
Sanjukta Manna
Department of Zoology, Immunoparasitology Research Unit, University of Calcutta, 35 Ballygunge Circular Road, Kolkata-700019, India
Rajdeep Chakravarty
Molecular Genetics Laboratory, Indian Institute of Chemical Biology,
4 Raja SC Mallick Road, Kolkata-700032, India
Buddhadeb Manna
Department of Zoology, Immunoparasitology Research Unit, University of Calcutta, 35 Ballygunge Circular Road, Kolkata-700019, India
Amal Bhattacharya
Department of Zoology, Immunoparasitology Research Unit, University of Calcutta, 35 Ballygunge Circular Road, Kolkata-700019, India
Research Journal of Parasitology
Year: 2006 | Volume: 1 | Issue: 1 | Page No.: 48-58







ABSTRACT
Considering the invasion strategy of the Leishmania parasite the molecules present on the cell surface of promastigotes are of paramount importance as the parasite membrane antigens are the interface between the parasite and its vertebrate and insect hosts. Their topography and receptor interactions with host cell decide the acceptability, survival, multiplication, pathogenesis or killing of the parasite in the host. Gamma radiation had interesting effects on living cells and proteins, allowing new epitopes to express but parasites presented unequal radiosensitivity, related to their genome organization. After exposure to gamma radiation from Co60 source, Leishmania donovani undergo profound changes in ultrastructure, microviscosity, protein profile and carbohydrate residues of plasma membrane. Upto 20 krad, the membrane integrity was maintained but above this dose the microtubules showed chracteristic fluence dependent changes in the number of protofilaments upon gamma radiation. Agglutination progressively decreased from nonirradiated to irradiated promastigotes. The most prominent epitopes at 53 and 66 kDa were expressed at 10 and 20 krad irradiated parasites surface. It was also indicated that 20 krad was the limiting dose at which cell can survive and compensate for altered conditions by a process known as homeoviscous adaptation so it can remain in the fluid phase even as the environment changes.
PDF Abstract XML References
How to cite this article
Sanjukta Manna, Rajdeep Chakravarty, Buddhadeb Manna and Amal Bhattacharya, 2006. Radiation Induced Alterations in Membrane Fluidity, Microtubular Structure, Glycoconjugates and Protein in Leishmania donovani. Research Journal of Parasitology, 1: 48-58.
URL: https://scialert.net/abstract/?doi=jp.2006.48.58
URL: https://scialert.net/abstract/?doi=jp.2006.48.58
INTRODUCTION
Visceral leishmaniasis (VL or kala-azar) caused by Leishmania donovani and Leishmania infantum in the old world and Leishmania chagasi in the new world, is the most dreaded disease and devastating among various forms of leishmaniasis (Dube et al., 2005). The disease has gained significance because of high mortality rate particularly amongst the children and poor people. In India high incidence has been reported from the states of Bihar, Assam, West Bengal and Eastern Uttar Pradesh where resistance and relapse are on increase (Dube et al., 2005).The annual incidence is estimated to about 600,000 new officially reported cases, the global prevalence is 12 million cases and the population at risk totals approximately 350 million (WHO, 1995).
Treatment of leishmaniasis with sodium stibogluconate, pentamidine or amphotericin B is expensive and potentially toxic and the administration of these drugs may require a prolonged stay in hospital. Thus the mere chemotherapy is not sufficient to combat the disease. Hence, in addition to the therapeutic measures, vaccination is the point of serious consideration. Immunization against leishmaniasis was achieved in the past by inoculating humans with living parasites that induced localized self-healing cutaneous lesion. Since then, a first generation of vaccines composed of formulations including killed parasites were developed against cutaneous but not visceral leishmaniasis and were used in large clinical trials on human populations of endemic areas (Handman, 1997; Khalil et al., 2000). For that, the authors are prompted to probe into the role of radiation, if any, on L. donovani to modulate its membrane antigen which is very much responsible for its virulence and infectivity, subsequently it may be utilized for further vaccine development. The perspective of the present investigation arose from several documents of application of radiation in parasitology especially in host parasite interface as has been highlighted by many workers in several human diseases (Rivier et al., 1993). Leishmania donovani resides and multiplies in hydrolytic environments throughout its developmental cycle, extracellularly in the insect vector alimentary tract and within the phagolysosomal system of mammalian macrophages (Dwyer, 1980). The plasma membrane of the parasite constitutes the major site of physical and physiological interactions between the host and the parasite. It plays a role in energy transduction and in maintenance of ion and solute gradients for a constant intracellular environment. They also provide a milieu where biological reactions can occur and act to regulate cellular growth and metabolism.
This is the very site where parasites extract nutrients to sustain their life and where they stop the parasiticidal actions of the hosts. Even more demanding are the cell surface related tasks for Leishmania, which must deal with not only extracellular but also intracellular microenvironments of the host. The promastigote surface displays several antigens; receptors such as surface carbohydrates (Grogl et al., 1987 ); lipophosphoglycan (McConville et al., 1992); ectoenzyme, as well as protease (gp 63) ( Murray et al., 1990 ); acid phosphatase (Glew et al., 1988) which help receptor mediated internalization of the parasite within macrophages (Channon et al., 1984 ). It is, therefore, considered of interest to study the changes occurring in certain membrane parameters of irradiated parasites which will be crucial for the outcome of somewhat modified infection and probable vaccine development. This study reports on the effect of gamma radiation on the membrane fluidity, ultrastructural changes, surface glycoconjugates and the expression of antigenic protein in promastigotes.
MATERIALS AND METHODS
Parasites and Culture Conditions
Promastigotes of Leishmania donovani (MHOM/IN/1983/AG83) were grown at 22°C in medium 199 (pH 7.4) supplemented with 10% heat inactivated fetal bovine serum, 2 m ML-glutamine, 100U of penicillin G sodium and 100 g of streptomycin sulfate per ml and subcultured in the same medium at an average density of 2x106 cell mL-1.
Irradiation
The late log phase cell culture (3.7x106cells mL-1) was taken for radiation exposure and exposed to a Co60 gamma source for irradiation at 23°C using doses in the range of 10, 20, 30 and 40 krad (Alexander, 1982). The dose rate (12 krad h-1) was measured with Fricke dosimeter (Spinks and Woods, 1976). The irradiated promastigotes were harvested from culture by centrifugation at 1500 xg at 4°C and taken for the experiment.
Preparation of Cell Membrane
The surface membrane of both nonirradiated and irradiated parasite was isolated and purified by the modification (Gottlieb and Dwyer, 1981) of the method originally devised by Dwyer (1980). Washed cells was resuspended in Tris-EDTA buffer (10 mM Tris-HCl, 2 mM EDTA, pH 8.0) and kept on ice for 30 min. The cells were disrupted by repeated sonication (MSE Soniprep 150 Sonicator) in TM buffer (20 mM Tris-HCl, 3 mM MgCl, pH 8.0) and homogenates were centrifuged at 8000xg for 30 min. The resulting pellets were resuspended in TM buffer and 0.146 M sucrose. The 10 mL of this membrane suspension was layered on 0.73 M sucrose in TM buffer and centrifuged at 8000 xg for 30 min. Pellet was again resuspended and pooled to a total volume of 24 mL in 0.152M sucrose in TM buffer. Two milliliter aliquots of this suspension was overlaid onto discontinuous gradients consisting of sequential layer of 1.23 M(12 mL), 1.52 M (20 mL) and 1.75 M(6.5 mL) sucrose in TM buffer in cellulose nitrate tubes and centrifuged at 42,000xg (Sorvall Ultra 80, rotor AH-627) for 8 h. The pellet membrane washed several times in TM buffer plus 0.438 M sucrose and kept on ice at -70°C until use.
Determination of Membrane Microviscosity (mv)
Both nonirradiated and irradiated parasite membrane mv was determined by fluorescence depolarization measurements using DPH (1,6-Diphenyl-1,3,5-hexatriene) as the fluorescent probe (Shinitzky and Barenholz, 1978).The promastigote membrane in phosphate buffered saline(pH 7.2) were incubated with DPH(5 M final concentrations) at 37°C for 60 min with constant stirring in the dark. The extraneous probe was washed off after the incubation period and the pellets were finally suspended in 2.0 mL of PBS. Measurements of fluorescence anisotropy were carried out using spectrofluorophotometer (Hitachi F-4500) equipped with rotating polarizers at room temperature, with excitation and emission wavelengths of 360 and 432 nm respectively (Dutta et al., 1990). All measurements were made promptly after incubation in order to avoid diffusion of the probe. After background correction of fluorescence intensities with an unlabelled sample, anisotropy were calculated from the following relationship:
where, ![]() and
and ![]() are the respective intensities observed through the polarizer oriented parallel and perpendicular to the plane of polarization of the excitation beam. The degree of polarization (p) was calculated as the difference between fluorescence intensities(
are the respective intensities observed through the polarizer oriented parallel and perpendicular to the plane of polarization of the excitation beam. The degree of polarization (p) was calculated as the difference between fluorescence intensities( ![]() ) divided by the sum of these two intensities(
) divided by the sum of these two intensities( ![]() ). The polarization of fluorescence reflects the mobility of the fluorescent molecules and transfer of excitation energy between them. From the fluorescence anisotropy ® and the maximal limiting fluorescence anisotropy (r0; it is 0.362 for DPH), the microviscosity [ = (r0/r-1)-1] were calculated in each case.
). The polarization of fluorescence reflects the mobility of the fluorescent molecules and transfer of excitation energy between them. From the fluorescence anisotropy ® and the maximal limiting fluorescence anisotropy (r0; it is 0.362 for DPH), the microviscosity [ = (r0/r-1)-1] were calculated in each case.
Transmission Electron Microscopic Preparation for Membrane
The pelleted membrane was washed twice in PBS (pH 7.2) and fixed for atleast one hour, at room temperature in 2.5% (v/v) glutaraldehyde plus 1% (v/v) paraformaldehyde in 0.1 M cacodylate buffer, pH 7.3 following Pral et al. (2003). After rinsing with the same buffer, the pellets were post fixed with 1% (w/v) osmium tetroxide for one hour, rinsed with saline, dehydrated in upper graded alcoholic series and embedded in epoxy resin. Then ultra thin sections were cut in Lieca Ultracut UCT microtome, stained sequentially in methanolic uranyl acetate and lead citrate at 4°C. The thin sections were examined in a Philips CM-10 Transmission Electron Microscope at accelerating voltage of 80 kv.
Preparation for Lectin Mediated Agglutination
The fluorescein isothiocyanate(FITC)-conjugated lectins ConcanavalinA (ConA: Sugar specificity -D-glucose, -D-mannose) (obtained from Sigma Chemical Co. St.Louis, Missouri) was adjusted to final concentrations of 1,000, 500, 250, 125, 62.5, 31.2, 15.6 and 7.8 g mL-1 in phosphate buffered saline. Lectin binding experiments were carried out after Bose et al. (1989) and observed with a Zeiss Incident Fluorescence Microscope (excitation 495 nm; Absorption filter 520 nm).
Sodium Dodecyl Sulfate-polyacrylamide Gel Electrophoresis (SDS-PAGE) of Membrane Protein
Soluble antigens for SDS-PAGE were prepared from nonirradiated and irradiated parasite membrane according to Leon et al. (1986). Dialyzed and lyophilized antigens were stored at-70°C in aliquots until use. Antigen preparations were analyzed by SDS-PAGE using 10% polyacrylamide resolving gels with 4% stacking gels in the 1.5M Tris-HCl buffer system, pH 8.8 and 6.8, respectively (Laemmli, 1970). Each sample was mixed with an equal volume of sample loading buffer (1M Tris-HCl, pH 6.8, 5% mercaptoethanol, 2.3% SDS, 10% glycerol, 0.1% bromophenol blue) and solubilized by boiling at 100°C for 15 min. The samples applied contained equal amounts of protein (50 μg/well) and electrophoresis (Bio-Rad ProteanII, Richmond, California) was performed at 25 mA (constant current) until the tracking dye reached the bottom of the gel. The resolved proteins were then stained with 0.25% (w/v) Coomassie blue.
RESULTS
Membrane Microviscosity (mv)
With increasing radiation doses, the emission anisotropy value decreases, so the fluidity increases (Fig. 1) When fluidity increases, the packing of membrane and its flexibility getting ruptured. The emission anisotropy value of 40 krad irradiated promastigotes was three times lower than that of nonirradiated promastigotes, therefore, the membrane of irradiated cells of higher doses was with more fluid than that of nonirradiated cells. This change in membrane fluidity is probably a consequence of the radiation induced modification of membranous lipid metabolism. Membrane fluidity is inversely proportional to anisotropy. Membrane fluidity of irradiated cells gradually increased with increase of radiation doses, but there was a break point at 20 krad dose (Fig. 1). At this dose, fluidity decreased and as anisotropy increased and approximately same to non irradiated cells.
Membrane Ultrastructure
On the basis of the present study, comparatively easy procedure involving controlled hypotonic shock was developed for preparing deflagellated round unsealed ghosts of membrane with original polarity. Total scanning of the field of both nonirradiated and irradiated promastigotes plasma membrane revealed that longitudinal parallel arrays of subpellicular membrane microtubules (MT) remained attached to the isolated unsealed cell membrane envelopes (Fig. 2) which run in parallel along the longitudinal axis of the cell body except in the region of the flagellar pocket. Characteristically, these MT were intimately associated with the PM inner lamina of intact cells and presumably function as cytoskeleton for structural support of the overlaying surface membrane. In nonirradiated promastigotes, individual microtubule was composed of linearly arranged protofilaments. The individual protofilaments were in turn composed of adjacent spheroid subunits i.e., vesicles with a diameter of 3.5 nm. Sidearm projections 13 nm long, were spaced periodically at 20 nm intervals along the lateral, longitudinal axis of individual microtubule. The sidearms presumably function as crossbridges anchoring the microtubule to the inner lamina of the surface membrane. The leishmanial cell surface lacked membrane ridges and folds and was incapable of endocytosis.
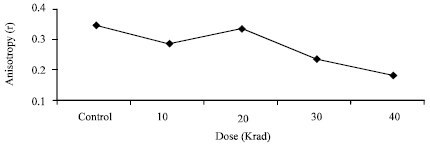 | |
| Fig. 1: | Change of membrane fluidity in different radiation doses |
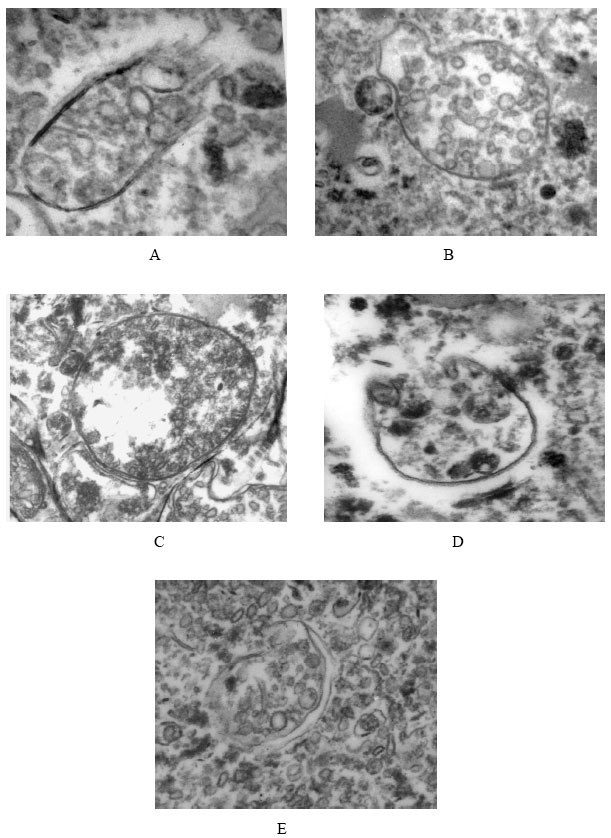 | |
| Fig. 2: | Electron microscopy of cytoplasmic membrane of nonirradiated and irradiated L.donovani. A. Non irradiated . Longitudinally arranged subpellicular microtubules are seen attached to the isolated PM. Vesicles of individual microtubules are iscernible. B: 10 krad rradiated. Afloculent filamentoid nexus appears to connect the microtubules to each other and to the inner lamina of the PM. C: 20 krad irradiated Microtubules densely vesiculate and inner architecture remain same that of nonirradiated. D: 30 krad irradiated. Vesicles do not show any obvious discontinuity and microtubules ended subterminally. E: 40 krad irradiated. Presence of broken membrane and vesicle aggregation are evident |
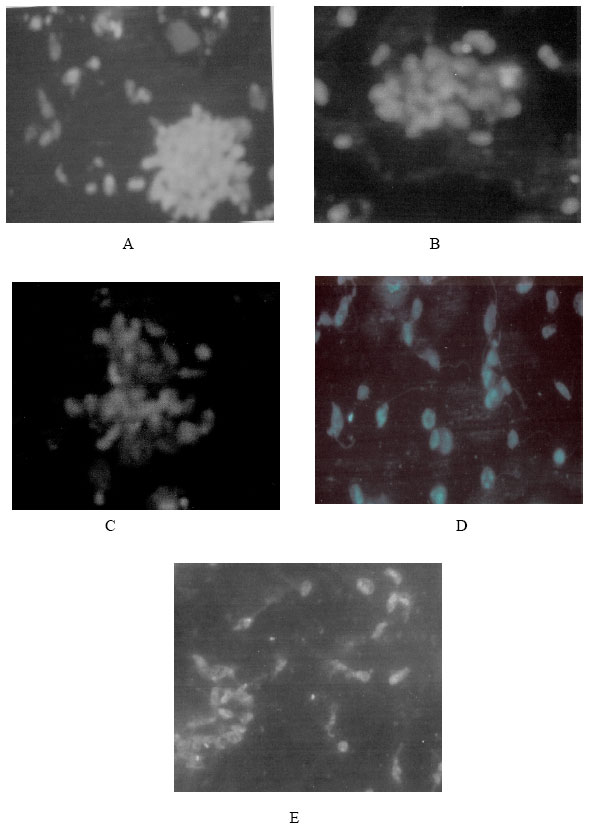 | |
| Fig. 3: | FITC-labeled ConA at a concentration of 500 μg mL-1 seen binding in a homogeneous pattern to non irradiated and irradiated promastigotes and the agglutination behaviour. A: Non irradiated, B: 10 krad irradiated, C: 20 krad irradiated, D:.30 krad irradiated, E: 40 krad irradiated |
In case of irradiated promastigotes the membrane integrity was lost with increasing radiation doses. It was found that at lower radiation doses, the vesicles did not show any obvious discontinuity and appeared to be sealed. The microtubules showed characteristic fluence dependent changes in the number of vesicles upon gamma radiation. In an attempt to quantify the surface density of these vesicles precisely, a large number of electron micrographs were examined. It was found that the number of plasma membrane vesicles was reduced by 6.4, 25.5, 45.1 and 49% of the normal vesicle density at gamma radiation of 10, 20, 30 and 40 krad, respectively. Upto 20 krad, the membrane integrity was maintained and could resist the shear forces. After that, with increasing radiation doses, the presence of some broken membranes and a few vesicles of original polarity were evident. Vesicle aggregation was also observed at 40 krad irradiated promastigote membrane. With further increase in radiation doses, there was total disruption of microtubuler structure and the vesicles of the ghost membrane scattered throughout the field and showed the irreparable damage.
Membrane Glycoconjugates
Several exposed carbohydrate residues on the L.donovani surface were detected by using lectin FITC-ConA. Agglutination experiments with nonirradiated and irradiated promastigotes incubated at room temperature with lectins were shown in Fig. 3. There was marked differences observed in the binding intensity of nonirradiated and irradiated promastigotes with FITC-ConA. The degree of agglutination was compared with control. The brightness of fluorescence were recorded as number of pulses (1+ to 4+) and used as a measures of the intensity of lectin binding at room temperature and were shown in Table 1 with all experiments performed a minimum of 3 times. Lectins listed as negative were tested at concentrations upto 500 μg mL-1 without detectable binding. Highest agglutination was noticed in case of nonirradiated parasites. The descending order of agglutination was observed in case of 10, 20, 30 and 40 krad irradiated parasites. Twenty krad irradiated parasites showed moderate response.
Analysis of Plasma Membrane Proteins
A characteristic change in the gel profile of plasma membrane proteins was observed, with certain protein bands reduced in intensity or missing completely (Fig. 4). Most major SDS-PAGE separated PM peptides banded in the range of 66-45 kDa.
| Table 1: | Intensity of agglutination |
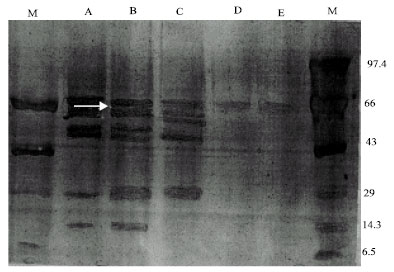 | |
| Fig. 4: | Sodium dodecyl sulfate-polyacrylamide gel electrophoresis of the purified plasma membrane fraction of nonirradiated and gamma irradiated Leishmania donovani. Lane A: Non irradiated, Lane B: 10 krad irradiated, Lane C: 20 krad irradiated, Lane D: 30 krad irradiated, Lane E: 40 krad irradiated,. Lane M: Marker protein. Arrowhead (white) indicates proteins that showed comparative decrease upon irradiation |
The most prominent bands in these gels had the apparent molecular weight of 53 and 65 kDa. There was no significant difference in the protein composition of membrane of nonirradiated, 10 and 20 krad irradiated parasites suggesting that the proteins that were lost were the same irrespective of radiation fluence. However, a gradual decrease in the intensity of certain bands (53, 45, 21 and 13 kDa) was observed with increasing doses of gamma radiation, indicating that the extent of proteins released from the membrane was proportional to the radiation fluence. In case of 30 and 40 krad irradiated parasites, a smear of membrane proteins was observed at 66 kDa (Fig. 4).
DISCUSSION
A substantial body of data in the field of radiation research published during the past decades sparked a debate between the seemingly two-edged beneficial vs. harmful nature of invisible and sometimes sinister radiation (Luckey, 1982). Research on the effects of radiation on protozoan mostly suggests significant inhibition on their life processes. Gamma radiation appears to be a strong membrane disorganizing agent (Mody et al., 1991). Cytoplasmic membrane provides a milieu where biological reactions can occur and act to regulate cellular growth, metabolism and stabilize protein structure which is important with regard to the function of membrane embedded enzymes.
Biological membranes have a fluid mosaic structure in which wide ranges of proteins are embedded in a lipid bilayer (Singer and Nicolson, 1972). It behaves as two-dimensional fluid in which molecules can rotate and move in lateral directions. Maintaining the proper fluidity of cytoplasmic membrane is important for optimum functioning of membrane proteins. Fluidity primarily related to lipid packing is determined by the shape of head group composition (Denich et al., 2003). However, there are other factors that may affect membrane lipid packing such as water, protein and carbohydrate content, while pH, temperature and radiation will also effect packing of phospholipids. It is clear from the present study that a correlation exists between fluidity of membrane and particular radiation doses on leishmanial cells. In present finding it is evident that beyond the limiting dose of gamma radiation a surviving phase exists in which these leishmanial cells probably survive but cannot grow. The radiation induced changes of plasma membrane was further confirmed by the decrease in vesicle density on the microtubule of the plasma membrane (Fig. 2). Irradiated cells have different levels of injury. The deformities of regular microtubular arrangement are increased at higher radiation doses. Although this damage does not always contribute to cell death but the damages are irreparable. This result was also supported by the membrane protein profile. Gamma-radiation induced degradation (cleavage) or modification (linking) of membrane proteins is clearly evident from the absence of certain protein bands on SDS polyacrylamide gels (Fig. 4).
The extent of radiation damage to the plasma membrane, in terms of both structural and functional changes was directly proportional to the gamma radiation and showed characteristic saturation kinetics. Only a few epitopes or perhaps only one, 65 kDa protein have their expression effectively increased on the parasite surface and was presumed, on the basis of its MW and the apparent complete solubility of PM-MT complexes in hot SDS-2ME buffer, to be the constituent microtubular protein, tubulin (Dwyer, 1980). One of the main findings of the present study is that the expression of a particular epitope from the major surface antigen of L. donovani promastigotes increases progressively as the radiation trigger in vitro transformation from promastigote to amastigote-like form. The experiments were performed to determine whether the surface expression of this epitope is involved in promastigote infectivity in vitro. It appears that, at least two mechanisms of surface antigenic change are associated: first, suggested by Sacks and Perkins (1984), is the appearance of new antigenic components on the parasite surface; the second, is the increasing expression of an epitope already present on the surface of infective promastigotes. The level of expression of 65 kDa major surface glycoprotein is down regulated in higher doses to a level that is not detectable under the experimental conditions used. Such radiation induced degradation or modification of membrane proteins could be due to any of the mechanisms proposed for protein inactivation by radiation (McLaren, 1955). These include (i) splitting of disulfide bonds of cystine or photooxidation of tyrosine, thereby changing the tertiary structure of proteins or (ii) ring cleavage in the aromatic amino acids such as tryptophan and phenylalanine. The resultant change in the protein structure, particularly for a transmembrane protein, would effect its interaction with surrounding lipids or with the neighbouring proteins, resulting in the release of damaged protein and alteration of membrane fluidity.
Membrane carbohydrate characterization of different Leishmania species of Indian origin by labeled and unlabeled lectins has been reported previously (Ghosh et al., 1990). The current paper provides additional information on nonirradiated and irradiated leishmanial membrane carbohydrate residues. ConA molecule is known to accommodate both D-mannose and D-glucose residues (Goldstein and Hayes, 1978). FITC-ConA bind to both nonirradiated and irradiated promastigotes of L.donovani. The fluorescent probe attach to plasma membrane and flagellar pocket. Fluorescence intensity progressively decreased from nonirradiated to irradiated promastigotes. This observation agrees with the suggestion that radiation induced transformation involves reduced exposure of subterminal galactose residues (Howard et al., 1987).
Attachment and phagocytosis of Leishmania sp. by mammalian macrophages is suspected that attachment of these organisms may occur via an exposed glycoconjugate on the parasite surface (Wyler and Suzuki, 1983). In the present study, we have surveyed such glycoconjugates on L.donovani by using lectins, documenting some surface characteristics that remain stable and some that evolve through the infective to noninfective stage conversion, which can provide hightened response.
It is clear that the action of gamma radiation on the plasma membrane does not reflect mutagenic alteration in the genes governing membrane properties since: (i) the effects were observed immediately after irradiation, before any genotypic alteration could be expressed; (ii) on examining a large number of electron micrographs, the significant irreparable membrane damage were found after 20 krad irradiation doses.
It was established that the phagocytosis of Leishmania by host macrophages is mediated by the interaction of ligands on the surface of parasite and macrophage receptor (Garg et al., 2005). From the above study it is finally concluded that the radiation dose of 20 krad is the standard dose to modify the surface ligands of the pathogen to obtain maximum protection against the infection and provides a chemical basis for defining the mechanisms by which the irradiated parasite circumvents destruction by the infected host.
ACKNOWLEDGMENTS
The authors are thankful to Head, Department of Zoology, University of Calcutta for providing laboratory facilities. Financial assistance in the form of Junior research fellowships to Sanjukta Manna from the Inter University Consortium, Department of Atomic Energy Facilities (An Autonomous Advance Research Institution of UGC, Govt. of India), Calcutta Centre is gratefully acknowledged. The authors are indebted to Dr T.C. Nag, Assistant Professor, Department of Anatomy, All India Institute Medical Sciences, New Delhi, India for his assistance with electron microscopy.
REFERENCES
- Bose, K., D.K. Ghosh and A. Bhattacharya, 1989 1989. Membrane carbohydrate characterization of Acanthamoeba astronyxis, A castellanii and Naegleria fowleri by fluorescein-conjugated lectins. Intl. J Parasitol., 19: 737-741.
PubMedDirect Link - Channon, J.Y., R.B. Robertes and J.M. Blackwell, 1984. A study of the differential respiratory burst activity elicited by promastigotes and amastigotes in murine resident peritoneal macrophages. Immunology, 53: 345-355.
Direct Link - Denich, T.J., L.A. Beaudette, H.Lee and J.T. Trevors, 2003. Effect of selected environmental and physicochemical factors on bacterial cytoplasmic membranes. J. Microbiol. Meth., 52: 149-182.
Direct Link - Dube, A., S. Naik, A. Pal, J.K. Srivastava and R. Garg, 2005. Isolation of integral membrane proteins of Leishmania promastigotes and evaluation of their prophylactic potential in hamsters against experimental visceral leishmaniasis. Vaccine, 23: 1189-1196.
CrossRefDirect Link - Dwyer, D.M., 1980. Isolation and partial characterization of surface membranes from Leishmania donovani promastigotes. J. Protozool., 27: 176-182.
PubMedDirect Link - Dube, A., S. Naik, A. Pal, J.K. Srivastava and R. Garg, 2005. Isolation of integral membrane proteins of Leishmania promastigotes and evaluation of their prophylactic potential in hamsters against experimental visceral leishmaniasis. Vaccine, 23: 1189-1196.
CrossRefDirect Link - Ghosh, D.K., K.N. Ghosh, A. De and A. Bhattacharya, 1990. Kinetoplastid flagellates: surface reactive carbohydrates detected by fluorescein-conjugated lectins. J. Parasitol., 76: 130-133.
Direct Link - Grogl, M., E.D. Franke, P.B. McGreevy and R.E. Kuhn, 1987. Leishmania braziliensis: protein, carbohydrate and antigen differences between log phase and stationary phase promastigotes in vitro. Exp. Parasitol., 63: 352-359.
PubMedDirect Link - Gottlieb, M. and D.M. Dwyer, 1981. Protozoan parasite of humans: Surface membrane with externally disposed acid phosphatase. Science, 212: 939-941.
CrossRefDirect Link - Howard, M.K., G. Sayers and M.A. Miles, 1987. Leishmania donovani metacyclic promastigotes: Transformation in vitro, lectin agglutination, complement resistance and infectivity. Exp. Parasitol., 64: 147-156.
Direct Link - Khalil, E.A.G., A.M. El. Hassan, E.E. Zijlstra, M.M. Mukhtar and H.W. Ghalib et al., 2000. Autoclaved Leishmania major vaccine for prevention of visceral leishmaniasis: a randomized, double blind. BCG-controlled trial in Sudan. Lancet, 356: 1565-1569.
Direct Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Leon, L.L., M.V. Haegaerden and Galvao-Castro, 1986. Comperative studies of Leishmania soluble antigens by crossed-immunoelectrophoresis: Demonstration of specific antigens to L. mexicana and L. braziliensis. Bull. WHO., 64: 113-119.
Direct Link - Luckey, T.D., 1982. Physiological benefits from low levels of ionizing radiation. Health Phys., 43: 771-789.
PubMedDirect Link - McLaren, A.D., 1955. Relations among cystine content, molecular weights and ultraviolet light sensitivity of proteins. Biochem. Biophys. Acta., 18: 601-602.
PubMedDirect Link - McConville, M.J., S.J. Turco, M.A.J. Ferguson and D.L. Sacks, 1992. Developmental modification of LPG during differentiation of Leishmania major promastigotes to an infectious stage. EMBO J., 11: 3593-3600.
Direct Link - Mody, R., B. Mody and P. Dave, 1991. Damage to the plasma membrane in Escherichia coli K-12 induced by far-ultraviolet radiation and its repair. Rad. Res., 127: 156-163.
Direct Link - Pral, E.M.F., R.M. MdaLuz, J.M.F. Balanco, V.R. Teixeira, R.V. Milder and S.C. Alfieri, 2003. Growth phase and medium pH modulate the expression of proteinase activities and the development of megasomes in axenically cultivated Leishmania (Leishmania) amazonensis amastigote-like organisms. J. Parasitol., 89: 35-43.
PubMedDirect Link - Rivier, D., R. Shah, P. Bovay and J. Mauel, 1993. Vaccine development against cutaneous leishmaniasis: Subcutaneous administration of radioattenuated parasites protects CBA mice against virulent L. major challenge. Parasite. Immunol., 15: 75-84.
PubMedDirect Link - Singer, S.J. and G.L. Nicolson, 1972. The fluid mosaic model of the structure of cell membranes. Science, 175: 720-731.
CrossRefDirect Link - Shinitzky, M. and Y. Barenholz, 1978. Fluidity parameters of lipid regions determined by fluorescence polarization. Biochim. Biophys. Acta., 515: 367-394.
PubMedDirect Link - Wyler, D.J. and K. Suzuki, 1983. In vitro parasite-monocyte interactions in human leishmaniasis: Effect of enzyme treatments on attachment. Infect. Immun., 42: 356-361.
Direct Link