
O.M. Ogundele
Department of Anatomy, Bingham University, Karu, Nigeria
J. Madukwe
Department of Histopathology, National Hospital, Abuja, Nigeria
O.A. Omotosho
Department of Anatomy, Bingham University, Karu, Nigeria
O. Adeeyo
Department of Anatomy, Bingham University, Karu, Nigeria
C.O. Akintayo
Department of Anatomy, Bingham University, Karu, Nigeria
E.A. Caxton-Martins
Department of Anatomy, University of Ilorin, Ilorin, Nigeria
Journal of Medical Sciences
Year: 2012 | Volume: 12 | Issue: 3 | Page No.: 92-98







ABSTRACT
Immunohistochemical localization of CD3 and Neuron Specific Enolase (NSE) to demonstrate neuronal metabolism in proliferation and cell recovery was carried out in this study to evaluate morphological and cellular changes in cyanide treated perfused cortical tissues of adult wistar rats in vitro. Four separately perfused tissues were stained with antibodies specific for neuron specific enolase using antigen retrieval method and colour reaction involving 3’3’-Di aminobenzidine tetrachloride (DAB) in peroxidase anti peroxidase method. Treated cells showed increased in enolase activity implying increase proliferation activity for repair purposes in the assaulted neuron (cyanide treated neurons). Presence of CD3 activity also indicates the presence of undifferentiated T-lymphocytes around the cells undergoing apoptosis in the cortex following treatment with cyanide.
PDF Abstract XML References Citation
Received: May 25, 2012;
Accepted: August 11, 2012;
Published: October 04, 2012
How to cite this article
O.M. Ogundele, J. Madukwe, O.A. Omotosho, O. Adeeyo, C.O. Akintayo and E.A. Caxton-Martins, 2012. Immunohistochemical Localization of Neuron Specific Enolase and CD3 Lymphocyte Activation. Journal of Medical Sciences, 12: 92-98.
DOI: 10.3923/jms.2012.92.98
URL: https://scialert.net/abstract/?doi=jms.2012.92.98
DOI: 10.3923/jms.2012.92.98
URL: https://scialert.net/abstract/?doi=jms.2012.92.98
INTRODUCTION
Neuron Specific Enolase (NSE) is present in high concentrations in neuronal cell clusters both in the Central Nervous System (CNS) and neuroendocrine system. The immunohistochemical technique is specific as anti-NSE antibodies stain cells localized only in the nervous system (Kaiser et al., 1989; Abdel-Wahab and Metwally, 2011). The NSE is an enzyme of the glycolytic pathway that catalyzes the conversion of phosphopglyceric acid into phosphopyruvic acid (Asa et al., 1984). During cyanide toxicity, the neurons of the different regions of the brain will respond to the assault differently based on the different cytotoxic pathways found in these regions. However, in order to account for the roles of neuronal connection in the concept of neuronal vulnerability, we used cortical tissue slices sub-cultured in Accessory Cerebrospinal Fluid (ACSF) treated with potassium cyanide (KCN) in vitro at 37°C.
A major process during cyanide toxicity is disruption of glucose metabolism, NSE being an acid soluble enzyme for the pathway that metabolizes glucose is a potent indicator of the change in metabolic activity in these cells both in vivo and in vitro (Silverman, 1991). Neuron undergoes certain sequence for morphological changes following cyanide assault. It is believed that neurons under assault shift the activity levels of NSE such that it assumes the growth levels as if differentiation was occurring thus increasing the level of this enzyme above the regular adult level under this condition (Zhou et al., 2011). It is also possible that this will occur during DNA fragmentation to increase synthesis leading to repair of the genetic material as a definitive response of the cell to aid survival by producing enough ATP for housekeeping jobs of the neurons (Ersahin et al., 2011). The purpose of this study was to examine whether changes in NSE levels corresponds with the survival of the cell and to describe a correlation pattern between elevated NSE levels and the activity of non specific lymphocytes in the brain (CD3) indicating regions of active apoptotic cells in cyanide treated neural tissues.
MATERIALS AND METHODS
All reagents were procured from Sigma Aldrich, Na+/K+ pump blockers were procured from Standard Pharma, Nigeria).
Preparation of ACSF: The ACSF was prepared so that it contains 18 mM glucose; 119 mM NaCl, 2.5 mM KCl, 1.3 mM MgSO4, 2.5 mM CaCl2, 26.2 mM NaHCO3, 1 mM NaH2PO4. The solution was prepared in distilled water to make up to 100 mL (Khakhalin and Aizenman, 2012). Oxygen was not included in the perfusion set up as we seek to study the activity of the pump blockers in cyanogenic-oxygen deprived environment.
Na+/K+ pump blockers for excitotoxicity: Digoxin and Promethazine were procured from Standard Biochem Co, Nigeria. Two hundred and fifty micrograms of powdered digoxin was dissolved in 50 mL dextrose saline solution (Kanno et al., 2011). Two milliliter ampoule of promethazine contains 50 mg promethazine in dextrose saline; the solution was diluted by making up to 50 mL in a clean glass measuring cylinder such that the new concentration is 1 mg mL-1 (Adolph et al., 2012).
Preparation of tissue for perfusion: Adult male wistar rat weighing 250 g was dissected to expose the brain using the method of Svendsen and Hau (1994). Cortical tissue blocks of 0.5 g were obtained and washed in dextrose saline and was immediately transferred into a test tubes. Six tubes labeled A, B, C, D and E such that A contains ACSF only control), B (ACSF+KCN), C1 (ACSF+KCN+digoxin), D (ACSF+promethazine), E (ACSF+digoxin). KCN was added to the perfusion fluid at a concentration of 25 mg kg-1 of tissue, digoxin concentration in ACSF was 0.3571 μg kg-1 while Promethazine concentration was 0.714 mg kg-1. Test tube C was re-perfused with the blockers every 30 min while the overall process lasted for 180 min such that the total volume of blocker perfused is between 150-250 μm.
Perfusion: The test tubes A, B, C1, C2, D and E were connected to a perfusion set up to run the respective ACSF (as described above for each tube) through the tube for 180 min, the set up was rocked gently at intervals to aid circulation and exchange in the tissue block. 10 μL metronidazole was added to each tube as an antibiotic to prevent bacteria action. The set up was incubated at 37°C.
Immunohistochemistry (antigen retrieval method): The tissues obtained were prefixed in formolcalcium to arrest all protein activities, after 48 h the tissues were then processed with embedding done at 50°C to obtain paraffin wax sections. The sections were mounted on a slide and dewaxed to expose the tissues. The exposed tissues were placed in the humidity chamber to rehydrate the tissue. It was then placed in a microwave using the method of Petersen et al. (2012).
DAB immunohistochemistry was carried out using the IgG peroxidase anti-peroxidase method (PAP). The sections were washed in Phosphate Buffer Saline (PBS), incubated in normal goat serum for 30 min and then incubated for 6 h at 37°C in a solution containing a monoclonal antibody against rat Bax, Cathepsin D and p53 proteins. Separate slides were incubated with at a dilution of 1:100 in PBS in separate reaction chambers. The sections were then treated with trypsin (0.1% in 0.1% calcium chloride) at 37°C for 15 min before blocking the non-specific reactions with hydrogen peroxide (0.3%). The antibodies were used at a dilution of 1:100 in PBS containing 0.1% bovine serum albumin and 0.005% sodium azide. The peroxidase in all the reactions was developed with 0.025% di aminobenzidine tetrachloride in phosphate buffer saline, pH 7.2, containing 0.03% hydrogen peroxide.
Transformation: Methenamine silver intensification was used on the immunoperoxidase preparation after the peroxidase/H2O2/DAB reaction has been carried out to give a brown deposit. The sections were then counterstained in hematoxylin. The counterstained sections were washed in running tap water, thoroughly rinsed in distilled water and placed in preheated methenamine silver solution at 60°C for 5 min. Although it could be occasionally longer if the intensification had been carried out at room temperature. In this study, to further increase the clarity, hematoxylin was removed from counterstained nuclei with acid alcohol before the silver intensification was carried out.
The composition of the stock solution was 0.125% silver nitrate in 1.5% hexamine. The solution was stored at 4°C. Prior to use, 2 mL of 5% tetraborate was added to 50 mL of the stock silver solution giving a pH of 8.0 which was then filtered into a coupling jar and protected from sunlight.
RESULTS
Both CD3 and NSE were expressed in the tissue rather than in the cells of the control group (Fig. 1a and b ACSF). This shows that metabolism and constant repair is a regular process in the neuronal cells. On comparison with the tissue treated with KCN, the CD3 activity occurred over a wider area as shown by arrow heads in (Fig. 2a, b: ACSF+KCN) and NSE was localized in the cells specifically in the centre of the cell (labeled*) thus indicating increased activity in the cells under cyanide toxicity. The CD3+ distribution also shows apoptotic activity occurring in a wide region in the tissue compared to specific point location in the control. CD3+ activities were localized around neurons with vacuolar spaces.
 | |
| Fig. 1(a-b): | Immunohistochemistry of tissues perfussed with ASCF only (control). Arrow heads show the space around neurons as a form of response to stress induced by oxygen deprivation, (*) indicates the sites of (a) CD3+ and (b) NSE activity, Magnification X400 |
This also support the idea that NSE activity will increase in response to repair or proliferation. The highest CD3 activity was observed in tissues treated with ACSF+KCN+Methyldigoxin (Na/K ATPase pump blocker) showing rapidly processing degenerative cells (Fig. 3a, b). Although in this tissue the NSE level was greatly reduced showing that the pump blockers compliment the toxicity of cyanide by reducing basal metabolic response in the cell and the ability of the neuron to produce ATP for recovery.
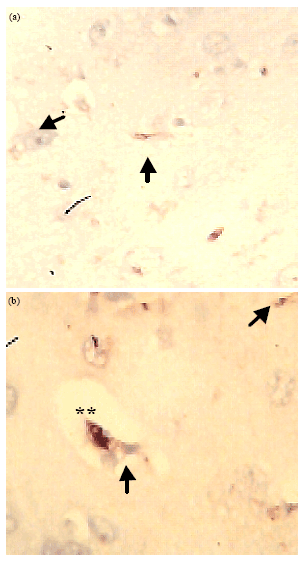 | |
| Fig. 2(a-b): | Immunohistochemical demonstration of (a) CD3+ and (b) NSE activity in tissues perfused with ACSF+KCN, (**) indicates centre of neuron, Arrow heads indicate the vacuolar spaces, Magnification X400 |
Higher NSE activity was seen in the neurons of ACSF+promethazine (another form of Na/K ATPase pump blocker) treated tissue sections. NSE activity was highest in this group due to excitotoxic effects of promethazine and the extent of the activity was observed to extends along the axonal length (arrow head in Fig. 4a and b: ACSF+promethazine). In this tissue CD3+ activity was restricted to regions around degenerating cells. This shows that the distinct effect of the pump blocker is to create excitation in the cell leading to cell enlargement and increased NSE due to increased metabolism as a result of over excitation demanding energy rather than as a result of DNA cleavage causing increased NSE activity for repair.
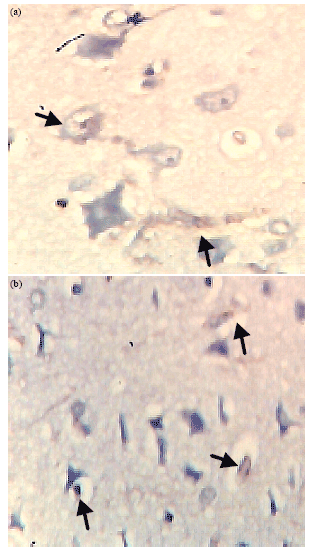 | |
| Fig. 3(a-b): | Immunohistochemical demonstration of (a) CD3+, Arrow heads indicate rapidly progressing degenerative cells and (b) NSE (arrow heads) activity in tissue treated with ACSF+KCN+methyldigoxin, magnification X400 |
While CD3+ activity was higher in ACSF+mehtyldigoxin treated tissue than promethazine treated one (Fig. 5a). However, NSE activity (Fig. 5b) in ACSF+mehtyldigoxin treated tissue was greatly reduced and present in small degenerating neurons only.
DISCUSSION
Immunohistochemical demonstration of NSE showed proliferation of neurons and axons in assaulted cells (Einstein et al., 2006). The disease related roles of NSE has been attributed more to its ability to bind to genetic materials rather than functioning as an enzyme.
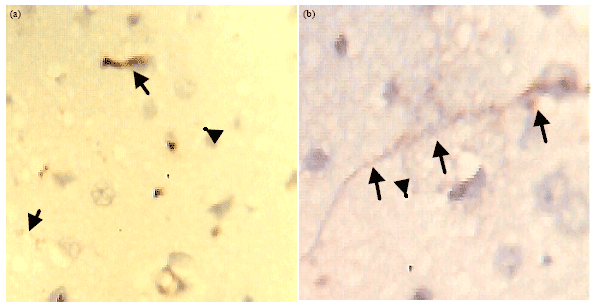 | |
| Fig. 4(a-b): | Immunohistochemical demonstration of (a) CD3+ and (b) NSE activity in tissue treated with ACSF+promethazine, arrow heads indicate CD3+ and NSE activity as the NSE activity extends long the length of the axon and cell body of neurons (arrow head in Fig. 4b). The CD3+ activity were restricted to regions around degenerating cells (arrow head in Fig. 4a), Magnification X400 |
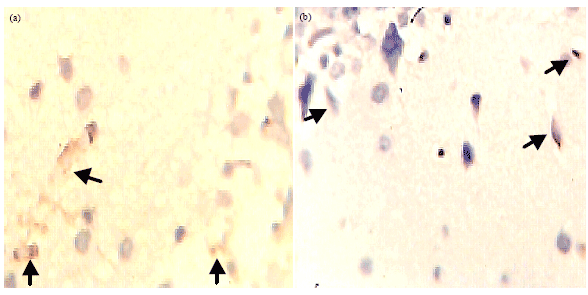 | |
| Fig. 5(a-b): | Comparative immunohistochemisty of (a) CD3+ and (b) NSE activity in cortical tissues treated with ACSF+methyldigoxin, arrow heads indicate CD3+ and NSE activities, Magnification X400 |
This might account for the location of this enzyme in the nucleus of treated neurons as shown in Fig. 1 (ACSF+KCN). This ability of the enzyme to bind genetic materials will always affect its three dimensional surface structure (Mukaratirwa et al., 2012).
The isozymes of NSE identified in the nervous system are the γ-sub-unit localized in the cell body and axons. NSE levels have also been used to determine tumorgenensis in the body (serum level). The origin of the NSE or rather the cause of the rise in NSE level is of great concern. Recently studies on blood parameters shows that rise in NSE levels may be as a result of activation of macrophages and lymphocytes (Sanchez-Ruiz et al., 2008). Activation of the T-lymphocytes (CD3) a form of macrophage can occur in the nervous system. In this study, the significance of the association between NSE and CD3 supports the fact that macrophage activity promotes activity of NSE (Forooghian et al., 2007). In nerve tissues, the NSE is used as an indicator to reflect the extent of damage to the neurons (Sanchez-Ruiz et al., 2008). NSE can also be assayed in the blood using radio immunoassay, this is because the platelets also contains level of this enzyme similar to those seen in the brain and 10-folds those seen in the erythrocyte (Jang et al., 2012). Negative correlations observed between NSE activity and platelet count may, partly, reflect platelet destruction due to pronounced coagulopathy (Zhang et al., 2011).
Reactive lymphocytes stain strongly for CD3 and was found to increase in neuronal cells tissues having cells with features of apoptosis. Comparing the activity of CD3+ and NSE for Fig. 4 and 5 (ACSF+promethazine and ACSF+methyldigoxin suggests that methyldigoxin blockade of the receptor gave more neurotoxic effect by completely reducing NSE activity while T-lymphocytes were activated around the degenerating cells (Fig. 4a, b). In the ACSF+promethazine treated tissues, excitotoxicity is a possible effect of this blocker as NSE activity was distributed along the length of the axon and CD3+ restricted to near surroundings of the cells.
To answer the primary question as to the role and source of NSE in cyanide toxicity; NSE in this study is believed to be of 2 sources, firstly the NSE activity observed in the intercellular spaces and sites of CD3 activity are as a result of activation of the T-lymphocytes specifically undifferentiated lymphocytes found in the brain and indicated by CD3 staining. Secondly, the NSE localized in the cells is an indication of increased proliferative metabolism in the neurons. This can be described as a form of neuron defense mechanism to counter the toxicity of cyanide. Further research can be conducted to analyze and compare the activity of NSE in peripheral nervous system to compare between the behavior of the neuron types in the CNS and the peripheral nervous system under oxygen deprivation and its role in cell survival in toxicity.
REFERENCES
- Abdel-Wahab, B.A. and M.E. Metwally, 2011. Ginkgo biloba enhances the anticonvulsant and neuroprotective effects of sodium valproate against kainic acid-induced seizures in mice. J. Pharmacol. Toxicol., 6: 679-690.
Direct Link - Adolph, O., S. Koster, M. Georgieff, E.M. Georgieff, W. Moulig and K.J. Fohr, 2012. Promethazine inhibits NMDA-induced currents new pharmacological aspects of an old drug. Neuropharmacology.
CrossRef - Asa, S.L., N. Ryan, K. Kovacs, W. Singer and P.J. Marangos, 1984. Immunohistochemical localization of neuron-specific enolase in the human hypophysis and pituitary adenomas. Arch. Pathol. Lab. Med., 108: 40-43.
PubMed - Einstein, O., N. Grigoriadis, R. Mizrachi-Kol, E. Reinhartz and E. Polyzoidou et al., 2006. Transplanted neural precursor cells reduce brain inflammation to attenuate chronic experimental autoimmune encephalomyelitis. Exp. Neurol., 198: 275-284.
PubMed - Ersahın, M., H.Z. Toklu, C. Erzık, D. Akakin, S. Tetık, G. Sener and B.C. Yegen, 2011. Ghrelin alleviates spinal cord injury in rats via its anti-inflammatory effects. Turk. Neurosurg., 21: 599-605.
PubMed - Forooghian, F., R.K. Cheung, W.C. Smith, P. O'Connor and H.M. Dosch, 2007. Enolase and arrestin are novel nonmyelin autoantigens in multiple sclerosis. J. Clin. Immunol., 27: 388-396.
PubMed - Jang, B., Y.C. Jeon, J.K. Choi, M. Park and J.I. Kim et al., 2012. Peptidylarginine deiminase modulates the physiological roles of enolase via citrullination: links between altered multifunction of enolase and neurodegenerative diseases. Biochem. J.
Direct Link - Kaiser, E., R. Kuzmits, P. Pregant, O. Burghuber and W. Worofka, 1989. Clinical biochemistry of neuron specific enolase. Clin. Chim. Acta, 183: 13-31.
CrossRef - Kanno, S., K. Watanabe, I. Yamagishi, S. Hirano, K. Minakata, K. Gonmori and O. Suzuki, 2011. Simultaneous analysis of cardiac glycosides in blood and urine by thermoresponsive LC-MS-MS. Anal. Bioanal. Chem., 399: 1141-1149.
Direct Link - Khakhalin, A.S. and C.D. Aizenman, 2012. GABAergic transmission and chloride equilibrium potential are not modulated by pyruvate in the developing optic tectum of xenopus laevis tadpoles. PLoS One, Vol. 7.
Direct Link - Mukaratirwa, S., P. Rogerson, A.L. Blanco, S.W. Naylor and A. Bradley, 2012. Spontaneous cerebellar primitive neuroectodermal tumor in a juvenile cynomolgus monkey (Macaca fascicularis). Toxicol. Pathol.
CrossRef - Petersen, M., L.M. Davids and S.H. Kidson, 2012. Simultaneous immunofluorescent labeling using anti-BrdU monoclonal antibody and a melanocyte-specific marker in formalin-fixed paraffin-embedded human skin samples. Appl. Immunohistochem. Mol. Morphol.
Direct Link - Sanchez-Ruiz, M., L. Wilden, W. Muller, W. Stenzel and A. Brunn et al., 2008. Molecular mimicry between neurons and an intracerebral pathogen induces a CD8 T cell-mediated autoimmune disease. J. Immunol., 180: 8421-8433.
Direct Link - Zhang, Q.H., Q. Chen, J.R. Kang, C. Liu and N. Dong et al., 2011. Treatment with gelsolin reduces brain inflammation and apoptotic signaling in mice following thermal injury. J. Neuroinflamm., Vol. 21.
CrossRef - Zhou, J.J., Y. Xie, Y. Zhao and Z.X. Li, 2011. Neuron specific enolase gene silencing suppresses proliferation and promotes apoptosis of lung cancer cells in vitro. Nan Fang Yi Ke Da Xue Xue Bao., 31: 1336-1340.
PubMed