
Mona Sharaf
Department of Protein Researches, Genetic Engineering and Biotech Institute, City for Scientific Research, New Borg Elarab, 21934 Alexandria, Egypt
Nehal M. El-Deeb
Department of Medical Biotechnology, Genetic Engineering and Biotech Institute, City for Scientific Research, New Borg Elarab, 21934 Alexandria, Egypt
Hala I. EL-Adawi
Department of Medical Biotechnology, Genetic Engineering and Biotech Institute, City for Scientific Research, New Borg Elarab, 21934 Alexandria, Egypt
Journal of Medical Sciences
Year: 2012 | Volume: 12 | Issue: 4 | Page No.: 107-113







ABSTRACT
Hepatitis C virus (HCV) is a major cause of chronic hepatitis, liver cirrhosis and hepatocellular carcinoma worldwide. A vaccine to prevent HCV infection has yet to be introduced and the treatment options for chronic HCV infection are limited. In this study, the effect of Grape Seed Extract (GSE) and its major constituents (Gallic acid, catechin and epicatechin) were investigated as anti- HCV by using the susceptible cell culture human hepatoma (HepG2 cell). In order to test the ability of the tested treatments to scavenge the free radicals, the Flow Cytometer assay was used on which, the scavenging HCV- induced free radical in HepG2 cell line was measured. Cytotoxicity assay was then used to determine the non-toxic concentrations to the culture cells. In order to study the inhibitory effect of GSE or its constituents, the HepG2 cells were infected with HCV and treated with GSE or one of its constituent. Inhibition of viral replication was detected by the amplification of viral RNA using the RT-PCR technique. Both the whole GSE and its constituents were considered active upon inhibiting the viral replication into HepG2 cells, as evidenced by the disappearance of the (+) and/or (-) strands of viral RNA- amplified products detected by RT-PCR (compared with the positive control). A quantitative analysis was then done by real time-qPCR on which three modes of treatments (post-, pre- and co-treatments) were carried out. The quantitative analysis by real time-qPCR revealed that, all treatments have the ability to inhibit the HCV with priority to epicatechin (76%), in addition, the potentiality to scavenge the free radicals reached up to 87.74%.
PDF Abstract XML References Citation
Received: July 07, 2012;
Accepted: September 26, 2012;
Published: December 18, 2012
How to cite this article
Mona Sharaf, Nehal M. El-Deeb and Hala I. EL-Adawi, 2012. The Potentiality of Grape Seed Extract as a Novel Anti-hepatitis C virus Agent. Journal of Medical Sciences, 12: 107-113.
DOI: 10.3923/jms.2012.107.113
URL: https://scialert.net/abstract/?doi=jms.2012.107.113
DOI: 10.3923/jms.2012.107.113
URL: https://scialert.net/abstract/?doi=jms.2012.107.113
INTRODUCTION
Hepatitis C virus (HCV) is a major cause of liver cirrhosis and hepatocellular carcinoma (Alter, 2007; Asselah et al., 2010). In 2009, the WHO report stated that, the number of people worldwide infected by HCV is estimated to be over 170 million (WHO, 2009). HCV remains endemic in many countries; Egypt reported the highest prevalence of HCV worldwide, ranging from 6% to more than 40% among different regions and demographic groups (Lehman and Wilson, 2009). There is no vaccine effective for HCV at present (Jawaid and Khuwaja, 2008). The current standard treatment for HCV is combination therapy consisting of pegylated interferon (PEG-INF) and Ribavirin. This therapy achieves 50% Sustained Virological Response (SVR) (Schvarcz et al., 1995; Reichard et al., 1998). In under developed countries, standard interferon is still the main choice for HCV therapy since the pegylated interferon is expensive. On the other hand, adverse effects with treatment are common, like hematological complications. Alternatively, new therapeutic approaches such as Direct Acting Antivirus (DAA), internal ribosome entry site (HCV IRES) inhibitors, NS3 and NS5a inhibitors, Ribavirin and interferon modified forms are still under study (Munir et al., 2010).
Since HCV infects the liver and requires two or more decades to extend into substantial disease, an alternative herbal supplement might decrease or stop disease development. More recent studies induce hope for HCV patient that is based on the Chinese medicinal herb Rhodiola kirilowii (Zuo et al., 2007). According to our previous study, the rhizomes of Zingiber officinale may also act as possible inhibitor of HCV (El-Adawi et al., 2011a).
Wine grapes’s (Vitis vinifera L.) seed extract has a broad spectrum of medicinal and therapeutic properties due to its high concentration of proanthocyanidins (Ray et al., 2001; Wang et al., 2005; Chang et al., 2006; El-Adawi et al., 2006; Abd El-Wahab et al., 2008). We previously screened qualitatively the possible inhibitory effect of Grape Seed Extract (GSE) on HCV (Abd El-Wahab and El-Adawi, 2009) and the promising result prompted us to confirm the previous work quantitatively and investigate the invasion inhibitor mechanism.
MATERIALS AND METHODS
Grape seeds extraction: The GSE was extracted and analyzed by HPLC as described in details by El-Adawi et al. (2011b).
Cell culture: Kato et al. (1996) have found that among several human hepatocyte cell lines analyzed, the hepatocellular carcinoma HepG2 cell line was found to be the most susceptible to HCV infection; therefore, HepG2 was used in this study. The procedure was carried out as described in details by El-Adawi et al. (2011a).
Cytotoxicity assay: For the determination of treatments concentration that does not exert a toxic effect on HepG2 cells, the cytotoxicity assay was carried out. A cell suspension of 6x104 cell mL-1 was collected and seeded in 96 well plates (100 μL cell suspension per well). The plates were incubated at 37°C in humidified 5% CO2 for 24 h, the exhausted old medium were discarded and 100 μL of different treatment concentrations (μg/100 μL RPMI) or medium (as a negative control) were added. The cell plates were incubated at the same growth conditions for 3 days. After 3 days, culture medium was discarded, 100 μL of neutral red stain (100 μg mL-1) was added to each well and incubated for 3 h at 37°C in humidified 5% CO2 (Borenfreund and Puerner, 1985).
Antioxidant activity of GSE and its constituents: The total intracellular Reactive Oxygen Species (ROS) generated during HCV infection was detected in HepG2 using 2',7'-dichlorofluorescein diacetate (DFCH-DA) method. DFCH-DA enters cells and further oxidized by ROS forming fluorescent product (DFC).
HepG2 cells were grown in 96 well plates in complete RPMI medium at 37°C in humidified 5% CO2 for 24 h, 100 μL of the recorded non-toxic doses of the extracts were added to cells and incubated at the previous incubation conditions for 24 h, after that, the treatments were removed and cells w ere washed again with pre-warmed PBS. Cells were incubated with DFCH-DA at a final concentration 50 μM for 30 min, after incubation, ROS was induced by loading infected serum in RPMI (50 mL of medium containing 1 mL of serum 1.09x109 copies mL-1). In the control groups, 100 μL RPMI was added. After incubation for 1 h, the stimulants were discarded and cells were washed three times with prewarmed PBS, all cell samples were analyzed using a BD FACSCalibur flow cytometer (Hafer et al., 2008) with Cell Quest software, Cells were excited at 485 nm and DCF fluorescence was read on FL1 (530 nm) in log scale with FL1 gain set to 443. The inhibition percentage of ROS production was calculated by formula:
where, F0 is fluorescence of the control sample and Ft is fluorescence of the treated sample.
Inhibition potential of the GSE on HCV: HepG2 cell culture were prepared as discussed, then aqueous extract of GSE as well as its major constituents (gallic acid, catechin and epicatechin) were added at non toxic concentrations of treatment. Positive and negative control cultures were included; the culture was allowed to be infected with 1 mL of serum (1.09x109 copies mL-1, genotype 4a) and then incubated for 90 min at 37°C, 5% CO2 and 95% humidity. The culture was then washed three times with PBS and incubated for seven days at 37°C followed by washing three times with 1 mL of PBS and further cultured for 7 days at 37°C, followed by total RNA extraction. The positive strand and its replicating form (negative strand) were detected by reverse transcriptase (RT) -PCR using HCV specific primers to the 5'-untranslated region of the virus.
RNA extraction from HepG2 cells: The (+) and /or (-) HCV-RNA strands was isolated from HepG2 as described by Lohr et al. (1995).
PCR of genomic and anti-genomic RNA strands of HCV: Reverse transcription-nested PCR was carried out according to Lohr et al. (1995) with few modifications. Retrotranscription was performed in 25 μL reaction mixture containing 20 U of AMV reverse transcriptase (Clontech, USA) with either 400 ng of total HepG2, 40 U of RNAsin (Clontech, USA), a final concentration of 0.2 mmol L-1 from each dNTP (Promega, Madison, WI, USA) and 50 pmol of the reverse primer 1 CH (for plus strand) or 50 pmol of the forward primer 2CH (for minus strand). The reaction was incubated at 42°C for 60 min and denatured at 98 for 10 min. Amplification of the highly conserved 5'-UTR sequences was done using two rounds of PCR with two pairs of nested primers. First round amplification was done in 50 μL reaction mixture, containing 50 pmol from each of 2CH forward primer and P2 reverse primer, 0.2 mmol L-1 from each dNTP, 10 μL from RT reaction mixture as template and 2 U of Taq DNA polymerase (Promega, USA) in a 1xbuffer supplied with the enzyme. The thermal cycling protocol was as follows: 1 min at 94°C, 1 min at 55°C and 1 min at 72°C for 30 cycles. The second round amplification was done similar to the first round, except for use of the nested reverse primer D2 and forward primer F2 at 50 pmol each. A fragment of 174 bp was identified in positive samples. Primer sequences were as follows: 1CH: 5'GGTGCACGGTCTACGAGACCTC-3', 2CH: 5'-AACTACTGTCTTCACGCAGAA-3', P2: 5'-TGC TCATGGTGCACGGTCTA-3', D2: 5'-ACTCGGCTAGCAGT CTCGCG-3' and F2: 5'-GTGCAGCCTCCAGGACCC-3'. To control false detection of negative-strand HCV RNA and known variations in PCR efficiency, specific control assays and rigorous standardization of the reaction were employed: (1) cDNA synthesis without RNA templates to exclude product contamination, (2) cDNA synthesis without RTase to exclude Taq polymerase RTase activity and (3) cDNA synthesis and PCR step done with only the reverse or forward primer to confirm no contamination from mixed primers. These controls were consistently negative. In addition, cDNA synthesis was carried out using only one primer present followed by heat inactivation of RTase activity at 95°C for 1 h, in an attempt to diminish false detection of negative-strand prior to the addition of the second primer.
Strand-specific RT-qPCR: The real time quantitative (RTq) PCR was done to the final PCR product based on the SYBR Green I dye and light cycler fluorimeter using a standard HCV infected serum samples (Komurian-Pradel et al., 2001). Amplicon synthesis was monitored continuously by SYBR Green I dye binding to double stranded DNA during PCR of the 5'HCV non-coding (NC) region. Specificity was verified by amplicon melting temperatures. An external standard curve was constructed with serial 10 fold dilutions of a modified synthetic HCV 5'NC RNA (22), The RT-qPCR step was followed by first round PCR using the 2CH and P2 primers in 50 μL reaction containing 20 μL cDNA, 50 pmol from each of 2CH forward primer and P2 reverse primer and 12.5 μL master mix SYBR green, The thermal cycling protocol was as follows: 30 cycles of 1 min each at 94, 55 and 72°C, with the final extension done at 72°C for 5 min.
The mode of action of the treatment: In order to confirm the mode of action of the most potent treatment, modified Acridine Orange (AO) stain methods was used to detect viral RNA content in proliferating cells according to Criswell et al. (1998) with some modifications. HepG2 cells (2x105) were allowed to grow on a sterile cover slip on 6 well tissue culture plate, cells were treated by the most potent treatment in three different manners (post-, pre- and co-treated). At the end of incubation, cells were washed twice with pre-warmed PBS. Cells on the cover slip were stained with 0.01% acridine orange in Gey’s solution for 45 sec at room temperature and washed with HBSS. The cells were then stained with 0.05% crystal violet in 0.15% NaCl for 45 sec and washed with HBSS. The slides were examined at 40x magnification, switching between fluorescent and phase contrast optics. The positive AO stain virus genome will take the bright yellowish-orange and the other cells will take the apple green.
RESULTS
Cytotoxicity assay: The cytotoxicity results (Fig. 1) indicated that, generally, GSE were safer than its constituents and have no TC50 even in the maximum concentrations 50%; followed by epicatechin, gallic acid and finally catechin that have TC50 30, 30 and 25% (μg/100 uL RPMI), respectively.
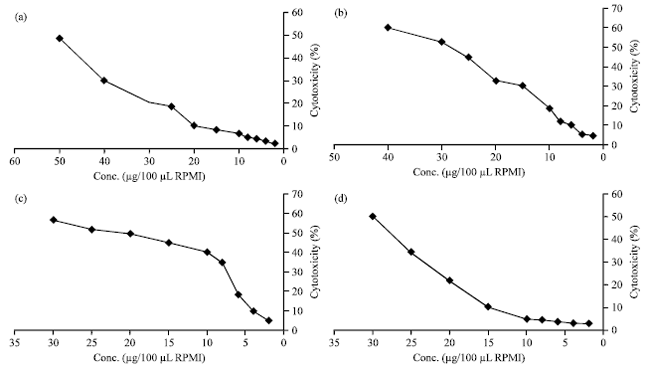 | |
| Fig. 1(a-d): | The cytotoxicity results of (a) GSE and its major constituents (b) Gallic acid, (c) Catechin and (d) Epicatechin on HepG2 cell line |
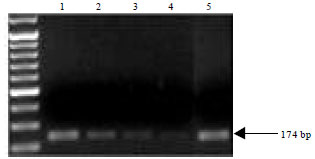 | |
| Fig. 2: | GSE and its major constituent’s activity against HCV, amplified RT- nested PCR products were resolved in 3% agarose gel/ethidium bromide staining, Lane 1: Gallic acid treatment, Lane 2: GSE treatment, Lane 3: Catechin, Lane 4: Epicatechin and Lane 5: Amplified 174 bp of HCV from used positive control |
The non toxic doses (recommended) of GSE, epicatechin, gallic acid and catechin reached up 20, 15, 6 and 4% (μg/100 uL RPMI), respectively.
The scavenging capacity of GSE and its constituents on ROS induced by HCV infection using flow cytometric analysis: Total induced ROS scavenging capacity results (Table 1) of the treatments clarified that, epicatechin was the most potent antioxidant constituent that showed a scavenging percentage reached 87.74%, followed by catechin and GSE that showed 85.35 and 62.24% scavenging percentage.
| Table 1: | Flow cytometry report of free radicals scavenging ability of GSE and its major constituents |
 | |
Finally, the gallic acid recorded the lowest percentage 50%.
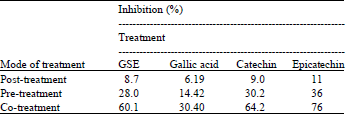
Antiviral effect of GSE and its constituents: Inhibition of viral replication was detected qualitatively by RT-nested PCR (Fig. 2) and quantitatively by amplification of the viral RNA segments using RT-qPCR technique (Table 2). The obtained results were calculated by analogy with a standard curve of different known virus concentrations in a positive control infected cells. The tested extract is considered to be active when it is capable of inhibiting the viral replication inside the HCV-infected cells, as evidenced by the inhibition of virus concentration (copies/mL) compared with positive control. Among all treatments, epicatechin showed the highest inhibition percentage (76%) where, it reduced the viral concentration from 1.09x109 to 2.4x108, while, catechin, GSE and gallic acid inhibited the HCV with a percentage 64.2, 60.1 and 30.4, respectively.
DISCUSSION
It was reported that the level of Radical Oxygen Species (ROS) as well as lipid peroxidation elevated in case of HCV infection due to the HCV core protein (Okuda et al., 2002; Waris et al., 2005). Such increase might induce fibrosis and carcinogenesis in chronic HCV (Koike, 2005). Based on these data, a rationale for antioxidant treatment of chronic hepatitis C was suggested. The difference in the ability of polyphenols to deactivate the ROS depends upon the variation in the chemical structure, position of hydroxyl groups and many other features of the plants secondary metabolties (polyphenols). In the case of GSE, the main polyphenol components are gallic acid, catechin, epicatechin (EC) and proanthocyanidin dimer B2 (EC-EC). It was shown in the results section that, both the crude extract and each main component possess a free radical scavenging activity with priority to epicatechin (87.74%). Such effect might help to reduce or protect from fibrosis and hepatocellular carcinoma by interrupting the initiation of a new ROS chain reaction.
In addition to scavenging activity of GSE, the inhibitory effect on HCV was also recorded qualitatively by RT-nested PCR (Fig. 2) and then quantitatively by RT-qPCR where the epicatechin inhibited the HCV by 76% (Table 2).
Regarding the mechanism underlying the antiviral activities (Table 2, Fig. 3), pretreatment of HepG2 cells with epicatechin before HCV inoculation moderately reduced the HCV infectivity and after HCV inoculation (post-treatment) slightly reduced the HCV infectivity.
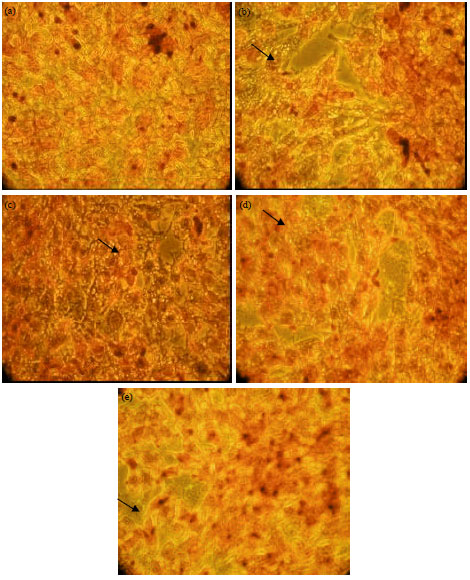 | |
| Fig. 3(a-e): | HEPG2 cell infected with HCV virus were stained by acridine orange stain after three different treatments with epicatechin, Acridine orange stain interacted with viral RNA inside infected cells giving a green fluorescent plaque, The small black arrows in the photos indicated the fluorescent plaque induced by viral RNA in (a) Negative control, (b) Positive control, (c) Post-treatment, (d) Pre-treatment and (e) Co-treatment |
| Table 2: | RT-qPCR results as inhibition of HCV due to the mode of treatment of GSE and its constituents, respectively |
 | |
On the other hand, the application of epicatechin during inoculation (co-infection) strongly inhibited HCV infectivity. These results could be interpreted at two levels: the first is at the intracellular level (viral replication) and the second is at the direct interaction with virus molecule (extracellular level).
At the intracellular level: absorption of epicatechin from the digestive tract has been reported (Schroeter et al., 2006) and may cross the blood-brain barrier (Abd El Mohsen et al., 2002) suggesting the possibility of epicatechin internalization into cells, the idea which has been proven recently by Maurya and Prakash (2011). Based on the last information, in case of pre- and post treatment, the epicatechin might bind to the translational initiation complex associated with HCV Internal Ribosome Entry Site (IRES) and thereby suppresses the HCV genome expression that means viral replication inhibition. However, the effect of natural product-derived polyphenolic compounds on HCV IRES-directed translation is poorly studied and further investigation is needed.
At the extracellular level: in case of co-infection, the epicatechin might inhibit the HCV entry through direct interaction with virus molecules rather than interaction with HepG2 cells. That explanation might interpret the strong inhibition in co-infection (low number of HCV genome inside the host cells) than both pre-and post treatment. The idea behind was epicatechin might inhibit viral attachment to the HepG2, probably by acting directly on the particle surface, thus disrupting the initial step of HCV cell entry. This entry inhibition recorded here agreed with the recent study by Ciesek et al. (2011) and Calland et al. (2012) where they have proven that epigallocatechin-3-gallate (EGCG), another polyphenol member, as a good inhibitor for HCV entry.
CONCLUSION
This study suggested that all treatments possessed anti-viral and anti-oxidative properties, which were associated with their direct free radical scavenging abilities. The GSE molecule, epicatechin has dual functions, at the extracellular level where it could inhibit the HCV entry and at the intracellular where it could interrupt the HCV replication. Further studies of the mechanism underlying epicatechin-mediated HCV inhibition may open new ways to design novel anti-HCV drugs.
ACKNOWLEDGMENT
The study has been received the financial support from City for Scientific Research and Technology Applications.
REFERENCES
- Alter, M.J., 2007. Epidemiology of hepatitis C virus infection. World J. Gastroenterol., 13: 2436-2441.
Direct Link - Borenfreund, E. and J.A. Puerner, 1985. Toxicity determined in vitro by morphological alterations and neutral red absorption. Toxicol. Lett., 24: 119-124.
CrossRefDirect Link - Chang, W.T., Z.H. Shao, T.L.V. Hoek, E. McEntee and S.R. Mehendale et al ., 2006. Cardioprotective effects of grape-seed proanthocyanidin, baicalin and wogonin: Comparison between acute and chronic treatment. Am. J. Chin. Med., 34: 363-365.
PubMed - Calland, N., A. Albecka, S. Belouzard, C. Wychowski and G. Duverlie et al., 2012. (-)-Epigallocatechin-3-gallate is a new inhibitor of hepatitis C virus entry. Hepatology, 55: 720-729.
CrossRefDirect Link - Ciesek, S., T. von Hahn, C.C. Colpitts, L.M. Schang and M. Friesland et al., 2011. The green tea polyphenol epigallocatechin-3-gallate (EGCG) inhibits hepatitis C virus (HCV) entry. Hepatology., 54: 1947-1955.
CrossRef - Criswell, K.A., G. Krishna, D. Zielinski, G.C. Urda, J.C. Theiss, P. Juneau and M.R. Bleavins, 1998. Use of acridin orange in: Flow cytomertric assessment of micronuclei induction. Mutat. Res., 414: 63-75.
CrossRef - El-Adawi, H., M. Abdel Mohsen, D. Youssef and S. El-Sewedy, 2006. Study on the effect of grape seed extract on hypercholestrolemia: Prevention and treatment. Int. J. Pharmacol., 2: 593-600.
CrossRefDirect Link - El-Adawi, H., M. El-Demellawy and A. Abd El-Wahab, 2011. Some medicinal plant extracts exhibit potency against viral hepatitis C. J. Biosci. Tech., 2: 223-231.
Direct Link - El-Adawi, H., D. El-Azhary, A. Abd El-Wahab, M. El-Shafeey and M. Abdel-Mohsen, 2011. Protective effect of milk thistle and grape seed extracts on fumonisin B1 induced hepato-and nephro-toxicity in rats. J. Med. Plants Res., 5: 6316-6327.
Direct Link - Hafer, K., K.S. Iwamoto and R.H. Schiestl, 2008. Refinement of the dichlorofluorescein assay for flow cytometric measurement of reactive oxygen species in irradiated and bystander cell populations. Radit. Res., 169: 460-468.
PubMedDirect Link - Jawaid, A. and A.K. Khuwaja, 2008. Treatment and vaccination for hepatitis C: Present and future. J. Ayub Med. Coll. Abbottabad, 20: 129-133.
PubMed - Kato, N., M. Ikeda , T. Mizutani , K. Sugiyama , M. Noguchi , S. Hirohashi and K. Shimotohno, 1996. Replication of hepatitis C virus in cultured non-neoplastic human hepatocytes. Jpn. J. Cancer Res., 87: 787-792.
PubMed - Koike, K., 2005. Molecular basis of hepatitis C virus-associated hepatocarcinogenesis: Lessons from animal model studies. Clin. Gastroenterol. Hepatol., 3: S132-S135.
CrossRefPubMedDirect Link - Lohr, H.F., B. Goergen, K.H. Meyer zum Buschenfelde and G. Gerken, 1995. HCV replication in mononuclear cells stimulates anti-HCV-secreting B cells and reflects nonresponsiveness to interferon-α. J. Med. Virol., 46: 314-320.
CrossRefPubMedDirect Link - Maurya, P.K. and S. Prakash, 2011. Intracellular uptake of (-)epicatechin by human erythrocytes as a function of human age. Phytother. Res., 25: 944-946.
CrossRefPubMedDirect Link - Munir, S., S. Saleem, M. Idrees, A. Tariq and S. Butt et al., 2010. Hepatitis C treatment: Current and future perspectives. Virol. J., Vol. 7.
CrossRefDirect Link - Okuda, M., K. Li, M.R. Beard, L.A. Showalter, F. Scholle, S.M. Lemon and S.A. Weinman, 2002. Mitochondrial injury, oxidative stress and antioxidant gene expression are induced by hepatitis C virus core protein. Gastroenterology, 122: 366-375.
CrossRefPubMedDirect Link - Ray, S.D., H. Parikh, E. Hickey, M. Bagchi and D. Bagchi, 2001. Differential effects of IH636 grape seed proanthocyanidin extract and a DNA repair modulator 4- aminobenzamide on liver microsomal cytochrome 4502E1-dependent aniline hydroxylation. Mol. Cell Biochem., 218: 27-33.
CrossRef - Schroeter, H., C. Heiss, J. Balzer, P. Kleinbongard and C.L. Keen et al., 2006. (-)-Epicatechin mediates beneficial effects of flavanol-rich cocoa on vascular function in humans. Proc. Natl. Acad. Sci., 103: 1024-1029.
CrossRefDirect Link - Schvarcz, R., Z.B. Yun, A. Sonnerborg and O. Weiland, 1995. Combined treatment with interferon alpha-2b and ribavirin for chronic hepatitis C in patients with a previous non-response or non-sustained response to interferon alone. J. Med. Virol., 46: 43-47.
PubMed - Wang, C.Z., A. Fishbein, H.H. Aung, S.R. Mehendale and W.T. Chang et al., 2005. Polyphenol contents in grape-seed extracts correlate with antipica effects in cisplatin-treated rats. J. Alter. Complement Med., 11: 1059-1065.
Direct Link - Waris, G., J. Turkson, T. Hassanein and A. Siddiqui, 2005. Hepatitis C virus (HCV) constitutively activates stat-3 via oxidative stress: role of stat-3 in HCV replication. J. Virol., 79: 1569-1580.
CrossRefDirect Link - Zuo, G., Z. Li, L. Chen and X. Xu, 2007. Activity of compounds from Chinese herbal medicine Rhodiola kirilowii (Regel) Maxim against HCV NS3 serine protease. Antiviral. Res., 76: 86-92.
PubMed