
M.A. Sabry
Department of Zoonoses, Faculty of Veterinary Medicine, Cairo University, Egypt
E.S. Taher
Research Institute of Ophthalmology, Department of Parasitology, El-Giza, Egypt
E.M.H. Meabed
Department of Parasitology, Faculty of Medicine, Fayoum University, Egypt
Research Journal of Parasitology
Year: 2009 | Volume: 4 | Issue: 4 | Page No.: 105-114







ABSTRACT
The aim of the present study was to investigate the possible public health significance of calves’ giardiasis in the village. Faecal examination of children and their contact calves revealed infection by Giardia cysts up to 25 and 30.8%, respectively. Protein fraction at molecular weight of 33 kDa as a giardins fraction was found to be closely related to zoonotic strain of calves’ Giardia using western blot analysis. Amplification and sequencing of a 292 bp fragment of 16S-rRNA ribosomal unit from 25 calves Giardia isolates using nested PCR, revealed that 20% of these isolates were belonging to the zoonotic Giardia Assemblage A, while the rest of the isolates (80%) were in Assemblage of livestock genotype (E). This level of calve zoonotic Giardia genotype demonstrates the possible role of calves as a reservoir for human infection in this village.
PDF Abstract XML References Citation
How to cite this article
M.A. Sabry, E.S. Taher and E.M.H. Meabed, 2009. Prevalence and Genotyping of Zoonotic Giardia from Fayoum Governorate, Egypt. Research Journal of Parasitology, 4: 105-114.
URL: https://scialert.net/abstract/?doi=jp.2009.105.114
URL: https://scialert.net/abstract/?doi=jp.2009.105.114
INTRODUCTION
The intestinal protozoan parasite, Giardia duodenalis (Syn. Giardia intestinalis, Giardia labmlia) has recently emerged as an important cause for diarrhoea in human and animals. It is distributed worldwide and presents high levels of genetic diversity. The disease is most common in developing countries and other areas where sanitation and hygiene are poor (Nygard et al., 2006). Increased prevalence in human as well as in some of the surrounding animals offers an emerging concern about the role played by some animals in human giardiasis (Olson, 2004). About 200 million people in the world are infected with G. lamblia. Each individual eliminates up to 900 million cysts per day. Higher prevalence is found in tropical and subtropical areas, in urban than in rural where G. lamblia affects up to 30% of the population (Minvielle et al., 2004).
Other than detection of Giardia stages in faeces by traditional ways, several enzyme immunoassay kits are also available for detecting soluble stool antigens. However, they are qualitative; can not distinguish between genotypes and unable to detect low levels of infection. Moreover, false negative results have been obtained in some cases (Guy et al., 2004). Characterization of specific and sensitive protein fraction from the parasite antigens and diagnosis by immunoblot technique may offer some degree of accuracy.
Several protein fractions extracted from Giardia species cysts were described as being specific for diagnosis of infection using immunoblot assays. These fractions at Molecular Weight (MW) corresponding to 21-39, 66,78, 92, 103 kDa (Reiner et al., 1989) 26, 28, 38, 42, 46 kDa (Campbell and Faubert, 1994) 29, 75, 88, 102 kDa (Erlandsen et al., 1986) and at 112.19, 65.13, 52.3, 28.9, 21.3 kDa (Ghattas, 2005).
Trout et al. (2004) determined a specific fraction at MW of 29-33 kDa from human G. lamblia and identified them as giardins. Fractionation of giardins using sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS-PAGE) revealed five protein bands assigned with relative MW of 38, 36, 34, 33 and 29 kDa. These fractions were considered to be specific for detection of anti-parasite antibodies (Ab) in sera of infected cases (Peattie et al., 1989).
Lack of morphological differences between genetic variants of Giardia found in mammals has resulted in an informal categorization of these genotypes based on genetic differences (Guy et al., 2004). Thus, recent application of molecular characterization procedures based on PCR has made an enormous contribution to the understanding the genetic structure of Giardia species, zoonotic transmission and molecular epidemiology of infection (Thompson, 2000).
Molecular characterization and phylogenetic analysis of G. duodenalis isolates from different hosts revealed the existence of seven major genotypes. Assemblages A and B have the widest host ranges, encompassing humans and a variety of other animals. Assamblages C and D have been reported only in dogs, assemblage E has been reported only in hoofed livestock, assemblage F has been reported only in cats and assemblage G has been reported only in rats (Hsu et al., 2007).
Cattle are susceptible to infection with two genotypes of G. duodenalis, the zoonotic genotype (Assemblage A), or the non-zoonotic livestock genotype (Assemblage E) (Olson, 2004). Moreover, Lalle et al.(2005) stated that only assemblages A and B appeared to be associated with human infections, but G. duodenalis isolates within these assemblages have also been identified from various animals, including cattle, dogs and cats. In Egypt, the genotyping study of G. intestinalis is still limited (Foronda et al., 2008).
During the last year, an emerging diarrhoea problem was recorded in children in a small village at Tamia-Fayoum Governorate, Egypt. Giardia trophozoites and cysts were diagnosed on examination of the children faecal samples. Most people in this area were farmers and lived in intimate contact to cattle and calves. For this reason, the current study was aimed to investigate the possible significance of calves’ giardiasis on public health in this area. The prevalence of Giardia in a selected group of children and calves with a history of diarrhea was estimated. The relation between anti Giardia antibodies in sera and antigenic fractions of different Giardia isolates was investigated. Finally Giardia cysts isolated from calves were characterized at the molecular level and genotype.
MATERIALS AND METHODS
Sample Collection and Examination
Between September and November 2006, identified faecal and blood samples were collected from 180 children and 182 beef cattle calves from El-Azezia village at Tamia Province, Fayoum Governorate, Egypt. Two age groups, males and females, were investigated (one week-less than one year) and (1-2 years old). Both children and calves presented with diarrhoea or previously had history of diarrhoea. Rectal or fresh deposited faecal samples were preserved in an ice boxes and were transferred directly to Department of Zoonoses, College of Veterinary Medicine, Cairo University for examination. Giardia cysts had been detected. The level of infection was estimated by light microscopy after analysis using modified Telemann technique (including lugol’s staining) according to Gamboa et al. (2003). Blood samples were left to clot then sera were separated by centrifugation and stored at -20°C until used.
Cyst Purification and Disruption
According to O’Handley et al. (2000), suitable amount of faecal samples were filtered through two layers of surgical gauze and expressed to yield approximately 7 mL of filtrate. The filtrate was layered over 5 mL of 1 M sucrose (specific gravity = 1.13) then centrifuged at 800x g for 5 min to concentrate the cysts at the sucrose/water interface. The interface and upper layer of liquid were transferred using a pipette to a clean centrifuge tube and re-centrifuged at 800x g for 5 min. After decanting the supernatant, the pellet was suspended in PBS to a volume of 1 mL and stored at 4°C until used.
Cysts were disrupted according to Minvielle et al. (2008). The purified cysts were subjected to six cycles of freezing-thawing (-70°C/+70°C, 30 min) then it proceeded to one of two uses, (1). For DNA isolation, the mixture was incubated with proteinase K (1 mg mL-1) and lysis buffer (NaCl 0.15 M, EDTA 0.1 M, SDS 0.5%, pH 7.8, at 37°C for 24 h. This suspension was stored at -20°C. (2) For Giardia cyst antigen extraction according to Vinayak et al. (1978), the mixture was centrifuged (15000 rpm for 1 h at 4°C). The supernatant was aspirated and used as crude Giardia cyst antigen/after estimation to its protein content, stored at -70°C until use.
Fractionation of Giardia Cyst Antigen
Crude Giardia cyst antigen/(represent human and animals from each farm were fractionated using SDS-PAGE according to (Laemmli, 1970). Using PROTEAN II Xi cell, Bio-Rad, the antigens were resolved using 1.5 mm thickness, in 12% polyacrylamide gel slabs in Tris-glycine buffer (pH 8.3) under reducing conditions. The stacking gel consisted of 5% acrylamide prepared in 12.5 mM Tris-HCl buffer (pH 6.7) (Sigma chemical Co.). Prestained MW standard was employed (Bio-Rad Cat.#161-0318). The comb was adjusted as one small well for standard and one large for the sample. The electrophoretically separated proteins were transferred to nitrocellulose paper (0.2 μm pore size) according to the method of Towbin et al. (1979), by transferring them overnight at 30 V and then for 2 h at 70 V.
Western-Blot Assay (EITB)
Giardia cyst specific protein fractions were determined using EITB according to Towbin et al. (1979). Longitudinal NC strips (15x0.3 cm) contained Giardia species fractionated antigens were cut out, treated with infected serum (1:100) collected from infected animals of known cyst/gram faeces in comparison with negative control sera. Protein A-peroxidase conjugate (Sigma Immunochemicals) was used at 1:2000 dilution and finally the colour developed in 4-chloro-1-naphthol substrate. Specific Giardia protein fractions are that reacted positively versus its target antibody and in the same time did not react after treatment of similar strips by negative serum samples. In order to investigate the relation between anti-Giardia antibodies in sera and antigenic structures of different Giardia isolates, sera of infected human and calves were selected from each study site (that have high cyst/gram faeces) and tested separately versus NC strips carrying fractionated Giardia cyst of human (one isolate) or animal (2 isolates/farm) origin separately.
Giardia Cyst Purification and DNA Extraction
Approximately 100 μL of sucrose-purified cyst suspension from selected Giardia positive calves samples were used for DNA extraction using Genomic DNA purification kit (Gentra system Instruction, Minnesota USA), following the instructions and guided by O’Handly et al. (2000) with modification. Samples were subjected to six freeze-thaw cycles in liquid nitrogen. Then 100 μL of tissue lysogenic buffer was added to each sample.
Samples were incubated at 80°C for 5 min to lyse the cells and centrifuged at 10000x g for 15 sec. The supernatant was transferred to a clean tube and 15 μL RNase solution was added. Mix for 25 times and incubated at 37°C for 15-60 min. The samples were left to cool at room temperature; 1 mL protein precipitation solution was added to the cell lysate, exposed to high speed vortices for 20 sec then centrifuged at 2000x g for 10 min. The supernatant containing the DNA was poured into a clean tube containing 3 mL of absolute Isopropanol, mixed well then centrifuged at 2000x g for 3 min, where the DNA was precipitated. The supernatant was discarded and the DNA washed several times by addition of 70% cold ethanol, DNA was precipitated again by centrifugation (2000x g min-1). The alcoholic supernatant was poured out and 200 μL DNA hydration solutions were added and left for 1 h at 65°C. Spectrophotometer quantitization of the DNA was done according to Manose et al. (1989) and stored at -20°C for PCR amplification. DNA was extracted from Giardia cyst isolates collected from 5 highly infected animals representing each farm.
Amplification and Sequencing
A fragment of the 16S-rRNA ribosomal unit was amplified using the PCR protocol described by Appelbee et al. (2003) after modification. A nested PCR protocol was used, utilizing initial primers Gia2029 (5'-AAGTGTGGTGCAGACGGACTC-3') and Gia2150c (5'-CTGCTGCCGTCCTTGGATGT-3') amplifying a 497 bp product and secondary primers RH11 (5'-CATCCGGTCGATCCTGCC-3') and RH4 (5'-AGTCGAACCCTGATTCTCCGCCAGG-3') generating a 292 bp fragment. PCR reactions consisted of 1-5 μL DNA, 2.0 mM MgCl2, 200 μM each dNTP, 5% DMSO, 12.5 pmol each forward and reverse primer, 0.5 Units of Taq polymerase (Qiagen, Germany), 0.5 Units of TAQ Extender™ (Stratagene, USA) to minimize PCR error and 1xPCR buffer (Qiagen). Thirty-five PCR cycles (96°C for 45 sec, 55°C for 30 sec, 72°C for 45 sec) with an initial hot start (96°C for 4 min) and final extension (72°C for 4 min) was carried out on a Perkin-Elmer Gene/Amp PCR System 2400 (USA). The secondary PCR cycle conditions were identical to the primary round except that the annealing temperature was increased to 59°C. PCR products were purified using Qiagen PCR Agarose Purification Kit (Qiagen, 28004) and sequenced in both directions using ABI Prism™ Rhodamine Terminator Cycle Sequencing Ready Reaction Kit (Applied BioSystems, California) according to the manufacturers instructions, except that the annealing temperature was reduced to 59°C, the cycles increased to 35 and reaction volumes halved to 10 μL. Sequences were analyzed using SeqEd v1.0.3. (Applied BioSystems). Alignment of sequence information obtained from calves’ isolates were compared with that obtained from GenBank accession numbers AF113902 and AF199446u) using CLUSTAL W (Thompson et al., 1994).
RESULTS
Microscopic examination of faecal samples collected from 180 children and 182 calves in two age groups from five farms as described in Table 1 revealed that the prevalence of infection was higher in calves (30.8%) than in their contact children (25%). Infection was also higher in the group of less than one year old (34.9 and 28.7%) than in the older one (27.1 and 22%) both in calves and children, respectively. Infection rate varied between different sampling sites, from 20 to 41.2% in calves and from 20 to 38.9% in children and the higher rate of infection was still in the group of less than one year old.
Using EITB technique, sensitivity and specificity of fractionated Giardia cysts isolates versus different sera was detected (Fig. 1). The data demonstrated that treatment of human fractionated Giardia isolates on NC strips using known infected human sera revealed a number of 10 protein fractions at MW of 21, 26, 29, 33, 38, 60, 68, 78, 92 and 102 kDa. (Lane 2).
| Table 1: | Prevalence of Giardia cysts in faeces of children and their contact calves |
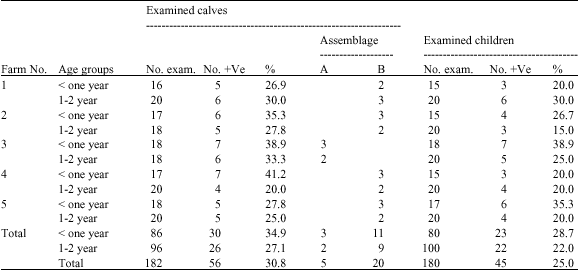 | |
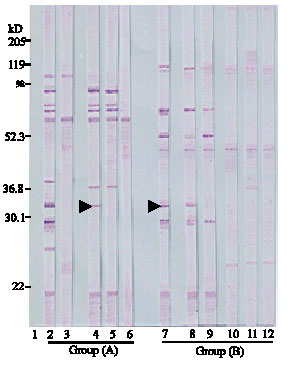 | |
| Fig. 1: | Reaction of different Giardia isolates fraction versus infected human and animal sera. Lane 1: Molecular weight protein standard. Group (A) fractionated Giardia cysts (Human isolates): Lane 2: NC strip reacted versus infected child sera, Lane 3: NC strip reacted versus negative healthy child sera, Lane 4: NC strip reacted versus infected calve sera (farm 3) (see reaction at 33 kD, arrow), Lane 5: NC strip reacted versus other infected calve sera (No reaction at 33 kDa) and Lane 6: NC strip reacted versus negative healthy calve sera. Group (B) fractionated Giardia cysts (Calve isolates): Lane 7: NC strip reacted versus infected calve sera, Lane 8: NC strip (Farm 3 isolate) reacted versus infected human sera (reaction at 33 kD, arrow), Lane 9: NC strip (Other farm isolate) reacted versus infected human sera (no reaction at 33 kD), Lane 10: NC strip reacted versus negative health calve sera, Lane 11: NC strip (Farm 3 isolate) reacted versus negative human sera and Lane 12: NC strip (Other farm isolate) reacted versus negative human sera |
| Table 2: | Genetic sequences of 16-rRNA gene of Giardia species isolates from beef calves in Egypt |
 | |
The fractions at 102 and 60 kDa falsely reacted versus non-infected human sera (Lane3). Screening of infected calves’ sera versus the same fractions revealed that serum samples collected from infected animals in farm No.3 (lane 4) demonstrated positive reactions versus 6 fractions at MW of 21, 33, 36.8, 68, 78 and 92 kDa. In the same time, the same fractions except for that at 33 kDa (arrow) were detected by the selected sera from the other farms (Lane 5). None of the above mentioned fractions reacted versus negative calves’ sera (Lane 6).
With treatment of 5 separated Giardia cysts (calves isolates) versus infected calves sera collected from different farms, seven protein fractions reacted at MW of 21, 29, 33, 46, 52.3, 68 and 112 kDa (lane 7). Bands at MW of 46 and 112 kDa were recorded with negative serum samples (lane10). Treatment of NC strips carrying the fractionated Giardia cysts isolated from calves in farm No. 3 versus sera of infected human demonstrated the same previous protein bands (Lane 8). While treatment of fractionated Giardia cysts isolates from other farms versus the same infected human sera (lane 9), detected all of the same previous bands except the fraction at 33 kDa. None of these bands were diagnosed using negative human sera versus both types of Giardia calves isolates (Lane 11-12).
Concerning the fraction at MW of 33kDa, this fraction was diagnosed in human isolates versus human infected sera and also with calves’ sera from farm No. 3 only. The same fraction in calves’ isolates was diagnosed by different infected calves’ sera, but infected human sera reacted versus this band in fractionated Giardia isolates; originated from calves of farm No.3 only.
From 58 calves’Giardia isolates, a number of 25 isolates were selected, five samples representing infected animals in each farm. The samples were sequenced at the 16S-rRNA locus using nested PCR. Twenty isolates from farm No. 1, 2, 4 and 5 (80%) had similar sequences to the previously described genotype (Assemblage) A sequence except that they had a single nucleotide difference, A-G transition at the nucleotide position 92. These sequences aligned with the proposed Hoofed livestock genotype (AF113902) (E). The isolates selected from farm number three (5 isolates; 20%) had the identical genetic sequence to Assemblage A genotype (AF199446) (Table 2).
DISCUSSION
Giardiasis has received an increasing attention in recent years, partly because the infections in animals cause numerous outbreaks in human. This is usually associated with contamination of fruits and vegetables through contaminated irrigation water and manure fertilizer (Nygard et al., 2006). Despite giardiasis is self limited and age dependant, the chronic infection leads to malabsorption, villus atrophy, diffuse shortening of microvilli, reduced disaccharidase activity, loss of epithelial barrier function, increased permeability and enterocyte apoptosis (Buret, 2007).
The parasite has a complex series of transmission routes, including anthroponotic and zoonotic transmission, as well as waterborne and foodborne transmission. The shed cysts are remarkably stable and can survive for several weeks or months in the environment. Furthermore, the infective dose is low, as even a single cyst carries some probability of causing infection. In addition, most faeces that contain cysts end up in the environment and can spread to foods by irrigation or by direct contact. Viable cysts can persist in water, as routine treatments eliminate only a fraction of these stages (Cacci, 2004).
The present study which was conducted to investigate the impact of calves’ giardiasis on human health in a small Egyptian village was based on the following assumptions. Did the disease spread from calves to children or vice versa? Can Immunoblot analysis suggest the presence of genetic variations between different isolates? And finally, what are the genotypes of calves’ giardiasis in this village?
The prevalence of giardiasis in the study area was estimated using microscopic examination as recommended by Guy et al. (2004), who considered microscopic examination the standard method for diagnosis. Infection rates were 30.8% in calves and 25% in children with higher prevalence in individuals less than one year old. This incidence was lower than that mentioned by Foronda et al. (2008) as they recorded a higher percentage of 34.6% in children in Egypt. This difference may be attributed to the difference in the study areas. On the other hand, the obtained rate of infection in calves was in agreement with Ghattas (2005). The present data recorded higher prevalence in younger than in older groups. This was in agreement with Buret (2007). However, Olson (2004) stated that giardiasis was not age dependant.
At the level of fecal examination, no wide difference was detected between the rate of infection of the examined calves and children. So, it was difficult to determine the source of infection (human or calves). For this reason the study was directed to evaluate the antigenic relations between the human and animal Giardia cyst isolates. In an attempt to determine the antigenic relations between Giardia cysts isolates from children and that from calves, Western blot technique was adopted versus sera of infected cases. The reacted protein fractions for Giardia cysts either from animal or human isolates were in accordance with that previously mentioned by Reiner et al. (1989), Campbell and Faubert (1994) and Ghattas (2005) using the same technique. Presence of some differences in the molecular weight and number of the diagnosed fractions between the present study and some of these authors may be related to difference in the concentration of the acrylamid used during fractionation as well as due to difference in the localities.
This part of the study clarifies the presence of an interesting specific band at MW of 33 kDa. This protein band was diagnosed in fractionated human isolates after treatment with infected human sera. It could also react with calve sera collected from farm No. 3 only. On testing fractionated calves’ isolates, this protein band reacted with sera of different infected calves. On treatment of different calve isolates versus infected human sera; this band was also detected in Giardia isolates extracted from calves of farm No. 3 only. This protein fraction comes within the MW range defined by Trout et al. (2004) as one of giardins fractions since giardins MW range from 29-33 kDa. The same researchers considered giardins as one of the most specific antigens in Giardia. In the researchers opinion, Western blot analysis for this specific band can play a role for potential detection of suspected zoonotic genotypes especially if several isolates of the parasite were tested by human and animal sera form the same geographic locality. This can help in exclusion of some isolates and reduce the number of samples that will be amplified using PCR.
Progress in genotyping facilitates clear determination of various strains of Giardia of humans and animal origin (Santin et al., 2007). In order to investigate the possible public health significance of calves’ giardiasis, Giardia cysts isolated from calves were characterized at a molecular level and genotyped. The choice of a particular assay mainly depends on the amount of information carried by the genetic locus under analysis (Cacci, 2004). The locus of 16SrRNA conceded as one of the highly conserved gene regions used in the detection of Giardia (Appelbee et al., 2003). In the present study, 25 isolates were selected representing approximately 50% of the total isolates from 5 farms. They were genotyped using nested PCR. Amplification and sequencing of a 292 bp fragment of the 16s-rRNA ribosomal unit indicated that the examined calves in this study were predominantly (80%) infected with G. duodenalis livestock genotype, while Assemblage A genotype was represent 20%. The same ratio between the two Assemblages was previously recorded by Appelbee et al. (2003) in dairy calves in Canada. Also, Read et al. (2004) mentioned that livestock-specific assemblage E of G. duodenalis appears to be the most frequent genotype in calves. However, this genotype is not considered of a zoonotic importance (Santin et al., 2007). Moreover, the potentially zoonotic Assemblage A. was previously found in calves (Trout et al., 2004). Diagnosis of Giardia Assemblage A in calves of farm (3) in particular highlighted the value of the protein fraction at 33 kDa. This protein fraction was previously detected in the isolate of this group only when tested versus infected human sera. So this protein fraction can be closely related to the zoonotic genotype of calves Giardia.
The frequency of Giardia transmission is rather high among calves. Although, calves may be exposed to more than one genotype, the livestock genotype does not compete with human genotype in calves. The results suggest that calves can be infected by livestock genotype. They can also be infected with a genotype potentially infectious to humans according to the nearest source of infection in the area.
In the present study, the hoofed livestock genotype of Giardia is predominant in the examined calves. However, the zoonotic genotype, Assemblage A, represented 20% and diagnosed in one farm only. Even at this level and according to O’Handly et al. (2000), calves infected with Giardia commonly shed from 105 to 106 cysts/g faeces. Consequently, a few calves infected with genotype assemblage A isolates could pose a significant public health risk. In a recent study in Egypt, Foronda et al. (2008) sequenced the DNA for G. intestinalis of humans and found that the Assemblage B genotype was the most prevalent (80%). Moreover, 15% of the positive samples belonged to Assemblage E and 5% of them belonged to Assemblage A. They further clarified the presence of Assemblage E in humans for the first time. However, further examination of other areas is required to confirm this result. In the scope of the present study, Foronda et al. (2008) data could increase the risk of cattle as a source of zoonotic giardiasis if their results were proved.
For conclusion it is important to note that this is the first study about genotype of calve giardiais in Egypt. The study proved that calves could act as an important reservoir for human giardiasis via both direct transmission and contamination of water catchments areas. These findings recommend application of strict control measures to minimize the spreading of infection from calve farms to cause outbreaks in the surrounding people.
REFERENCES
- Appelbee, A.J., L.M. Frederick, T.L. Heitman and M.E. Olson, 2003. Prevalence and genotyping of Giardia duodenalis from beef calves in Alberta, Canada. Vet. Parasitol., 25: 289-294.
CrossRefDirect Link - Buret, A.G., 2007. Mechanisms of epithelial dysfunction in giardiasis. Gut, 56: 316-317.
CrossRefDirect Link - Cacci, S.M., 2004. New methods for the diagnosis of Cryptosporidium and Giardia. Parasitologia, 46: 151-155.
PubMedDirect Link - Campbell, J.D. and G.M. Faubert, 1994. Recognition of Giardia lamblia cyst-specific antigens by monoclonal antibodies. Parasite Immunol., 16: 211-219.
PubMedDirect Link - Foronda, P., M.D. Bargues, N. Abreu-Acosta, M.V. Periago and M.A. Valero(et al)., 2008. Identification of genotypes of Giardia intestinalis of human isolates in Egypt. Parasitol. Res., 103: 1177-1181.
CrossRefDirect Link - Gamboa, M.I., J.A. Basualdo, M.A. Córdoba, B.C. Pezzani, M.C. Minvielle and H.B. Lahitte, 2003. Distribution of intestinal parasitoses in relation to environmental and sociocultural parameters in La Plata, Argentina. J. Helminthol., 77: 15-20.
CrossRefPubMedDirect Link - Guy, R.A., C. Xiao and P.A. Horgen, 2004. Real-time PCR assay and genotype differentiation of G.lamblia in stool specimens. J. Clin. Microbiol., 42: 3317-3320.
Direct Link - Hsu , B.M., H.Y. Wun and P.C. Hsu, 2007. Prevalence and genotyping of Giardia in husbandry system in Taiwan. Parasitol. Res., 101: 275-280.
PubMedDirect Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Lalle, M., E. Pozio, G. Capelli, F. Bruschi, D. Crotti and S. Caccio, 2005. Genetic heterogeneity at the giardin locus among human and animal isolates of Giardia duodenalis and identification of potentially zoonotic genotypes. Int. J. Parasitol., 35: 207-213.
CrossRef - Minvielle, M.C., N.B. Molina, D. Polverino and J.A. Basualdo, 2008. First genotyping of G. lamblia from human and animal feaces in Argentina, South America. Mem. Inst. Oswaldo Cruz, Rio de Janeiro, 103: 98-103.
Direct Link - O'Handly, R.M., M.E. Olson, D. Fraser, P. Adams and R.C. Thompson, 2000. Prevalence and genotype characterisation of Giardia in dairy calves from Western Australia and Western Canada. Vet. Parasitol., 90: 193-200.
CrossRef - Peattie, D.A., R.A. Alonso, A. Hein and J.P. Caulfield, 1989. Ultrastructural Localization of Giardins to the edges of disk microribbons of G.lamblia and the nucleotide and Deduced protein sequence of alpha giardin. J. Cell Biol., 109: 232-235.
Direct Link - Read, C.M., P.T. Monis and R.C.A. Thompson, 2004. Discrimination of all genotypes of Giardia duodenalis at the glutamate dehydrogenase locus using PCR-RFLP. Infect. Genet. Evol., 4: 125-130.
CrossRefDirect Link - Reiner, D.S., H. Douglas and F. Gilin, 1989. Identification and localization of cyst-specific antigens of G. lamblia. Infect. Immun., 57: 963-968.
Direct Link - Santin, M., J.M. Trout and R. Fayer, 2007. Prevalence and molecular characterization of Cryptosporidium and Giardia species and genotypes in sheep in Maryland. Vet. Parasitol., 146: 17-24.
CrossRefDirect Link - Thompson, R.C.A., 2000. Giardiasis as a re-emerging infectious disease and its zoonotic potential. Int. J. Parasitol., 30: 1259-1267.
Direct Link - Thompson, J.D., D.G. Higgins and T.J. Gibson, 1994. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res., 22: 4673-4680.
CrossRefPubMedDirect Link - Towbin, H., T. Staehelin and J. Gordon, 1979. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA., 76: 4350-4354.
PubMedDirect Link