
Mai A. Hegazi
Department of Parasitology, Faculty of Medicine for Girls, Al-Azhar University, Cairo, Egypt
Madiha Mahmoud
Department of Parasitology, Faculty of Medicine for Girls, Al-Azhar University, Cairo, Egypt
Research Journal of Parasitology
Year: 2013 | Volume: 8 | Issue: 2 | Page No.: 55-63







ABSTRACT
The study aimed to examine the effect of combined therapy of artemether (ART) and somatostatin (SOM) on liver fibrosis in experimentally infected mice with Schistosoma mansoni. Infected mice were investigated following administration of either SOM, ART or their combination. Each regimen was administered at 2, 5 and 13 weeks post-infection (PI) to separate groups which were respectively sacrificed at 5, 8 and 16 weeks PI. Liver fibrosis was assessed by chemical measurement of heptic hydroxyproline as a marker of collagen content in liver. The extent of infection was monitored by liver egg load. The results were interpreted in comparison with corresponding ones in untreated S. mansoni-infected mice and age-matched normal control mice. Combined treated group showed the highest significant lower mean of hepatic hydroxyproline content than the infected control in 2, 5 and 13 weeks PI (793.41±64.91, 1010.87±75.67 and 1021.42±135.60) respectively in comparison to control group. ART administration group attained nearby significant lower measures as that of combined therapy group in the three successive periods of the experiment (893.57±31.83, 1035.91±42.24, 1228.34±26.52) in comparison to control group. SOM treated group attained significant lower hydroxyproline content in only 5 weeks PI (1145.30±80.91) and 13 weeks PI (1154.53±60.84). The study concluded that the combined therapy seemed not to be more effective than ART alone. By implication SOM therapy does not appear to have an additive effect on ART treatment.
PDF Abstract XML References Citation
Received: January 25, 2013;
Accepted: May 21, 2013;
Published: July 15, 2013
How to cite this article
Mai A. Hegazi and Madiha Mahmoud, 2013. Efficacy of Combined Therapy of Artemether and Somatostatin in
Heptic Hydroxyproline in Experimental Murine Schistosomiasis
Mansoni. Research Journal of Parasitology, 8: 55-63.
URL: https://scialert.net/abstract/?doi=jp.2013.55.63
URL: https://scialert.net/abstract/?doi=jp.2013.55.63
INTRODUCTION
The main inflammatory reaction against Schistosome eggs dislodged in the hepatic portal venules is the granulomatous formation with subsequent fibrosis (Alves Oliveir et al., 2006). Hepatic fibrosis precipitates presinusoidal portal hypertension and bleeding oesophageal varices (Mahmoud, 1984; Mansy et al., 1990, 1992). The degree of hepatic fibrosis depends on the viability of Schistosome eggs and their number. Viable eggs trigger the host granulomatous reaction as miracidium in mature eggs secrete antigens while no response is observed around dead eggs (Pellegrino and Katz, 1969; Warren and Boros, 1975; Cheever et al., 1992). There is also a positive correlation between the severity of hepatic fibrosis and the number of eggs in the tissue (Yue et al., 1984). The main feature of Liver fibrosis is the accumulation of extracellular matrix proteins including collagen (Chatterjee et al., 2005). Livers contain 20 times more collagen than normal in experimental hepatosplenic schistosomiasis (Dunn et al., 1977). Among protein collagen hydroxyproline is a major component (Szpak, 2011). Hydroxyproline is a reflection of liver collagen synthesis during liver fibrosis (Ala-Kokko et al., 1987). Hepatic egg granulomas contains the majority of proline incorporated into hydroxyproline (Olds et al., 1985). It is suggested that there are factors in S. mansoni egg granulomas which elevate the free L-proline content in the fibrotic liver (Tanabe et al., 1991). Inhibition of the deposition of extracellular matrix proteins is the aim of any antifibrotic therapy (Bataller and Brenner, 2005). Early administration of antischistosomal drugs might lead to resorption of collagen, while late treatment at chronic stages leads only to little diminution in fibrosis (Mehlhorn et al., 1982).
Artemether is already being widely used against malaria (McIntosh and Olliaro, 2001) has been shown to have antischistosomal properties against all human Schistosome species (Utzinger et al., 2002, 2003). Its effect is directed towards reduction of adult worms especially females (Xiao et al., 2000, 2001). It reduces number of excreted ova (Mahmoud et al., 2006) with increase number of dead ova (Seif el-Din et al., 2011; Sayed El-Ahl et al., 2013). Significant reduction in eggs prevents granuloma formation with subsequent morbidity in liver (Botros et al., 2010). ART anti-pathologic activities is apparent especially in early treatment after infection (Hamza et al., 2012). Even granulomata after ART treatment show lower collagen content than control (Mahmoud et al., 2006).
Somatostatin is a peptide hormone with regulatory effect on the endocrine system. It affects neurotransmission and cell proliferation. Somatostatin inhibits the release of numerous secondary hormones through interacting with G-protein-coupled somatostatin receptors (Chatterjee and Van Marck, 2001). Clinically, Somatostatin, is reported to decrease portal pressure, control variceal bleeding and reduce hepatic fibrosis (Fevery and Raptis, 1997). In schistosomiasis that cause clinical morbidity in the rodent model SOM has been associated with significant decrease in liver ova count and a significant increase in dead ova (Mansy et al., 1998). Also significant decreased size of cellular and fibrocellular granulomas (Mansy et al., 1998). SOM administration has its direct morphological morbid effect on the adult worm of Schistosoma mansoni (Sayed El-Ahl et al., 2013). Significant reduction of hepatic hydroxyproline levels is accompanied by treatment of acute and chronic infected animals with somatostatin (Chatterjee et al., 2005). SOM can immunomodulate schistosomiasis-induced inflammatory responses in the liver and intestines and so it has a direct effects on schistosomiasis-caused morbidity (Mansy et al., 1998). It has possible effects on the Schistosoma mansoni parasite stages (Sayed El-Ahl et al., 2013).
Based on these reports, the hypothesis that combined treatment of ART and SOM could potentiate alleviation of hepatic fibrosis caused by schistosomiasis is presented. To prove this ART and SOM are administrated as combined and single therapies for Schistosoma mansoni infected mice. Heptic hydroxyproline as a marker of collagen deposition in infected mice was monitored compared to uninfected or untreated groups.
MATERIALS AND METHODS
Experimental animals and infection: Laboratory bred male Swiss albino mice weighing about 20-25 gm and Schistosoma mansoni cercariae (Egyptian strain) were obtained from Schistosoma Biological Supply Program (SBSP) Unit at Theodor Bilharz Research Institute (TBRI), Giza, Egypt. Cercariae were used immediately after shedding from Biomphalaria alexandrina snails. Cercarial count was done according to Moore et al. (1977). Infection was done in a dose of 60±10 cercariae/mouse by body immersion technique according to Staden (1949). The experiment started with 200 mice but actually carried out on 90 survived mice.
Mice grouping: Group I (18 mice): Infected non treated control group. Group II (18 mice): non Infected non treated control group. Group III (54 mice): Infected group subdivided into three subgroups each of 18 mice: 1st subgroup: treated at 2 weeks PI and sacrificed at 5 weeks, 2nd subgroup treated at 5 weeks PI and sacrificed at 8 weeks, 3rd subgroup: treated at 13 weeks PI and sacrificed at 16 weeks. The infected treated mice were equally divided between three regimens of treatment: SOM alone, ART alone and combined SOM plus ART.
Drug regimen: SOM was administered as Octreotid (SOM analogue) in a dose of 0.006 mg subcutaneously in 2 equal divided doses daily for two weeks, ART as single dose of 300 mg kg-1 intramuscularly and combined treatment with both SOM and ART by the same mentioned doses. Octreotid is a somatostatin analog having the same pharmacological actions as the native SOM. It was used due to its higher specificity, better potency, longer duration of action and different routes of administration (Lemaire et al., 1989). It was supplied as watery ampoules each contain 0.1 mg mL-1.
Sacrificing mice was done by decapitation without anesthesia. Each mouse of the studied groups was processed for assessment of the following: tissue bounding ova (oogram) pattern was done to determine the percentage of immature, mature and dead S. mansoni eggs according to Pellegrino et al. (1962), ova count per gram tissue (liver and intestine) according to Cheever (1968).
Determination of hepatic content of hydroxyproline: One gm of liver tissue was taken and total liver collagen was determined as hydroxyproline (Trans-4 hydroxy-L-proline) according to the method of Woessner (1961).
Statistical analysis: Results were collected, tabulated, statistically analyzed using one-way analysis of Variance (ANOVA) according to Campbell (1989). Comparison between two groups was done by the Student’s t-test. The p-value of 0.05 or less was taken to signify statistical significance.
RESULTS AND DISCUSSION
On reviewing the literature, trial of combined regimen of ART and SOM administration in treatment of schistosomiasis is the first one to general knowledge. In the present study, combined treatment group attained the lowest significant mean hepatic hydroxyproline content over ART and SOM single therapy groups. At 2,5 and 13 weeks PI it was 793.41±64.91, 1010.87±75.67 and 1021.42±135.60 in comparison to normal control (660.30±51.48, 660.30±51.48, 660.30±51.48) and the infected control (1209.88±80.12, 1436.57±89.11, 1696.21±90.41), respectively (Table 1). This significant low hepatic hydroxyproline content in treated group could be correlated with the significant low mean egg load in the same group (Table 2) in the corresponding periods. It was 2.22±1.13, 2.47±0.37, 11.40±1.42 in comparison to infected control (11.32±1.41, 10.39±0.53, 17.01±1.68). At 2 weeks PI (Table 2) there was complete absence of immature and mature ova (0±0) and higher significant mean level of dead ova (100±0) in comparison to infected control (40.33±2.42, 47.67±2.29, 12.00±1.36). The other two periods (5 and 13 weeks PI) also showed highest mean dead ova (91.71±3.91, 84.78±3.21) in comparison to control (11.00±1.57, 29.25±6.35).
| Table 1: | Hepatic Hydroxyproline content and egg load in mice liver following drug treatment at 2, 5 and 13 weeks post infection with S. mansoni cercariae and sacrificed at 5, 8 and 16 weeks post infection respectively. Values given are Means±SE |
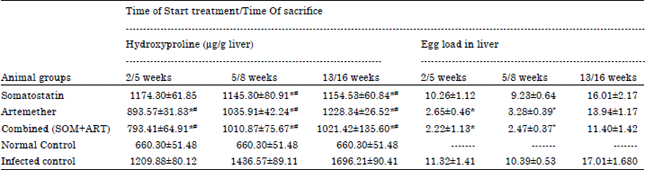 | |
| SOM: Somatostatin, ART: Artemether, PI: Post infection. *Significant difference from infected control at p<0.05. #Significant difference from normal control at p<0.05 | |
| Table 2: | Oogram pattern in liver and intestine following drug treatment at 2, 5 and 13 weeks post infection with S. mansoni cercariae and sacrificed at 5, 8 and 16 weeks post infection respectively. Values given are Means±SE |
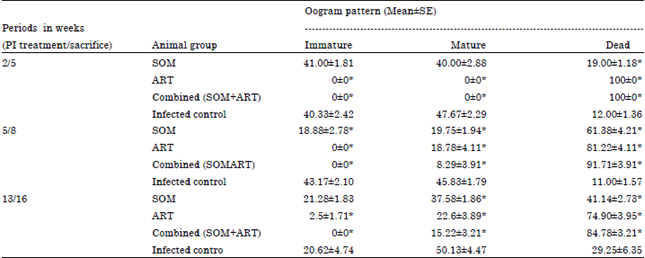 | |
| SOM: Somatostatin, ART: Artemether, PI: Post infection . *Significant difference from infected control at p<0.05 | |
The present results coincide with Pellegrino and Katz (1969), Warren and Boros (1975), Cheever et al. (1992) and Yue et al. (1984) where granulomatous response and subsequently hydroxyproline level is affected by reduction on egg load and increased number of dead ova.
In the present study, mice group under ART single administration recorded nearby results of combined treatment group. ART single therapy group followed combined treatment group in attaining lower significant mean Hydroxyproline level in the 2, 5 and 13 weeks period PI (893.57±31.83, 1035.91±42.24, 1228.34±26.52) than normal control and infected control. This also could be correlated with significant lower mean egg load by ART therapy (Table 2) in the corresponding 3 periods (2.65±0.46, 3.28±0.39. 13.94±1.17) in comparison to control. As combined therapy at 2 weeks PI ART single therapy group showed complete absence of immature and mature ova (0±0) and higher significant mean level of dead ova (100±0) in comparison to infected control (Table 2). The other two periods (5 and 13 weeks PI) also showed higher mean dead ova (81.22±4.11, 74.90±3.95) in comparison to control group (11.00±1.57, 29.25±6.35) which was reflected on the hydroxyproline level. In agreement of the present study is Mahmoud et al. (2006) where complete absence of all egg developmental stages and the high reduction of egg load in treated mice with ART (200 mg k-1) at 5,6,7 weeks PI (95.5%) are detected. There are also minimal number of portal cellular granuloma with lower diameter and moderate collagen content in comparison to control. Dead ova have no granulomatous reaction around (Pellegrino and Katz, 1969; Warren and Boros, 1975; Cheever et al., 1992). There is also a positive correlation between the severity of hepatic fibrosis and the number of eggs in the tissue (Yue et al., 1984). ART reduces no of excreted ova (Mahmoud et al., 2006) with increase no of dead ova (Seif el-Din et al., 2011; Sayed El-Ahl et al., 2013). Significant reduction in eggs prevents granuloma formation with subsequent morbidity in liver (Botros et al., 2010). Anti-pathologic activities of ART is apparent especially in early treatment after infection (Hamza et al., 2012). Even granulomata after ART treatment shows lower collagen content than control (Mahmoud et al., 2006). In the present study, at 13 weeks PI none of combined therapy or single therapy with ART showed lower mean egg load this could be attributed to the late period of ART administration to infected mice groups. Early use of ART on schistosomiasis infection is more effective (Mahmoud et al., 2006) as juvenile stages of schistosomes are more susceptible to artemether than adult worms (Xiao et al., 2000; Hamza et al., 2012). Diminution or even no egg granulomas in liver sections of mice treated with artemether, at early administration after infection could be detected (Mahmoud and Botros, 2005; Botros et al., 2010).
In the present study (Table 2), SOM treated group attained non-significant lower mean egg load (10.26±1.12, 9.23±0.64, 16.01±2.1) at the three periods in comparison to infected control group. However, SOM followed ART therapy in attaining significant lower mean hydroxyproline content in 5 and 13 weeks PI (1145.30±80.91, 1154.53±60.84), respectively in comparison to infected control group. This could be attributed to the significant higher dead ova after SOM administration in 2, 5 and 13 weeks PI (19.00±1.18, 61.38±4.21, 41.14±2.73) in comparison to control. There was also significant reduction of mean number of mature ova 5 and 13 weeks PI periods (19.75±1.94, 37.58±1.86) in comparison to control group. So the decreased liver ova count and the increased dead eggs detected in these subgroups may participate in part in decreasing hepatic fibrogenesis which is reflected on hydroxyproline level. These findings are supported by Chatterjee et al. (2005) where treatment of acute and chronic infected animals with somatostatin significantly reduces hepatic hydroxyproline levels. Associated significant decrease in liver ova count and a significant increase in dead ova after SOM treatment has been domcumented by Mansy et al. (1998). Lower mean hydroxyproline of the present study coincide with significant decreased size of cellular and fibrocellular granulomas after SOM administration (Mansy et al., 1998). In schistosomiasis the reaction to Schistosoma eggs can be minimized by stopping ova production by living worms, killing of female worms or by suppressing immunological reactions around S. mansoni eggs (Mahmoud and Warren, 1974). Somatostatin has strong affinity for the SOM receptor subtypes 2, 3 and 5 and can interact with them on S. mansoni egg and worm stages preventing the pathological morbidity of the parasite in the body (Chatterjee et al., 2005). Sayed El-Ahl et al. (2013) reveals that SOM therapy has its direct morphological morbid effect on the adult worm of Schistosoma mansoni including male and female worms in experimentally infected mice. An observation of the present study is that hepatic hydroxyproline content (Table 1) reached its lowest level at 5 weeks PI in SOM treated group. This is in agreement with Chatterjee et al. (2005). Somatostatin therapy is effective after two days of treatment with no further reduction in pathology after five days of therapy (Chatterjee et al., 2007). It is worthy to note that exogenous administration of SOM in experimental animals could alleviate the pathology caused by schistosomiasis through reducing either the number of parasite eggs or the secretion of fibrosis inducing-mediators (Chatterjee et al., 2005). There is also direct association between S. mansoni induced fibrosis and low endogenous SOM levels in human subjects (Chatterjee et al., 2004). Octreotid as SOM analogue, has been shown to modulate fibroblast activation in vitro (Priestley et al., 1994; Pasquali et al., 2000). It exerts antifibrotic properties in hepatic, esophageal, or digestive fibrosis of rat models of (Fort et al., 1998; Wang et al., 2001; Turkcapar et al., 2003) and decreases the accumulation of connective tissue in mice (Mansy et al., 1998). Some data also indicate that octreotid inhibits the in vivo expression of transforming growth factor-b, a key fibrogenic mediator in rat models of intestinal and peritoneal fibrosis (Gunal et al., 2001). SOM is shown to significantly reduce growth hormone secretion which can lead to a decline in circulating insulin-like growth factor-1 (IGF-1). In turn this may affect collagen production as insulin-like growth factor and growth hormone stimulate growth and collagen deposition (Patel, 1990; Allen and Goldberg, 1992; Cotran et al., 1994). The main feature of the presented data is the nearby results of ART single administration to that of combined treatment while that of SOM is somewhat away even the significant results.
CONCLUSION
All over the study periods combined therapy showed the upper hand in lowering hydroxyproline in liver of infected mince. Mostly the combined treatment was followed by ART single therapy with nearby results then SOM single therapy. So, it is concluded that the use of SOM plus ART in alleviating hepatic fibrosis in experimental infection of Schistosomia mansoni is not powerfully recommended since the major effect is always referred to ART alone.
REFERENCES
- Ala-Kokko, L., F. Stenback and L. Ryhanen, 1987. Preventive effect of malotilate on carbon tetrachloride-induced liver damage and collagen accumulation in the rat. Biochem. J., 246: 503-509.
Direct Link - Allen, D.B. and B.D. Goldberg, 1992. Stimulation of collagen synthesis and linear growth by growth hormone in glucocorticoid treated children. Pediatrics, 89: 416-421.
Direct Link - Alves Oliveir, L.F., E.C. Moreno, G. Gazzinelli, O.A. Martins filho and A.M. Silveira et al., 2006. Cytokine production associated with periportal fibrosis during chronic schistosomiasis mansoni in humans. Infect. Immun., 74: 1215-1221.
CrossRefDirect Link - Bataller, R. and D.A. Brenner, 2005. Liver fibrosis. J. Clin. Invent., 115: 209-218.
CrossRefPubMedDirect Link - Botros, S.S., O. Hammam, M. Mahmoud and R. Bergquist, 2010. Praziquantel efficacy in mice infected with PZQ non-susceptible S. mansoni isolate treated with artemether: Parasitological, biochemical and immunohistochemical assessment. APMIS, 118: 692-702.
CrossRefDirect Link - Chatterjee, S., J.O. De Beeck, A.V. Rao, D.V. Desai and G. Vrolix et al., 2007. Prolonged somatostatin therapy may cause down-regulation of SSTR-like GPCRs on Schistosoma mansoni. J. Vect. Borne. Dis., 44: 164-180.
PubMedDirect Link - Chatterjee, S. and E. Van Marck, 2001. The role of somatostatin in schistosomiasis: A basis for immunomodulation in host-parasite interactions? Trop. Med. Int. Health, 6: 578-581.
CrossRefDirect Link - Chatterjee, S., A. Mbaye, A.T. Alfidja, J. Weyler and J.T. Scott et al., 2004. Circulating levels of the neuropeptide hormone somatostatin may determine hepatic fibrosis in Schistosoma mansoni infections. Acta Tropica, 90: 191-203.
CrossRefDirect Link - Chatterjee, S., G. Vrolix, I. Depoortere, T. Peeters and E. Van Marck, 2005. The therapeutic effect of the neuropeptide hormone somatostatin on Schistosoma mansoni caused liver fibrosis. BMC Infect Dis., Vol. 5.
CrossRef - Cheever, A.W., 1968. Condition affecting the accuracy of potassium hydroxide digestion techniques for counting Schistosoma mansoni eggs in tissues. Bull. World Health Org., 39: 328-331.
PubMed - Cheever, A.W., J.G. Maccedoni, S. Deb, E.A. Cheever and J.E. Mosimann, 1992. Persistence of eggs and hepatic fibrosis after treatment of infected mice. J. Trop. Med. Hyg., 46: 752-758.
PubMedDirect Link - Dunn, M.A., M. Rojkind, K.S. Warren, P.K. Hait, L. Rifas and S. Seifter, 1977. Liver collagen synthesis in murine schistosomiasis. J. Clin. Invest., 59: 666-674.
CrossRefDirect Link - Fort, J., F. Oberti, C. Pilette, N. Veal and Y. Gallois et al., 1998. Antifibrotic and hemodynamic effects of the early and chronic administration of octreotide in two models of liver fibrosis in rats. Hepatology, 28: 1525-1531.
CrossRefDirect Link - Gunal, A.I., S. Duman, S. Sen, A. Unsal, E. Terzioglu, F. Akcicek and A. Basci, 2001. By reducing TGF beta 1, octreotid lessens the peritoneal derangements induced by a high glucose solution. J. Nephrol., 14: 184-189.
PubMedDirect Link - Hamza, R.S., A.S. Metwaly and D.A. Abo El-Maaty, 2012. Effects of artemether treatment on prepatent and patent schistosoma mansoni infection in experimentally infected mice. PUJ, 5: 147-154.
Direct Link - Lemaire, M., M. Azira, R. Dannecker, P. Marbach, A. Schweitzer and G. Maurer, 1989. Disposition of Sandostatin, a new synthetic somatostatin analog, in rats. Drug. Metab. Dispos., 17: 699-703.
Direct Link - Mahmoud, M., F. Ebeid and M. Nosseir, 2006. Enhanced role of grapefruit juice on the antischistosomal activity of artemether on the liver of Schistosoma haematobium infected hamsters. Sci. Pharm., 74: 59-75.
Direct Link - Mahmoud, M.R. and S.S. Botros, 2005. Artemether as adjuvant therapy to praziquantel in murine Egyptian Schistosomiasis mansoni. J. Parasitol., 91: 175-178.
CrossRefDirect Link - Mansy, S.S., H.A. Yehia, M.M. Hassan, E.A. Hassan, M.M. Youssef, A.A. Hadi and C.D. Mackenzie, 1998. Effect of octreotide on the pathology of hepatic schistosomiasis. Arzneimittelforschung, 48: 855-861.
PubMed - Mehlhorn, H., J.K. Frenkle, P. Andrews and H. Thomas, 1982. Light and electron microscope studies on Schistosoma mansoni granulomas of mouse liver following treatment with praziquantel. Treponmed. Parasit., 33: 229-239.
PubMed - Moore, D.L., D.I. Grove and K.S. Warren, 1977. The Schistosoma mansoni egg granuloma: Quantitation of cell populations. J. Pathol., 121: 41-50.
PubMedDirect Link - Olds, G.R., A. Griffin and T.F. Kresina, 1985. Dynamics of collagen accumulation and polymorphism in murine Schistosoma japonicum. Gastroenterology, 89: 617-624.
PubMed - Pasquali, D., P. Vassallo, D. Esposito, G. Bonavolonta, A. Bellastella and A.A. Sinisi, 2000. Somatostatin receptor gene expression and inhibitory effects of octreotide on primary cultures of orbital fibroblasts from Graves ophthalmopathy. J. Mol. Endocrinol., 25: 63-71.
PubMed - Szpak, P., 2011. Fish bone chemistry and ultrastructure: Implications for taphonomy and stable isotope analysis. J. Archaeol. Sci., 38: 3358-3372.
CrossRefDirect Link - Pellegrino, J. and N. Katz, 1969. Laboratory evaluation of antischistosomal agents. Ann. N. Y. Acad. Sci., 160: 429-460.
CrossRefDirect Link - Pellegrino, J., C.A. Oliveira, J. Faria and A.S. Cunha, 1962. New approach to the screening of drugs in experimental schistosomiasis mansoni in mice. Am. J. Trop. Med. Hyg., 11: 201-215.
CrossRefPubMedDirect Link - Priestley, G.C., R.D. Aldridge, P.J. Sime and D. Wilson, 1994. Skin fibroblast activity in pretibial myxoedema and the effect of octreotid (Sandostatin) in vitro. Br. J. Dermatol., 131: 52-56.
PubMed - Sayed El-Ahl, S.A., M.A. Hegazi, M. Mahmoud, F. El-Zahraa, M. Awadallah and M.A. Abd Rabo, 2013. Parasitological changes within experimentally murine schistosomiasis mansoni upon treatment by somatostatin, artemether and their combination. Res. J. Parasitol., 8: 1-13.
CrossRefDirect Link - Seif el-Din, S.H., A.M. Al-Hroob and F.A. Ebeid, 2011. Schistosoma mansoni: N-acetylcysteine downregulates oxidative stress and enhances the antischistosomal activity of artemether in mice. Exp. Parasitol., 128: 230-235.
CrossRefPubMedDirect Link - Staden, O.D., 1949. Experimental schistosomiasis: Maintenance of schistosoma mansoni in the laboratory with some notes on experimental infection with S. haematobium. Ann. Trop. Med. Parasit., 43: 268-283.
PubMed - Tanabe, M., N. Kaneko and T. Takeuchi, 1991. Schistosoma mansoni: Higher free proline levels in the livers of infected mice. Exp. Parasitol., 72: 134-144.
PubMed - Turkcapar, N., S. Bayar, A. Koyuncu and K. Ceyhan, 2003. Octreotid inhibits hepatic fibrosis, bile duct proliferation and bacterial translocation in obstructive jaundice. Hepatogastroenterology, 50: 680-683.
PubMed - Utzinger, J., J. Chollet, Z. Tu, S. Xiao and M. Tanner, 2002. Comparative study of the effects of artemether and artesunate on juvenile and adult Schistosoma mansoni in experimentally infected mice. Trans. R. Soc. Trop. Med. Hyg., 96: 318-323.
PubMed - Utzinger, J., J. Keiser, X. Shuhua, M. Tanner and B.H. Singer, 2003. Combination chemotherapy of schistosomiasis in laboratory studies and clinical trials. Antimicrob. Agents Chemother., 47: 1487-1495.
CrossRef - Wang, J., H. Zheng and M. Hauer-Jensen, 2001. Influence of short-term octreotid administration on chronic tissue injury, transforming growth factor beta (TGFbeta) overexpression and collagen accumulation in irradiated rat intestine. J. Pharmacol. Exp. Ther., 297: 35-42.
PubMed - Woessner Jr. J.F., 1961. The determination of hydroxyproline in tissue and protein samples. Arch. Biochem. Biophys., 93: 440-447.
PubMed - Xiao, S.H., M. Booth and M. Tanner, 2000. The prophylactic effects of artemether against Schistosoma japonicum infections. Trends Parasitol., 16: 122-126.
CrossRef - Xiao, S.H., J. Chollet, J. Utzinger, H Matile, J.Y. Mel and M. Tanner, 2001. Artemether administered together with haemin damages shcistosmes in vitro. Trans. Royal Soc. Trop Med. Hyg., 95: 67-71.
PubMed