
Magda M. Sanad
Department of Zoology, College of Sciences, Female Center for Scientific and Medical College,
King Saud University, P.O. Box 22452, Riyadh 11495, Saudi Arabia
Ebtesam M. AL Olayan
Department of Zoology, College of Sciences, Female Center for Scientific and Medical College,
King Saud University, P.O. Box 22452, Riyadh 11495, Saudi Arabia
Research Journal of Parasitology
Year: 2011 | Volume: 6 | Issue: 2 | Page No.: 60-73







ABSTRACT
Toxoplasma gondii has been considered as one of the most successful models for intracellular parasitism. Throughout the last two decades, considerable progress has been made to clarify the obscurity of both pathogenicity and long-lasting chronicity of toxoplasmosis. The recent integration of the many sources of genomic, proteomic and metabolomic data has allowed good clarification of the molecular details of the parasite-dependent effects for Toxoplasma-infected cells. Mechanisms of host-cell invasion, intracellular survival, stage-conversion of the slow-growing bradyzoite within long-lasting tissue cysts to a rapidly replicating highly pathogenic tachyzoite and vice versa, structure and composition of the apical organelles, subversion of inflammatory host response by the parasite, interference with effector functions of immune cells, evasion of host immune defenses, modulation of host cell apoptosis and the recent achievements in the use of lipid synthetic pathways by the parasite etc., all have been recently highlighted but still in need for further elucidation. This review offers a brief declaration of the progress that has been made to understand the curious intimate relationship between Toxoplasma gondii and its host cell.
PDF Abstract XML References Citation
Received: February 07, 2011;
Accepted: March 01, 2011;
Published: December 02, 2011
How to cite this article
Magda M. Sanad and Ebtesam M. AL Olayan, 2011. Toxoplasma gondii Strategy for Intracellular Survival: Is it Still Enigmatic?. Research Journal of Parasitology, 6: 60-73.
URL: https://scialert.net/abstract/?doi=jp.2011.60.73
URL: https://scialert.net/abstract/?doi=jp.2011.60.73
INTRODUCTION
Successful parasites are able to use complex strategies to seclude from hostile conditions in their environment. Some parasites arise a fascinating behavior by using cytokines of host origin induced by the infectious process for their own development, others gain the success of genetic restriction of the immune response or able to invade immunologically competent macrophages and to avoid the triggering of killing mechanisms of these cells (Camus et al., 1995). Concealment of specific antigens from the immune attack is an additional strategy. In a previous work (Azab and Sanad,1993; Sanad and Emara, 1994), localization of secreted antigens of Trichinella spiralis infective larva have been detected in the cytoplasm and in hypertrophied nuclei of the parasitized muscle fiber and proved to have a role in the formation and maintenance of the nurse cell, the strategy by which the parasite uses the host cell for its long-term survival.
Toxoplasma gondii, a ubiquitous apicomplexan parasite of humans and other worm-blooded animals, has been considered as the cause of the most prevalent parasitic zoonosis (Sanad and Al-Ghabban, 2007; Sabry and Reda, 2008; Asgari et al., 2009) and is the causal agent of significant morbidity and mortality among humans worldwide (Blader et al., 2001; Nissapatorn, 2007; Nissapatorn et al., 2007). One of toxoplasmosis mysteries is how, in immunocompetent patients, a balanced host-parasite relationship is usually established and infections are often benign and chronic and how in immunocompromised patients, the balance of such chronic infection can readily be disrupted leading to an overt disease. The mechanisms of stage-conversion of the slow-growing bradyzoite within long-lasting tissue cysts to a rapidly replicating highly pathogenic tachyzoite and vice versa, still poorly understood.
Genome sequencing efforts and the broad availability of DNA microarrays that cover the genomes of several major host species have allowed high-throughput approaches for the analyses of host cell modifications by Toxoplasma (Blader et al., 2001; Saeij et al., 2007). Furthermore, significant technical improvements in proteome analyses have recently enabled investigations into how intracellular T. gondii manipulates its host cell at a post-transcriptional level (Nelson et al., 2008).
This review explains how the recent advent of genetic tools applicability for use in T. gondii creates the hope for convenient clarification of curiosity of both pathogenicity and long-lasting chronicity of toxoplasmosis.
Structure, life cycle and transmission of Toxoplasma gondii (Fig. 1-4): Toxoplasma gondii has three infectious stages: the tachyzoites (in groups), the bradyzoites (in tissue cysts) and the sporozoites (in oocysts).
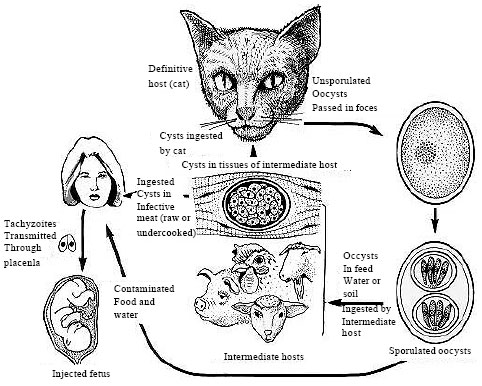 | |
| Fig. 1: | Life cycle of Toxoplasma gondii (From Dubey JP, Toxoplasmosis. J Am Vet Med Assoc 189:166-170, 1986) Cats, the definitive hosts of T gondii, can become infected by ingesting sporulated oocysts or (most often) infected animals. The oocysts are infectious to most mammals and birds. Toxoplasma can be transmitted to intermediate hosts through oocysts, by carnivorism, or transplacentally. Transplacental transmission is most important in humans and sheep |
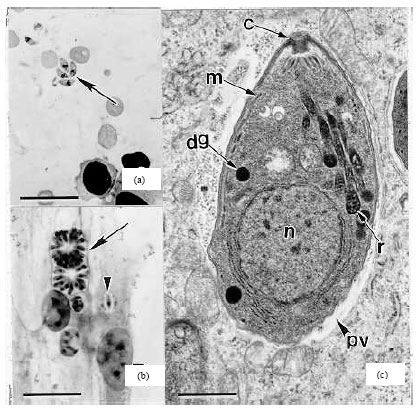 | |
| Fig. 2: | Tachyzoites of T. gondii (By D.S. Lindsay, Auburn University, Auburn, AL. Quoted from Baron Samuel Medical Microbiology, 4th edition, University of Texas Medical Branch at Galveston, 1996, Chapter 84 Toxoplasma gondii). (a) Extracellular (arrow) released from host cells. Impression smear, Giemsa stain. Bar = 20 μm. (b) Intracellular in cell culture. A group arranged in a rosette (arrow) and vacuole (arrowhead) around a tachyzoite. Immunohistochemical stain with a tachyzoite-specific monoclonal antibody. Bar = 20 μm. (c)Transmission electron micrograph of an intracellular tachyzoite. A Parasitophorous Vacuole (PV) around the tachyzoite. Parasite organelles visible in this picture include a conoid (c), micronemes (m), dense granules (dg) nucleus (n) and rhoptries (r). Bar = 0.8 μm |
Once tissue cysts or the environmental oocysts are ingested, their contents i.e .,the bradyzoites and sporozoites respectively, invade host cells and differentiate into tachyzoites. The tachyzoite has a pellicle, a poler ring, a conoid, rhoptries, micronemes, mitochondria, subpellicular microtubules, endoplasmic reticulum, a Golgi apparatus, ribosomes, rough surfaced endoplasmic reticulum, a micropole and a well defined nucleus. Following active penetration of the host cell membrane, the tachyzoite becomes surrounded by a Parasitophorous Vacuole (PV) with numerous intravacuolar tubules connecting the parasitophorous vacuolar membrane to the parasite pellicle (Dubey and Beattie, 1988). The tachyzoite multiplies asexually by repeated endodyogeny within many 6different types of the host cell. After a few divisions, tachyzoites differentiate into latent bradyzoite forms which are surrounded by a carbohydrate-rich cyst wall within the parasitophorous vacuole. The development of these tissue cysts are more prevalent in the neural and muscular tissues, such as the brain, eye, skeletal and cardiac muscle. Within the tissue cyst, bradyzoites multiply slowly by endodyogeny.
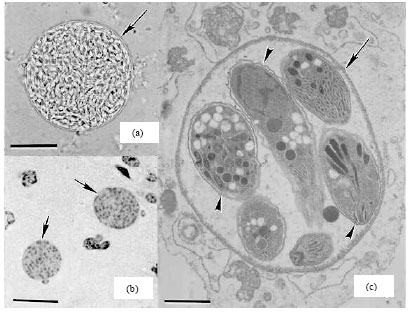 | |
| Fig. 3: | Tissue cysts of T. gondii. (By D.S. Lindsay, Auburn University, Auburn, AL. Quoted from Baron Samuel Medical Microbiology, 4th edition, University of Texas Medical Branch at Galveston, 1996, Chapter 84 Toxoplasma gondii). (a) Tissue cyst freed from mouse brain. A thin (arrow) cyst wall enclosing hundreds of bradyzoites. Unstained. Bar = 20 μm. (b) Two tissue cysts (arrows) in section of brain. Hematoxylin and eosin stain. Bar = 20 μm. (c) Transmission electron micrograph of a small tissue cyst in cell culture. Thin cyst wall (arrow) enclosing 6 bradyzoites (arrowheads). Bar = 1.0 μm |
Tissue cysts are the terminal life-cycle stage in the intermediate host and intact tissue cysts can persist for the life of the immunocompetent host without causing any harm (Tenter et al., 2000).
Cats, the sole definitive host, shed oocysts after ingesting any of the three infectious stages of T. gondii. After the ingestion of tissue cysts, the released bradyzoites initiate development of numerous asexual enteroepithelial generations of T. gondii that divide by endodyogeny, endopolygeny or schizogony before gametogony occurs (Dubey and Frenkel, 1972). The fertilized gamete becomes surrounded by oocyst wall and unsporulated oocysts are shed to the external environment where sporulation takes place within 1-4 days and the oocysts remain infective for months in shaded moist soil (Dubey et al., 1970).
Toxoplasma gondii is unusual in that its propagation does not require passage through the definitive host . In humans, infection is commonly acquired by oral ingestion of undercooked infected meat used as a food, including sheep, pigs and rabbits (Cook et al., 2000; Dubey, 2008) or by consumption of water or food products that are contaminated with oocysts (Dubey and Jones, 2008). Vertical transmission of the rapidly growing tachyzoite occurs if an immunologically naïve mother acquires a new infection during pregnancy. Less frequently, human infection can occur directly through blood transfusion and organ transplants from an infected person (Feigin and Cherry, 1998), by sexual means and by ingestion of unboiled goat's milk or raw hen's eggs (Dubey, 1998). Hay, straw and grain that are contaminated with cat faeces have been identified as sources of infection for livestock (Chinchilla et al., 1994).
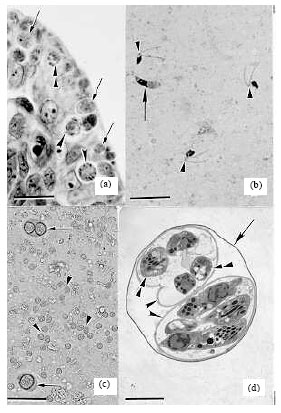 | |
| Fig. 4: | Sexual stages of T gondii (By D.S. Lindsay, Auburn University, Auburn, AL. Quoted from Baron Samuel Medical Microbiology, 4th edition, University of Texas Medical Branch at Galveston, 1996, Chapter 84, Toxoplasma gondii). (a) Schizonts (double arrowheads), female gamonts (arrows) and male gamonts (arrowheads) in section of superficial epithelial cells of the small intestine of a cats. Hematoxylin and eosin stain. Bar = 15 μm. (b) Three male gametes each with 2 flagella (arrowheads) compared with a merozoite (arrow). Impression of intestinal epithelium of a cat. Giemsa stain. Bar = 10 μm. (c) Unsporulated oocytes (arrowheads) in feces of a cat. Two oocysts of another feline coccidium, Isospora felis (arrowheads). Isospora felis sporulates faster than T gondii. The oocysts on top of the picture already contains 2 sporocysts while all T gondii oocysts are unsporulated. Unstained. Bar = 65 μm. (d) Transmission electron micrograph of a sporulated oocyst. Thin oocyst wall (arrow), 2 sporocysts (arrowheads) and 4 sporozoites (double arrowheads) in sporocysts. Bar = 2.25 μm |
Population structure of Toxoplasma gondii: Toxoplasma has a remarkably clonal population structure. However, there have been some notable surprises about its population biology. First is that a few strains have come to dominate in some very broad niches (Sibley and Boothroyd, 1992). Within Europe and North America, most strains isolated from humans and domestic/farm animals are one of just three clonal types I, II and III (Howe and Sibley, 1995). Equally surprising, these three strains are closely related with types I and III appearing to be the products of a cross between type II and one or other of two strains, alpha and beta (Saeij et al., 2005). In other regions of the world, types I, II and III do not predominate. Strains from America may have the greatest ecological diversity (Lehmann et al., 2006). A tempting scenario for today’s situation, as described by Boothroyd (2009) is that the enormous ecological disruption that accompanied European colonization of the Americas and Africa made possible matings between previously isolated strains of Toxoplasma (e.g., type II, alpha and beta).
Types I, II and III strains differ in many interesting and important biological properties, not least of which is the course of the infections they produce in mice. The genetic basis of these phenotypes can be mapped in F1 progeny of crosses between types I and III or II and III (Saeij et al., 2006).
In addition to the gross biological phenotypes of disease severity, in vitro methods can be used to look for molecular differences in how a host cell responds to the invaded parasite. Microarray technology has given the field a way to interrogate the entire transcriptome of the host cell to determine which host genes might respond differently to different strains of Toxoplasma (Kim et al., 2004). The results have revealed a startling number of strain-specific differences in host response (Saeij et al., 2007). Mapping these differences back onto the parasite genome has been done and one polymorphic parasite gene, ROP16, has so far been found to be a crucial driver of differences in the host transcriptional response (Boothroyd, 2009).
Correlation between Toxoplasma gondii virulence and growth rate: The T. gondii parasite burden is a major contributor to Toxoplasma pathogenesis in mice. It causes overstimulation of the immune system with a resultant high levels of T helper cell type 1 (Th1) cytokines, increased apoptosis and organ damage (Nguyen et al., 2003). Although one tachyzoite of a type I strain is sufficient to generate high parasite loads and high levels of Th1 cytokines, high parasite loads of a type II strain generated by increased inoculation, will lead to equally high levels of cytokines and pathology (Saeij et al., 2005). Different pathology and repeated acute toxoplasmosis have been suggested to be due to difference in strains virulence (Abdul-Fattah et al., 1992).
Intracellular parasitism and role of organelles: Multiplication of Toxoplasma gondii only occurs intracellularly inside a specialized compartment called the Parasitophorous Vacuole (PV). Regulated secretory processes are key to the success of the intracellular parasitism of Toxoplasma as the parasite extensively modifies the newly formed vacuole using secreted proteins (Beckers et al., 1994). The main structural modification of the PV consists of elaboration of a network of tubular membranes that are continuous with the vacuolar membrane (Sibley et al., 1986).
Toxoplasma cells contain three distinct secretory organelles: the rhoptries, the micronemes and the dense granules which are specialized for the invasion and remodeling of the host. The micronemes secrete a collection of adhesion proteins termed MICs that mediate host cell entry (Cerede et al., 2005). Rhoptries, which are discharged immediately following micronemes, are comprised of two different substructures named the rhoptry neck and rhoptry bulb. During invasion, rhoptry proteins and lipids, with the assistance of microneme protein AMA1 form the moving junction (Alexander et al., 2005), a tight apposition of the parasite and host plasma membranes visible during cell invasion. The moving junction ensures the formation of the Parasitophorous Vacuole Membrane (PVM) from the host cell membrane, that enables efficient procurement of nutrients and evasion of host immune defenses (Boothroyd and Dubremetz, 2008). Proteomic analysis of the rhoptries has resulted in the identification of rhoptry neck proteins (RONs) in addition to traditional rhoptry proteins (ROPs) (Bradley et al., 2005). These RONs and ROPs are involved in different aspects of formation of the moving junction during invasion (Alexander et al., 2005). ROPs are also injected into the host cell resulting in extensive modification of host gene expression and signaling pathways (Bradley and Sibley, 2007).
Shortly after invasion, the association between the PV and host organelles such as mitochondria and Endoplasmic Reticulum (ER) is seen (Magno et al., 2005). One parasite secretory protein, ROP2, directly anchors host mitochondria to the PV membrane (Sinai and Joiner, 2001). Presumably, other rhoptry proteins are responsible for anchoring host ER to the PV membrane. Finally, dense granule proteins appear to facilitate formation of specialized tubules that enable nutrient acquisition by the parasite and some of these proteins, most notably GRA7 are also secreted into host cells (Coppens et al., 2006).
Host cell invasion and egress by Toxoplasma gondii: Both host cell invasion and egress by Toxoplasma gondii are essential for infection and replication and involve morphological changes, motility and secretion from various organelles. Success of intracellular survival depends on the parasite ability to propagate from one cell to another and thus invasion and egress are highly and precisely regulated events. The intracellular growth of an apicomplexan parasite eventually causes lethal lysis of the host cells in a mechanism termed egress, which results in the exit of infective parasites from their parasitophorus vacuoles. In T. gondii, host cell lysis is a less critical issue in comparison to other apicomplexan parasites since endodyogenic division allows production of infective parasites at the end of every round of replication. In T. gondii, egress is an active process relying on the parasites’ ability to sense that their host cell is dying or dead (Carruthers, 2002).
Much is still to be learned about the proteins and signals involved in either invasion or egress. However, it has become clear that fluctuation in the parasite’s intracellular concentration of calcium ion plays a central role in both these vital events of the parasite’s life cycle. Treatment with agents, such as the Ca2+ ionophore (A23187) which leads to an increase in the intra-parasitic level of calcium, is a potent artificial inducer of parasite egress as early as 2 h post infection (Caldasa et al., 2007). During this process the micronemes discharge their contents, the conoid extends and the parasites become motile.
Toxoplasma gondii acquires host cholesterol that is endocytosed by the Low Density Lipoprotein (LDL) pathway, a process that is specifically increased in infected cells. Interference with LDL endocytosis or cholesterol translocation reduces the intracellular survival of T. gondii (Coppens et al., 2000). Host cell cholesterol is implicated in the entry and replication of an increasing number of intracellular microbial pathogens. However, recently new mechanism has been described by which host cholesterol specifically controls entry of an intracellular pathogen. Briefly, the Parasitophorus Vacuole Membrane (PVM) surrounding T. gondii contains cholesterol. At the time of cell entry, host plasma membrane cholesterol is incorporated into the forming PVM during invasion. Depleting host cell plasma membrane cholesterol blocks parasite internalization by reducing the release of rhoptry proteins that are necessary for invasion (Coppens and Joiner, 2003).
Videomicroscopy studies clearly show that individual tachyzoites exit a host cell at different sites in a manner that morphologically resembles invasion (Black and Boothroyd, 2000). Egress can be stimulated by certain reducing agents such as dithiothreitol (DTT) which is thought to activate parasite derived nucleotide triphosphatases (NTPases) that rapidly consume the host cell’s ATP (Silverman et al., 1998). Precisely how exhaustion of ATP leads to egress remains to be determined.
Proteomics in Toxoplasma gondii: Understanding the differences in protein expression is an essential aspect of elucidating biological phenomena. Proteomics efforts in T. gondii have included general surveys of the proteome of T. gondii tachyzoites as well as other significant subproteomes such as membrane associated proteins and the cytoskeleton (Hu et al., 2006). Gene cloning of P43 surface protein of T. gondii tachyzoite and bradyzoite (SAG3) and preparation of recombinant protein to be used as antigen have been achieved (Kazemi et al., 2007). Parasite subproteomes have been particularly informative. The excreted secreted antigen protein profile consists of microneme and dense granule proteins. This repertoire of secreted proteins is surprisingly complex with evidence for multiple redundant adhesive complexes whose components undergo extensive proteolysis during organelle maturation and host cell invasion (Zhou et al., 2004, 2005). The rhoptry proteome was defined after subcellular fractionation of rhoptry organelles followed by a multiplatform proteomic analysis (Bradley et al., 2005). These studies identified numerous new candidate rhoptry proteins including the kinases and phosphatases that probably mediate the extensive remodeling of host gene expression and metabolism in response to parasite infection.
More recent studies have identified a more extensive and previously unappreciated set of glycosylated proteins. Glycosylation is probably significant for interactions with host cells as well as intracellular signaling. Among the glycosylated proteins are rhoptry proteins, microneme proteins and components of the glideosome, the molecular motor with which T. gondii actively invades host cells (Luk et al., 2008; Fauquenoy et al., 2008). Glycosylated proteins are also a major component of the cyst wall that forms during the stress induced differentiation of bradyzoite forms (Zhang et al., 2001). Proteomic analysis of these unique and biologically significant subproteomes will complement the genome-wide studies currently underway.
Strategies of long-term survival within the host: A mysterious mark of the life cycle of T. gondii is the parasite’s ability to induce long-term chronic infections in any mammalian or avian host. This is facilitated by specific parasite-host adaptations including differentiation processes of the parasite, i.e., conversion from the replicative and lytic tachyzoite stage to the quiescent bradyzoite stage, formation and remodelling of a PV as a safe intracellular compartment and modulation of host cell responses to either external signals or the intracellular infection and also preferential localisation of T. gondii in immune privileged sites like the brain during the chronic phase of infection. All such parasite adaptations remain in need for further elucidation (Luder et al., 2001).
Significant technical improvements in proteome analyses have recently enabled investigations into how intracellular T. gondii manipulates its host cell at a post-transcriptional level. Valuable information about the regulation of glycolysis, lipid and sterol metabolism, the cell cycle, apoptosis and the cytoskeleton in the host cell have been gained (Blader et al., 2001; Saeij et al., 2007). The complexity of detailed analyses of post-translational protein modifications, i.e., phosphorylation, methylation, acetylation, sumoylation and others remains to be resolved. Among the functionally important T. gondii-induced modifications that have already been investigated are the inhibition of inflammatory host responses, the evasion of anti-parasitic effector mechanisms and the prevention of host cell apoptosis by T. gondii.
The anti-inflammatory effects of the parasite: Immunity against T. gondii predominantly relies on Th1-type responses. However, it is now evident that the parasite partially antagonises or even subverts inflammatory host responses. Besides the well-known up-regulation of host anti-inflammatory molecules such as IL-10, Transforming Growth Factor (TGF)-b and lipoxin A4, direct modulations of different pro-inflammatory signaling cascades by intracellular T. gondii have been characterized (Butcher et al., 2005; Shapira et al., 2005). Together, anti-inflammatory effects during Toxoplasma infection restrict the production of IL-12, IFN-c and TNF-a by dendritic cells, macrophages and/or T cells, thereby down-regulating both host immunopathology and anti-parasitic effector mechanisms.
Infection of macrophages or dendritic cells with T. gondii rapidly induces phosphorylation and subsequent inactivation of the cellular NF-jB inhibitor IjB (Butcher et al., 2001) possibly by a kinase activity at the T. gondii PVM (Kim et al., 2004). The nuclear factor jB (NF-jB) and Mitogen Activated Protein Kinase (MAPK) pathways are critical for inducing expression of pro-inflammatory TNF-a and IL-12. Importantly, lipopolysaccharide (LPS)-induced activation of the NF-jB pathway is also temporarily inhibited (Butcher et al., 2001; Molestina and Sinai, 2005), thereby diminishing pro-inflammatory cytokine production in response to inflammatory stimuli. Likewise, early activation of specific MAPKs by T. gondii renders infected macrophages refractory to subsequent LPS-induced IL-12 and TNF-a production (Kim et al., 2004). Recently, parasite-driven activation of Signal Transducers and Activators of Transcription 3 (STAT3) has been shown to suppress LPS-induced TNF-a and IL- 12 production in infected host cells (Butcher et al., 2005). Whereas in non-infected cells STAT3 is activated by IL-10 receptor ligation, in infected cells T. gondii activates STAT3 independently of IL-10 to establish an environment in which anti-inflammatory effects counterbalance pro-inflammatory responses of the host. Remarkably, the ability to modulate STAT3 signaling significantly differs between distinct genotypic lineages of T. gondii and depends on the polymorphic rhoptry kinase ROP16 (Saeij et al., 2007). It is evident that host cell modification by T. gondii is an important pathogenic factor that may also critically influence disease outcome.
Subversion of effector functions of the immune cells: T. gondii has a major impact on regulatory and effector functions of its host cells. Expression of Major Histocompatibility Complex (MHC) class I and II molecules is strongly diminished in T. gondii-infected cells including macrophages and Dendritic Cells (DCs) (Shapira et al., 2005; McKee et al., 2004). Importantly, T. gondii not only fails to activate macrophages and DCs during host cell entry and subsequent intracellular development, but also blocks activation-induced MHC expression. Consequently, the antigen presentation to CD4+ T cells by infected host cells is also inhibited (Shapira et al., 2005; McKee et al., 2004) and it can be hypothesised that this subverts CD4+ T cell expansion and effector functions against the parasite. Expression of MHC class II molecules in macrophages is regulated by IFN-c-triggered activation of the Janus kinase/Signal Transducers and Activators of Transcription (JAK/STAT) pathway. It appears that intracellular T. gondii parasites do not interfere with initial activation and nuclear translocation of the transcription factor STAT1 but nevertheless block the activity of STAT1-dependent promoters (Lang et al., 2006; Kim et al., 2007). Since IFN-c is the major mediator of resistance against T. gondii, such general interference with IFN-c-regulated gene expression may be critical for the parasite’s intracellular survival by evasion of anti-parasitic effector mechanisms.
Modulation of host cell apoptosis: Modulation of host cell apoptosis is a prominent feature of T. gondii infections and may be critical for the course of infection. Toxoplasma gondii can exert opposite effects on the cell death program of the host, being able to both trigger (Gavrilescu and Denkers, 2001; Mordue et al., 2001) and inhibit (Goebel et al., 2001; Payne et al., 2003) apoptosis. Obviously, induction of apoptosis in T and B lymphocytes, Natural Killer (NK) cells, macrophages and granulocytes contributes to evasion of the host’s immune attack during the acute phase of infection (Gavrilescu and Denkers, 2001). Whereas moderate levels of leukocyte apoptosis may both assist intracellular survival of T. gondii and restrict immunopathology, exaggerated leukocyte apoptosis leads to unrestricted parasite multiplication and the host’s death. High levels of apoptosis result from an overwhelming inflammatory response after infection and may be mediated by both Fas/FasL-dependent and TNF-a/TNFR1- dependent mechanisms (Gavrilescu and Denkers, 2001; Mordue et al., 2001). Besides triggering apoptosis in certain cell populations during acute infection, T. gondii exerts prominent anti-apoptotic effects. As with other intracellular pathogens, inhibition of host cell apoptosis by intracellular T. gondii may be required to counteract the intrinsic cellular suicide program of infected cells as well as the extrinsic FasL or granzyme/perforin-mediated killing of infected host cells by T and NK lymphocytes. In recent years, considerable progress has been made in understanding the molecular mechanisms of the anti-apoptotic effects of T. gondii. It has been established that modification of multiple host cell targets at both transcriptional and post-transcriptional levels play an important role (Luder et al., 2009).
The above-mentioned parasite-dependent survival mechanisms all rely on an inhibition of apoptotic signaling pathways. Recently, an alternative explanation has been put forward. It has been shown that death receptor ligation or perforin treatment results in a Ca2+-dependent egress of T. gondii leading to necrotic host cell death (Persson et al., 2007).
CONCLUSION
T. gondii has emerged as one of the most successful models of intracellular parasitism. Genetic and cell biological studies have begun to identify and elucidate the games played by such enigmatic opportunistic parasite for its long- lasting survival and maintained pathogenicity within the host. The components of the host cell invasion management and the specific role of each protein and how these components cooperate to adjust cell entry by T. gondii, all have been precisely discussed. However, it is still not possible to fully understand Toxoplasma invasion , a process found to be an active event highly distinct from host cell entry by other intracellular microorganisms. For the benefit of human health and for the care of infected livestock and hoping for translating enormous advances about Toxoplasma and its interaction with its host, research on Toxoplasma still in need to be continued.
REFERENCES
- Abdul-Fattah, M.M., M.M. Sanad, R.A. Darwish and S.M. Yousef, 1992. Is immunoprotection against Toxoplasma strain specific. J. Egypt. Soc. Parasitol., 22: 77-82.
PubMed - Asgari, Q., D. Mehrabani, M. Moazzeni, F. Akrami-Mohajeri, M. Kalantari, M.H. Motazedian and G.R. Hatam, 2009. The seroprevalence of ovine toxoplasmosis in Fars Province, Southern Iran. Asian J. Anim. Vet. Adv., 4: 332-336.
CrossRefDirect Link - Alexander, D.L., J. Mital, G.E. Ward, P. Bradley and J.C. Boothroyd, 2005. Identification of the moving junction complex of Toxoplasma gondii: A collaboration between distinct secretory organelles. PLoS. Pathog., 1: e17-e17.
PubMed - Beckers, C.J., J.F. Dubremetz, O. Mercereau-Puijalon and K.A. Joiner, 1994. The Toxoplasma gondii rhoptry protein ROP 2 is inserted into the parasitophorous vacuole membrane, surrounding the intracellular parasite and is exposed to the host cell cytoplasm. J. Cell Biol., 147: 947-961.
PubMed - Black, M.W. and J.C. Boothroyd, 2000. Lytic cycle of Toxoplasma gondii. Microbiol. Mol. Biol. Rev., 64: 607-623.
CrossRefDirect Link - Blader, I.J., I.D. Manger and J.C. Boothroyd, 2001. Microarray analysis reveals previously unknown changes in Toxoplasma gondii-infected human cells. J. Biol. Chem., 276: 24223-24231.
CrossRef - Boothroyd, J.C., 2009. Toxoplasma gondii: 25 years and 25 major advances for the field. J. Parasitol., 39: 935-946.
CrossRef - Boothroyd, J.C. and J.F. Dubremetz, 2008. Kiss and Spit: The dual roles of Toxoplasma rhoptries. Nat. Rev. Microbiol., 6: 79-88.
CrossRef - Bradley, P.J. and L.D. Sibley, 2007. Rhoptries: An arsenal of secreted virulence factors. Curr. Opin. Microbiol., 10: 582-587.
CrossRef - Bradley, P.J., C. Ward, S.J. Cheng, D.L. Alexander and S. Coller et al., 2005. Proteomic analysis of rhoptry organelles reveals many novel constituents for hosteparasite interactions in Toxoplasma gondii. J. Biol. Chem., 280: 34245-34258.
PubMed - Butcher, B.A., L. Kim, P.F. Johnson and E.Y. Denkers, 2001. Toxoplasma gondii tachyzoites inhibit proinflammatory cytokine induction in infected macrophages by preventing nuclear translocation of the transcription factor NF-κB1. B. J. Immunol., 167: 2193-2201.
Direct Link - Butcher, B.A., L. Kim, A.D. Panopoulos, S.S. Watowich, P.J. Murray and E.Y. Denkers, 2005. IL-10-independent STAT3 activation by Toxoplasma gondii mediates suppression of IL-12 and TNF-α in host macrophages1. J. Immunol., 174: 3148-3152.
Direct Link - Carruthers, V.B., 2002. Host cell invasion by the opportunistic pathogen Toxoplasma gondii. Acta. Trop., 81: 111-122.
CrossRef - Caldasa, L.A., W. de Souzaa and M. Attias, 2007. Calcium ionophore-induced egress of Toxoplasma gondii shortly after host cell invasion. Vet. Parasitol., 174: 210-220.
CrossRef - Luder, C.G.K., R.R. Stanway, M. Chaussepied, G. Langsley and V.T. Heussler, 2009. Intracellular survival of apicomplexan parasites and host cell modification. Int. J. Parasitol., 39: 163-173.
CrossRef - Camus, D., M.G. Vannier, M.A. Santos and D.M. Banic, 1995. The art of parasite survival. Braz. J. Med. Biol. Res., 28: 399-413.
PubMed - Cerede, O., J.F. Dubremetz, M. Soete, D. Deslee, H. Vial, D. Bout and M. Lebrun, 2005. Synergistic role of micronemal proteins in Toxoplasma gondii virulence. J. Exp. Med., 201: 453-463.
CrossRef - Chinchilla, M., O.M. Guerrero, A. Castro and J. Sabah, 1994. Cockroaches as transport hosts of the protozoan Toxoplasma gondii. Rev. Biol. Trop., 42: 329-331.
PubMed - Cook, A.J., R.E. Gilbert, W. Buffolano, J. Zufferey and E. Petersen et al., 2000. Sources of toxoplasma infection in pregnant women: European multicentre case-control study. Br. Med. J., 321: 142-147.
CrossRefDirect Link - Coppens, I. and K.A. Joiner, 2003. Host but not parasite cholesterol controls Toxoplasma cell entry by modulating organelle. Mol. Bio. Cell., 14: 3804-3820.
CrossRef - Coppens, I., J.D. Dunn, J.D. Romano, M. Pypaert, H. Zhang, J.C. Boothroyd and K.A. Joiner, 2006. Toxoplasma gondii sequesters lysosomes from mammalian hosts in the vacuolar space. Cell., 125: 261-274.
CrossRef - Coppens, I., A.P. Sinai and K.A. Joiner, 2000. Toxoplasma gondii exploits host low-density lipoprotein receptor-mediated endocytosis for cholesterol acquisition. J. Cell Biol., 149: 167-180.
PubMed - Dubey, J.P., 2008. The history of Toxoplasma gondii-the first 100 years. J. Eukaryotic Microbiol., 55: 467-475.
CrossRefDirect Link - Dubey, J.P. and J.K. Frenkel, 1972. Cyst-induced toxoplasmosis in cats. J. Eukaryotic Microbiol., 19: 155-177.
CrossRef - Dubey, J.P. and J.L. Jones, 2008. Toxoplasma gondii infection in humans and animals in the United States. Int. J. Parasitol., 38: 1257-1278.
CrossRefPubMedDirect Link - Dubey, J.P., N.L. Miller and J.K. Frenkel, 1970. Charecterization of the new fecal form of Toxoplasma gondii. J. Parasitol., 56: 447-456.
PubMed - Fauquenoy, S., W. Morelle, A. Hovasse, A. Bednarczyk and C. Slomianny et al., 2008. Proteomic and glycomic analyses of N-glycosylated structures involved in Toxoplasma gondii host cell interactions. Mol. Cell Proteomics, 7: 891-910.
CrossRef - Gavrilescu, L.C. and E.Y. Denkers, 2001. IFN-g overproduction and high level apoptosis are associated with high but not low virulence Toxoplasma gondii infection. J. Immunol., 167: 902-909.
PubMed - Goebel, S., U. Gross and C.G. Luder, 2001. Inhibition of host cell apoptosis by Toxoplasma gondii is accompanied by reduced activation of the caspase cascade and alterations of poly(ADP-ribose) polymerase expression. J. Cell Sci., 114: 3495-3505.
PubMed - Howe, D.K. and L.D. Sibley, 1995. Toxoplasma gondii comprises three clonal lineages: Correlation of parasite genotype with human disease. J. Infect. Dis., 172: 1561-1566.
PubMed - Hu, K., J. Johnson, L. Florens, M. Fraunholz and S. Suravajjala et al., 2006. Cytoskeletal components of an invasion machine e the apical complex of Toxoplasma gondii. PLoS. Pathog., 2: e13-e13.
CrossRef - Kazemi, B., L. Maghen, M. Bandehpour, S. Shahabi and A. Haghighi, 2007. Gene cloning of P43 surface protein of Toxoplasma gondii tachyzoite and bradyzoite (SAG3). Res. J. Microbiol., 2: 170-174.
CrossRefDirect Link - Kim, L., B.A. Butcher and E.Y. Denkers, 2004. Toxoplasma gondii interferes with lipopolysaccharide-induced mitogen-activated protein kinase activation by mechanisms distinct from endotoxin tolerance. J. Immunol., 172: 3003-3010.
PubMed - Kim, S.K., A.E. Fouts and J.C. Boothroyd, 2007. Toxoplasma gondii dysregulates IFN-{γ}-inducible gene expression in human fibroblasts: Insights from a genome-wide transcriptional profiling. J. Immunol., 178: 5154-5165.
PubMed - Lang, C., M. Algner, N. Beinert, U. Gross and C.G. Luder, 2006. Diverse mechanisms employed by Toxoplasma gondii to inhibit IFN-gamma-induced major histocompatibility complex class II gene expression. Microbes Infect, 8: 1994-2005.
PubMed - Lehmann, T., P.L. Marcet, D.H. Graham, E.R. Dahl and J.P. Dubey, 2006. Globalization and the population structure of Toxoplasma gondii. Proc. Natl. Acad. Sci. USA., 103: 11423-11428.
PubMed - Luk, F.C.Y., T.M. Johnson and C.J. Beckers, 2008. N-linked glycosylation of proteins in the protozoan parasite Toxoplasma gondii. Mol. Biochem. Parasitol., 157: 169-178.
CrossRefDirect Link - Luder, C.G., W. Walter, B. Beuerle, M.J. Maeurer and U. Gross, 2001. Toxoplasma gondii down-regulates MHC class II gene expression and antigen presentation by murine macrophages via interference with nuclear translocation of STAT1alpha. Eur. J. Immunol., 31: 1475-1484.
PubMed - Magno, R.C., L. Lemgruber, R.C. Vommaro, W. De Souza and M. Attias, 2005. Intravacuolar network may act as a mechanical support for Toxoplasma gondii inside the parasitophorous vacuole. Microsc. Res. Tech., 67: 45-52.
PubMed - McKee, A.S., F. Dzierszinski, M. Boes, D.S. Roos and E.J. Pearce, 2004. Functional inactivation of immature dendritic cells by the intracellular parasite Toxoplasma gondii. J. Immunol., 173: 2632-2640.
Direct Link - Molestina, R.E. and A.P. Sinai, 2005. Detection of a novel parasite kinase activity at the Toxoplasma gondii parasitophorous vacuole membrane capable of phosphorylating host IkappaBalpha. Cell Microbiol., 7: 351-362.
PubMed - Mordue, D.G., F. Monroy, M. La Regina, C.A. Dinarello and L.D. Sibley, 2001. Acute toxoplasmosis leads to lethal overproduction of Th1 cytokines. J. Immunol., 167: 4574-4584.
Direct Link - Nelson, M.M., A.R. Jones, J.C. Carmen, A.P. Sinai, R. Burchmore and J.M. Wastling, 2008. Modulation of the host cell proteome by the intracellular apicomplexan parasite Toxoplasma gondii. Infect. Immun., 76: 828-844.
CrossRef - Nguyen, T.D., G. Bigaignon, D. Markine-Goriaynoff, H. Heremans and T.N. Nguyen et al., 2003. Virulent Toxoplasma gondii strain RH promotes T-cell-independent overproduction of proinflammatory cytokines IL12 and -interferon. J. Med. Microbiol., 52: 869-876.
CrossRef - Nissapatorn, V., 2007. Toxoplasmosis: A silent threat in Southeast Asia. Res. J. Parasitol., 2: 1-12.
CrossRefDirect Link - Nissapatorn, V., C.K.C. Lee, Y.A.L. Lim, K.S. Tan and I. Jamaiah et al., 2007. Toxoplasmosis: A silent opportunistic disease in HIV/AIDS patients. Res. J. Parasitol., 2: 23-31.
CrossRefDirect Link - Payne, T.M., R.E. Molestina and A.P. Sinai, 2003. Inhibition of caspase activation and a requirement for NF-kappaB function in the Toxoplasma gondii mediated blockade of host apoptosis. J. Cell Sci., 116: 4345-4358.
PubMed - Persson, E.K., A.M. Agnarson, H. Lambert, N. Hitziger and H. Yagita et al., 2007. Death receptor ligation or exposure to perforin trigger rapid egress of the intracellular parasite Toxoplasma gondii. J. Immunol., 179: 8357-8365.
PubMedDirect Link - Sabry, M.A. and W.W. Reda, 2008. Infection by cyst producing Protozoa among human and food producing animals in Egypt. J. Boil. Sci., 8: 889-895.
CrossRefDirect Link - Saeij, J.P.J., J.P. Boyle, S. Coller, S. Taylor and L.D. Sibley et al., 2006. Polymorphic secreted kinases are key virulence factors in toxoplasmosis. Science, 314: 1780-1783.
PubMed - Saeij, J.P.J., S. Coller, J.P. Boyle, M.E. Jerome, M.W. White and J.C. Boothroyd, 2007. Toxoplasma co-opts host gene expression by injection of a polymorphic kinase homologue. Nature, 445: 324-327.
CrossRef - Sanad, M.M. and A.J. Al-Ghabban, 2007. Serological survey of toxoplasmosis among sheep and goats in Tabouk, Saudi Arabia. J. Egypt Soc. Parasitol., 37: 329-340.
PubMed - Shapira, S., O.S. Harb, J. Margarit, M. Matrajt and J. Han et al., 2005. Initiation and termination of NF-kappaB signaling by the intracellular protozoan parasite Toxoplasma gondii. J. Cell Sci., 118: 3501-3508.
PubMed - Sibley, L.D. and J.C. Boothroyd, 1992. Virulent strains of Toxoplasma gondii comprise a single clonal lineage. Nature, 359: 82-85.
CrossRef - Sibley, L.D., J.L. Krahenbuhl, G.M. Adams and E. Weidner, 1986. Toxoplasma modifies macrophage phagosomes by secretion of a vesicular network rich in surface proteins. J. Cell Biol., 103: 867-874.
PubMed - Silverman, J.A., H. Qi, A. Riehl, C. Beckers, V. Nakaar and K.A. Joiner, 1998. Induced activation of the Toxoplasma gondii nucleoside triphosphate hydrolase leads to depletion of host cell ATP levels and rapid exit of intracellular parasites from infected cells. J. Biol. Chem., 173: 12352-12359.
PubMed - Sinai, A.P. and K.A. Joiner, 2001. The Toxoplasma gondii protein ROP2 mediates host organelle association with the parasitophorous vacuole membrane. J. Cell. Biol., 154: 95-108.
PubMed - Tenter, A.M., A.R. Heckeroth and L.M. Weiss, 2000. Toxoplasma gondii: From animals to humans. Int. J. Parasitol., 30: 1217-1258.
CrossRefPubMedDirect Link - Zhang, Y.W., S.K. Halonen, Y.F. Ma, M. Wittner and L.M. Weiss, 2001. Initial characterization of CST1, a Toxoplasma gondii cyst wall glycoprotein. Infect. Immun., 69: 501-507.
CrossRef - Zhou, X.W., M.J. Blackman, S.A. Howell and V.B. Carruthers, 2004. Proteomic analysis of cleavage events reveals a dynamic two-step mechanism for proteolysis of a key parasite adhesive complex. Mol. Cell Proteomics, 3: 565-576.
CrossRefDirect Link - Zhou, X.W., B.F. Kafsack, R.N. Cole, P. Beckett, R.F. Shen and V.B. Carruthers, 2005. The opportunistic pathogen Toxoplasma gondii deploys a diverse legion of invasion and survival proteins. J. Biol. Chem., 280: 34233-34244.
CrossRef