
H.A. Shalaby
Department of Parasitology and Animal Diseases, National Research Center, P.O. Box 12622, Giza, Egypt
F.M. El-Moghazy
Department of Biology, College of Education for Girls- Al-Kharj, Scientific Departments, Al-Kharj University, Kingdom of Saudi Arabia
F.A.M. Khalil
Department of Parasitology and Animal Diseases, National Research Center, P.O. Box 12622, Giza, Egypt
Research Journal of Parasitology
Year: 2010 | Volume: 5 | Issue: 4 | Page No.: 224-230







ABSTRACT
In this study, specificity of snail feet and hepatopancreases antigens in antibody detection of their trematode parasite was evaluated via Western blot technique. Snails, of two different families, antigens; Lymnaea cailliaudi as intermediate host of Fasciola gigantica and Biomphalaria alexandrina as intermediate host of Paramphistomum microbothrium, were evaluated in detection of IgG antibodies against their trematode parasites after preparation of the required Hyper-Immune Sera (HIS) in rabbits. The results revealed higher specificity of snail feet in antibody detection than hepatopancreases antigens. Where, three of sex polypeptides of L. cailliaudi feet antigen; identified by F. gigantica HIS, showed specific positive reactivity. These polypeptides were at molecular weights of 59, 57 and 52 kDa. While, one of sex polypeptides of L. cailliaudi hepatopancreases antigen; identified by F. gigantica HIS, at molecular weight of 57 kDa was specific. Similarly, two polypeptides of B. alexandrina feet antigen; at molecular weights of 54 and 45 kDa, showed specific reactivity toward anti-paramphistomum antibodies. At a time, no specific reactivity had been shown by the antigenically active polypeptides of B. alexandrina hepatopancreases antigen. Owing to its cross-reactivity with incompatible trematode parasite, snail antigen can be used as a general starting antigen for immunodiagnosis of trematode infections.
PDF Abstract XML References Citation
Received: April 25, 2010;
Accepted: June 19, 2010;
Published: August 07, 2010
How to cite this article
H.A. Shalaby, F.M. El-Moghazy and F.A.M. Khalil, 2010. Specificity of Snail Derived Antigens in Diagnosis of their Trematode Parasites. Research Journal of Parasitology, 5: 224-230.
URL: https://scialert.net/abstract/?doi=jp.2010.224.230
URL: https://scialert.net/abstract/?doi=jp.2010.224.230
INTRODUCTION
The concept of antigen sharing between trematodes and their intermediate hosts and the possibility of using common antigens for serodiagnosis of their target parasites are currently receiving considerable attention. This is particularly evident in the area of schistosomiasis, where common antigens between schistosomes and their intermediate hosts were described (Wide et al., 2006). A shared antigen between Schistosoma mansoni, Fasciola hepatica and B. glabrata was isolated, purified and characterized by Rasmussen et al. (1985). This antigen showed a limited value for the specific immunodiagnosis of schistosomiasis. Alarcon de Noya et al. (1989) cleared that serum from Schistosoma infected persons reacted versus crude B. glabrata antigen by ELISA; with sensitivity rate reached up to 100%. Chacon et al. (2002) confirmed the presence of common antigens between S. mansoni and its vector, B. glabrata and suggested that soluble B. glabrata antigen constituted a rich source of candidate antigens for diagnosis and prophylactic studies. Three Bithynia species, the snail hosts of Opisthorchis viverrini, provided antigenic materials for the antibody detection of human opisthorchiasis (Chanawong and Waikagul, 1991). It was found that each of whole body, head-foot and hepatopancreatic tissue antigens of B. funiculata was as effective as crude O. viverrini adult worm antigen for the ELISA detection of O. viverrini antibodies in infected patients.
In previous study, Shalaby (2004) demonstrated successful ELISA detection of Fasciola gigantica IgG antibodies in naturally infected animals using Lymnaea cailliaudi feet and hepatopancreases antigens and higher sensitivity of the latter antigen in antibody detection than the former. At a time, he found by SDS-PAGE that there were shared antigens between F. gigantica excretory-secretory, L. cailliaudi feet and L. cailliaudi hepatopancreases antigens. But, specificity of those antigens in detection of trematode infection still remains as a question. Consequently, in this study, specificity of snails, of two different families, antigens; L. cailliaudi as intermediate host of Fasciola gigantica and B. alexandrina as intermediate host of Paramphistomum microbothrium, in detection of IgG antibodies against their trematode parasites was evaluated using Western blot technique after preparation of the required hyper-immune sera in rabbits.
MATERIALS AND METHODS
The study was undertaken during the period from Oct. 2009 to Feb. 2010, at National Research Center, Egypt.
Preparation of Antigens
Snail Antigens
Snails of two different families were selected; L. cailliaudi as intermediate host of F. gigantica and B. alexandrina as intermediate host of Paramphistomum microbotyrium. Field collected snails were identified according to Brown (1994) and reared in the laboratory for production of laboratory-bred snails according to El-Bahy (1984). They were used for antigen preparation according to Khalil et al. (1985) and El-Bahy and Shalaby (2004). Where, feet and hepatopancreases of laboratory bred mature non-infected snails were dissected. The collected tissues were homogenized in an equal volume of 0.01 M phosphate buffered saline, pH 7.4 (PBS) and sonicated for 5 min under 150 watt interrupted pulse cycle using a sonifier cell disrupter. Then, they were centrifuged at 5000 rpm for 1 h at 4°C. The supernatant was dialyzed in 6-8 kDa dialysis tubes overnight at 4°C against 4 M urea buffer. Thereafter, they were concentrated by absorption against polyvinyl pylorridone. The protein content was measured by the method of Lowry et al. (1951) and stored at -70°C until used.
Parasite Antigens
F. gigantica and P. microbothrium crude worm antigens (CWAs) were prepared from anterior parts of fresh extracted adult worms collected from fresh condemned buffaloes' liver and rumen, respectively, at a local abattoir. The specimens were washed repeatedly in 0.01 M PBS, pH 7.4. and homogenized at 6000 rpm for 20 min and then subjected to high-speed centrifugation (10000 rpm) for 1 h at 4°C. The supernatant was separated as crude antigen after the protein content had been measured as above and stored at -70°C until used (Shalaby, 1998).
Preparation of Rabbit Hyper-Immune Sera (HIS)
Rabbit hyper-immune sera were raised against all previous snail and parasite antigens as described by Langley and Hillyer (1989) via initial subcutaneous injection in an equal volume of Freund’s complete adjuvant and three consecutive intramuscularly injections in an equal volumes of Freund’s incomplete adjuvant during 60 days. The level of specific antibodies in sera of immunized rabbits was evaluated before slaughter. Rabbits were bled before immunization as negative control sera.
Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS- PAGE) and Western Blot Techniques
Protein fractions of tested snail antigens, L. cailliaudi feet, L. cailliaudi hepatopancreases, B. alexandrina feet and B. alexandrina hepatopancreases, were demonstrated using 12% SDS-PAGE (100 μg/lane) according to Laemmli (1970) with the aid of high and low molecular weight standards (Pharmacia Biotech). The fractionated snail antigens were transferred onto nitrocellulose sheet for Western blot technique according to Towbin et al. (1979). The nitrocellulose strips blotted with snail antigens were tested in group of four as in Fig. 2:
| • | The first strip was used to test the recognition of the original molecules of snail antigen by homologous immune sera |
| • | The second one was allowed to react with rabbit HIS raised against the compatible trematode parasite |
| • | The third one was allowed to react with rabbit HIS raised against the incompatible trematode parasite |
| • | The fourth one was allowed to react with rabbit pre-immune control sera |
The molecular weight of specific and non-specific polypeptides was determined using molecular weight standard curve as described by the producer (Pharmacia).
RESULTS
Electrophoretic profile of snail antigens was resolved by SDS-PAGE into multiple components at both high and low molecular weight ranges (Fig. 1). L. cailliaudi feet and hepatopancreases antigens revealed at least 10 and 13 polypeptides, respectively. Those polypeptides molecular weights ranged from 16-160 kDa. Of them, the protein bands at molecular weight of about 66, 45, 34, 28 and 20 kDa in L. cailliaudi feet and 97, 54, 45 and 28 kDa in L. cailliaudi hepatopancreases were major bands (Fig. 1).
Concerning B. alexandrina feet and hepatopancreases antigens, there were 8 and 7 polypeptides in each antigen, respectively. Their molecular weights ranged from 21-97 kDa. The components of 97 and 30 kDa were major bands in B. alexandrina feet while, that of 66, 30 and 20 kDa were major bands in B. alexandrina hepatopancreases (Fig. 1).
To clarify specificity of the recorded snail polypeptides, the adopted Western blot technique (Fig. 2) revealed that antigenically active components in L. cailliaudi feet, on reaction with its homologous HIS, were eight polypeptides at molecular weights of 94, 85, 75, 70, 66, 59, 57 and 52 kDa (Fig. 2, Lane A). All those polypeptides except that of 85 and 75 kDa were recognized by F. gigantica HIS (Fig. 2 Lane B). While, P. microbothrium HIS reacted crossly with three polypeptides at molecular weights of 94, 70 and 66 kDa (Fig. 2, Lane C). At a time, no polypeptides were identified by pre-immune rabbit sera (Fig. 2 Lane D). Therefore, the three polypeptides of 59, 57 and 52 kDa were considered to be specific in detection of anti-Fasciola antibodies.
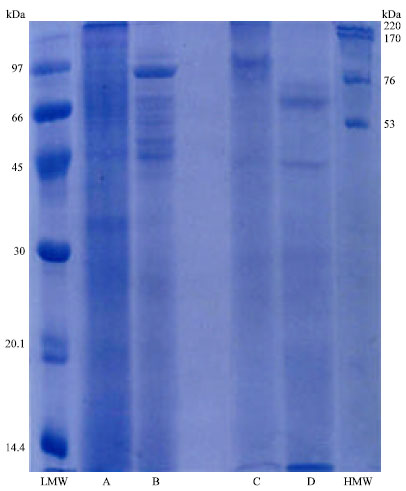 | |
| Fig. 1: | SDS-PAGE of L. cailliaudi feet (Lane A), L. cailliaudi hepatopancreases (Lane B), B. alexandrina feet (Lane C) and B. alexandrina hepatopancreases (Lane D) antigens. LMW. Low Molecular Weight marker; HMW. High Molecular Weight marker |
On the other hand, L. cailliaudi hepatopancreases antigen revealed seven antigenically active polypeptides at molecular weights of 97-85, 70, 57, 54, 45, 42 and 38 kDa (Fig. 2, Lane E). All those polypeptides were recognized by F. gigantica HIS except that of 38 kDa (Fig. 2 Lane F). From those polypeptides, five at molecular weights of 97-85, 70, 54, 45 and 42 kDa reacted crossly with P. microbothrium HIS (Fig. 2, Lane G). The polypeptide of 97-85 kDa was recognized by pre-immune rabbit sera (Fig. 2, Lane H). Thus, only the polypeptide at molecular weight of 57 kDa was specific in detection of anti-Fasciola antibodies.
Concerning B. alexandrina feet antigen, two polypeptides at molecular weights of 54 and 45 kDa of six antigenically active polypeptides recognized by its homologous HIS (Fig. 2, Lane I) showed specific reactivity toward anti-paramphistomum antibodies. Where, both polypeptides were identified by P. microbothrium HIS (Fig. 2, Lane J) and did not react crossly with neither F. gigantica HIS (Fig. 2, Lane K) nor pre-immune rabbit sera (Fig. 2, Lane L). On the other hand, the antigenically active polypeptides of B. alexandrina hepatopancreases antigen (Fig. 2, Lane M) had no specific reactivity toward anti-Paramphistomum antibodies. Where, similar antigenic profiles were observed with both P. microbothrium HIS (Fig. 2, Lane N) and F. gigantica HIS (Fig. 2, Lane O).
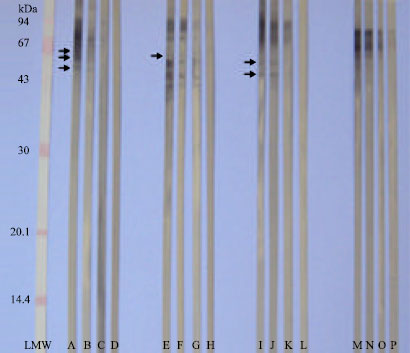 | |
| Fig. 2: | Recognition of different snail antigens by homologus and trematode parasites HIS using Western blot. Lanes A-D. L. cailliaudi feet antigen; Lanes E-H. L. cailliaudi hepatopancreases antigen; Lanes I-L. B. alexandrina feet antigen; Lanes M-P. B. alexandrina hepatopancreases antigen; Lanes A, E, I and M. With homologus HIS; Lanes B, F, J and N. With compatible trematode parasite HIS; Lanes C, G, K and O. With incompatible trematode parasite HIS; Lanes D, H, L and P. With pre-immune control sera; LMW. Low Molecular Weight marker stained with Ponceau S solution; Arrows point to the specific bands |
DISCUSSION
Detection of circulating anti-snail antibodies in trematode infected definitive hosts might prove of value in diagnosing trematode infection (Rasmussen et al., 1985). It had been proposed that the presence of common antigens between some parasites and their hosts would facilitate the search for diagnostic targets, when observing the abundance of antigenic material that could be obtained from those hosts, as demonstrated in fasciolosis. The existence of shared antigens between F. gigantica and L. cailliaudi had been shown by the presence of anti-snail antibodies in animals naturally infected with the parasite. The opposite had also been demonstrated in animals immunized with uninfected snails, which had shown anti-Fasciola antibodies (Shalaby, 2004). The previous study by Shalaby (2004), demonstrated higher sensitivity of L. cailliaudi hepatopancreases than feet antigens in detection of anti-Fasciola antibodies but, their specificity was not shown. That concept was extended in the present study to evaluate specificity of snail feet and hepatopancreases antigens in antibody detection of their trematode parasite via Western blot technique. The results revealed higher specificity of the former antigen in antibody detection than the latter. Where, three of the sex polypeptides of L. cailliaudi feet antigen; identified by F. gigantica HIS, showed specific positive reactivity. These polypeptides were at molecular weights of 59, 57 and 52 kDa. While, one of the sex polypeptides of L. cailliaudi hepatopancreases antigen; identified by F. gigantica HIS, at molecular weight of 57 kDa was specific. Similarly, two polypeptides of B. alexandrina feet antigen; at molecular weights of 54 and 45 kDa, showed specific reactivity toward anti-paramphistomum antibodies. At a time, no specific reactivity had been shown by the antigenically active polypeptides of B. alexandrina hepatopancreases antigen. In that sense, Watthanakulpanich et al. (1997) evaluated crude antigens, prepared from the whole body, head-foot and hepatopancreatic tissue of B. funiculata, for the ELISA detection of O. viverrini antibodies in infected patients. Although, it was promising to find that the snail antigens could be used to detect O. viverrini, cross-reactions with Paragonimus and Strongyloides occurred. In a chromatographic study (Waikagul et al., 2002), four peaks of B. funiculata extract were obtained and analyzed by indirect ELISA. Each of four peaks had a sensitivity of 100, 27.5, 28.8 and 37.5%, respectively and a specificity of 36, 96, 52 and 82%, respectively. They suggested that electroeluted antigen at molecular weight of 53 kDa, notwithstanding its lower antigenicity, might have an important role in diagnosis of opisthorchiasis (Waikagul et al., 2002).
The extensive cross-reactivity of snail antigens with incompatible trematode confirmed the findings reported by Kemp et al. (1982) and Rasmussen et al. (1984) in which antigens obtained from S. mansoni adult worms by affinity chromatography through an anti-B. glabrata column reacted strongly with serum of humans and mice infected with F. hepatica. Moreover, Rasmussen et al. (1985) isolated, purified and characterized shared antigens between S. mansoni and B. glabrata from S. mansoni adult worms through a CNBr-activated Sepharose 4B column coupled with rabbit IgG prepared against a homogenate of B. glabrata hepatopancreases which would be of use as a specific immunodiagnostic test for schistosomiasis. The eluted antigen (designated as SMw-53P) failed to qualify as a candidate for such a reagent due to its cross-reactivity with F. hepatica.
The present study provided further evidence of shared antigens between trematode parasite and its intermediate snail host and extended these observations by demonstrating the specificity of these antigenic epitopes. These epitopes showed limited degree of immunologic specificity due to its cross-reactivity with incompatible trematode parasite. Therefore, snail antigen can be used as a general starting antigen for immunodiagnosis of trematode infections. Its advantage is that large amounts of material can be obtained rapidly and at comparatively lower cost than trematode antigens which require maintenance of a complex life cycle.
REFERENCES
- Wide, A., J. Capaldo, N. Zerpa, R. Pabon and A. Noda et al., 2006. Sharing of antigens between Plasmodium falciparum and Anopheles albimanus. Rev. Inst. Med. Trop. Sao Paulo, 48: 327-332.
PubMed - Chacon, N., S. Losada, B. Noya, B. Alarcon de Noya and O. Noya, 2002. Antigenic community between Schistosoma mansoni and Biomphalaria glabrata: On the search of candidate antigens for vaccines. Mem. Inst. Oswaldo Cruz, 97: 99-104.
Direct Link - Chanawong, A. and J. Waikagul, 1991. Laboratory studies on host-parasite relationship of Bithynia and liver fluke, Opisthorchis viverrini. Southeast Asian J. Trop. Med. Public Health, 2: 235-239.
Direct Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Langley, R.J. and G.V. Hillyer, 1989. Detection of circulating parasite antigen in murine fascioliasis by two-site enzyme-linked immunosorbent assays. Am. J. Trop. Med. Hyg., 41: 472-478.
PubMedDirect Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Rasmussen, K.R., G.V. Hillyer and W.M. Kemp, 1985. Isolation and partial characterization of an antigen shared between Schistosoma mansoni, Fasciola hepatica and Biomphalaria glabrata. J. Parasitol., 71: 792-798.
Direct Link - Towbin, H., T. Staehelin and J. Gordon, 1979. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA., 76: 4350-4354.
PubMedDirect Link - Waikagul, J., P. Dekumyoy, K. Chaichana, M.T. Anantapruti, C. Komalamisra and V. Kitikoon, 2002. Serodiagnosis of human opisthorchiasis using cocktail and electroeluted Bithynia snail antigens. Parasitol. Int., 51: 237-247.
CrossRef - Watthanakulpanich, D., J. Waikagul, M.T. Anantapruti and P. Dekumyoy, 1997. Evaluation of Bithynia funiculata snail antigens by ELISA in serodiagnosis of human opisthorchiasis. Southeast Asian J. Trop. Med. Public Health, 3: 593-598.
Direct Link