., 2002, 2009; Asman et al., 2015). All of them screened the natural infection of Ticks with Toxoplasma. Sroka et al. (2002) detected T. gondii DNA was by PCR in 2 out of 92 I. ricinus ticks (2.8%) collected in the woodlands of eastern Poland. They suggested that ticks of this species may be involved in the spread of toxoplasmosis under natural conditions. The total examined ticks (92) included nymphs, male and females which were unfed and collected by flagging technique. The two positive I. ricinus ticks were unfed females. This means that the two females were infected by the individual of the previous stage (nymph) that fed on infected host with Toxoplasma under natural conditions. Sroka et al. (2009) examined unfed 715 I. ricinus ticks collected by flagging method for the presence T. gondii, using PCR and nested-PCR methods. The overall percentage of tick infection with T. gondii was 12.6%. The highest proportions of infections (23.5%) were detected in females, among all the developmental stages (4.4% in nymphs and 13.2% in males). This means that the nymphs are most acquiring Toxoplasma infection as reported before by Derylo et al. (1977). Incomparable with the present study the nymphs revealed positive infection. This means that the larvae of I. ricinus have ability to acquire Toxoplasma infection and the infection pass to the next stage. Asman et al. (2015) collected I. ricinus ticks from vegetation and pets in the spring period of their activity on the territory of the Tarnogórski district, Poland and examined them for the presence of T. gondii using nested PCR. They found that the percent of T. gondii infection in ticks collected from vegetation were 76.10, 75 and 5.55% in females, males and nymphs, respectively in Tarnogórski area. The T. gondii infection in ticks collected from pets was 96.42 and 100% in ticks fed on dogs and cats, respectively. The vegetation results give an indication to the larvae have low ability to acquire T. gondii infection, while nymphs are more susceptible to infection with T. gondii.
CONCLUSION
All tested organs of infected rabbits (liver, lung and heart) revealed the presence of tachyzoites of T. gondii. The nymphs moulted from infected larvae of R. sanguineus were free from T. gondii. Further investigations are needed on the other developmental stages of R. sanguineus to ensure the ability of this tick species in transmission of T. gondii.
REFERENCES
- Abdel-Shafy, S., M.S. Mahmoud and M.M. Abdel-Aziz, 2008. Evaluation of crude and fractionated gut extract antigens for protection against camel tick Hyalomma dromedarii (Acari: Ixodidae). J. Entomol., 5: 91-102.
CrossRefDirect Link - Asman, M., K. Solarz, P. Cuber, T. Gasior and P. Szilman et al., 2015. Detection of protozoans Babesia microti and Toxoplasma gondii and their co-existence in ticks (Acari: Ixodida) collected in Tarnogorski district (Upper Silesia, Poland). Ann. Agric. Environ. Med., 22: 80-83.
CrossRefDirect Link - Bancroft, J., A. Stevens and D. Turner, 1996. Theory and Practice of Histological Techniques. 4th Edn., Churchill, Livingston, New York, London, San Francisco, Tokyo Pages: 740.
CrossRefDirect Link - Burg, J.L., C.M. Grover, P. Pouletty and J.C. Boothroyd, 1989. Direct and sensitive detection of a pathogenic protozoan, Toxoplasma gondii, by polymerase chain reaction. J. Clin. Microbiol., 27: 1787-1792.
PubMedDirect Link - Castellani, P.M., 1970. [Transmission of Toxoplasma gondii by ticks of the Ornithodorus genus]. Giornale di Malattie Infettive e Parassitarie, 22: 226-227, (In Italian).
PubMedDirect Link - Dantas-Torres, F., 2008. The brown dog tick, Rhipicephalus sanguineus (Latreille, 1806) (Acari: Ixodidae): From taxonomy to control. Vet. Parasitol., 152: 173-185.
CrossRefDirect Link - Derylo, A., S. Tos-Luty, J. Dutkiewicz and J. Uminski, 1977. [Participation of Ixodes ricinus L. ticks in the biology and transmission of Toxoplasma gondii]. Wiadomosci Parazytologiczne, 24: 585-595.
PubMedDirect Link - Dubey, J.P., 2008. The history of Toxoplasma gondii-the first 100 years. J. Eukaryotic Microbiol., 55: 467-475.
CrossRefDirect Link - Esteban-Redondo, I., S.W. Maley, K. Thomson, S. Nicoll, S. Wright, D. Buxton and E.A. Innes, 1999. Detection of T. gondii in tissues of sheep and cattle following oral infection. Vet. Parasitol., 86: 155-171.
CrossRefDirect Link - Gidel, R. and A. Provost, 1965. [Isolation of Toxoplasma gondii in naturally infected Ixodidae of the genus Amblyomma]. Annales de Institut Pasteur., 109: 613-616, (In French).
PubMedDirect Link - Jagow, M. and G. Hoffmann, 1970. [Experiments on the transmission of Toxoplasma gondii in various developing stages of Ornithodoros moubata]. Zeitschrift fur Parasitenkunde, 33: 246-251, (In German).
PubMedDirect Link - Morales, J.A., M.A. Pena and J.P. Dubey, 1996. Disseminated toxoplasmosis in a captive porcupine (Coendou mexicanus) from Costa Rica. J. Parasitol., 82: 185-186.
PubMedDirect Link - Prickett, M.D., D.W. Dreesen, W.D. Waltman, J.L. Blue and J. Brown, 1985. Correlation of tissue infection and serologic findings in pigs fed Toxoplasma gondii oocysts. Am. J. Vet. Res., 46: 1130-1132.
PubMedDirect Link - Rifaat, M.A., A.A. Soliman and T.A. Morsy, 1963. Natural Toxoplasma infection sought in bloodsucking arthropods at Aburawash, Giza, U.A.R. J. Trop. Med. Hygiene, 66: 152-155.
PubMedDirect Link - Shaapan, R.M., F.A. El-Nawawi and M.A.A. Tawfik, 2008. Sensitivity and specificity of various serological tests for the detection of Toxoplasma gondii infection in naturally infected sheep. Vet. Parasitol., 153: 359-362.
CrossRefDirect Link - Shaapan, R.M., 2015. The common zoonotic protozoal diseases causing abortion. J. Parasitic Dis.
CrossRef - Shaapan, R.M. and A.A. Ghazy, 2007. Isolation of Toxoplasma gondii from horse meat in Egypt. Pak. J. Biol. Sci., 10: 174-177.
CrossRefPubMedDirect Link - Sharma, S.P. and J.P. Dubey, 1981. Quantitative survival of Toxoplasma gondii tachyzoites and bradyzoites in pepsin and in trypsin solutions. Am. J. Vet. Res., 42: 128-130.
PubMedDirect Link - Da Silva, A.V. and H. Langoni, 2001. The detection of Toxoplasma gondii by comparing cytology, histopathology, bioassay in mice and the Polymerase Chain Reaction (PCR). Vet. Parasitol., 97: 193-200.
CrossRefDirect Link - Singh, I., S.M. Basu, D. Narsimhan, D.N. Sardana and C.C. Kapila et al., 1965. Haemorrhagic disease following tick bites suspected toxoplasmosis. Lancet, 285: 834-838.
CrossRefDirect Link - Sreekumar, C., D.E. Hill, K.B. Miska, M.C.B. Vianna, L. Yan, R.L. Myers and J.P. Dubey, 2005. Genotyping and detection of multiple infections of Toxoplasma gondii using pyrosequencing. Int. J. Parasitol., 35: 991-999.
CrossRefDirect Link - Sroka, J., J. Chmielewska-Badora and J. Dutkiewicz, 2002. Ixodes ricinus as a potential vector of Toxoplasma gondii. Ann. Agric. Environ. Med., 10: 121-123.
PubMedDirect Link - Sroka, J., J. Szymanska and A. Wojcik-Fatla, 2009. The occurrence of Toxoplasma gondii and Borrelia burgdorferi sensu lato in Ixodes ricinus ticks from east Poland with the use of PCR. Ann. Agric. Environ. Med., 16: 313-319.
PubMedDirect Link - Sroka, J., A. Wojcik-Fatla, J. Zwolinski, V. Zajac, M. Sawczuk and J. Dutkiewicz, 2008. Preliminary study on the occurrence of Toxoplasma gondii in Ixodes ricinus ticks from north-western Poland with the use of PCR. Ann. Agric. Environ. Med., 15: 333-338.
PubMedDirect Link - Woke, P.A., L. Jacobs, F.E. Jones and M.L. Melton, 1953. Experimental results on possible arthropod transmission of toxoplasmosis. J. Parasitol., 39: 523-532.
CrossRefDirect Link - Szabo, M.P.J. and G.H. Bechara, 1997. Immunisation of dogs and guinea pigs against Rhipicephalus sanguineus ticks using gut extract. Vet. Parasitol., 68: 283-294.
CrossRefDirect Link