
I. M. Alhazza
Department of Zoology,
College of Science,
King Saud University,
P.O. Box 2455, Riyadh,
1 1451, Saudi Arabia
A. S.A. Haffor
Department of Zoology,
College of Science,
King Saud University,
P.O. Box 2455, Riyadh,
1 1451, Saudi Arabia
Journal of Medical Sciences
Year: 2007 | Volume: 7 | Issue: 5 | Page No.: 901-904







ABSTRACT
Heavy metal toxicity causes impairment of variety of cellular process. Cellular failure to maintain balance between the generation and elimination of reactive molecules results in tissue damage. The purpose of this study was to determine the effects of Lead (BP) toxicity on Free Radicals (FR) production. Sixteen adult rats matched with age were randomly assigned to two groups, control and experimental. The experimental group was injected (IP) 1 mg kg-1 of body weight BP daily for seven days. Mean FR in the control group was 206.13±12.39 CARR U. In BP treated group the mean (±SEM) FR rose to 324.13±12.61 CARR U. The change in FR corresponds to 57% increase in the lead treated group. Thus, BP toxicity exposure mediates mitochondria damage and the subsequent oxidative stress. Based on the results of the present study, it can be concluded that BP toxicity causes elevation of Reactive Oxygen Species (ROS) which mediate inhibition of mitochondrial respiration in rats.
PDF Abstract XML References Citation
How to cite this article
I. M. Alhazza and A. S.A. Haffor, 2007. Effects of Lead Toxicity on Free Radical Production in Rats. Journal of Medical Sciences, 7: 901-904.
DOI: 10.3923/jms.2007.901.904
URL: https://scialert.net/abstract/?doi=jms.2007.901.904
DOI: 10.3923/jms.2007.901.904
URL: https://scialert.net/abstract/?doi=jms.2007.901.904
INTRODUCTION
Industrial development and modern life style has resulted in accumulation of reactive heavy metals particulates and metals vapors in the atmosphere. The potential of oxidative stress of these gases and lead particulates as well as are responsible for initiating the oxidative destruction of biomolecules, enzymetic changes and the formation of lipid peroxidase (Haffor and Al-Ayed, 2003).
Furthermore, mitochondria are easily affected by oxygen toxicity (Ogburn et al., 1988; Yan et al., 1994; Haffor et al., 2002; Haffor, 2004). It has been shown that growth of houseflies in an atmosphere of 10% O2 markedly reduced their mean and maximum life span and increases the rate of accumulation of protein carbonyls in whole body extracts (Sohal et al., 1993) and in isolated mitochondria (Sohal and Duby, 1994). Furthermore free radicals promote formation of cross link between biomolcules (Dyer et al., 1991; Barlett and Stadman, 1997; Braun, 2000) which in turn reduce movement of molecules for normal chemical reactions.
Most studies are in agreement that mitochondria are the main source for free radicals production (Bioveris and Chance, 1973; Cutler, 1986; Dizdarglu, 1992a; Sohal et al., 1993; Barja, 1999). In addition, mitochondria DNA (mtDNA) damage occurs much faster than does nuclear DNA because mitochondrial DNA is not protected by proteins, rather it is attached to the inner mitochondrial membrane (Dizdaroglu, 1992b; Beckman and Ames, 1997). The damaged mtDNA accumulates in the cells progressively because damaged mitochondria replicate faster than undamaged mitochondria. It has not been reported whether heavy metals cellular toxicity is mediated via the formation of reactive bio-molecules. The present study was designed specifically to investigate the role of lead toxicity on free radicals production.
MATERIALS AND METHODS
Experimental design: Sixteen adult male rats, Ratus rattus, matched with age and body weight were assigned randomly to a control (C group) and an experimental group, lead (BP group). The BP group was injected (IP) 1 mL BP kg-1 BW at a concentration of 1 g L-1, daily for seven days. Following data collection mean group differences for FR were evaluated using unpaired t-test with homogenous variance.
Blood samples collection: Blood samples were collected from the retro-orbital plexus using capillary then analyzed for free radicals immediately.
Free radical determination: Free radical was measured, using the d-ROMs-2 test kits (FRAS-II, Italy) according to the manufacturer’s instructions. The test measures the levels of hydroperoxides (R-OOH) which are generated by peroxidation of biological compounds; lipid, amino acids, nucleic acids (FRAS-II, 2002). This test is based on the principle of the ability of hydrogen peroxides to generate free radicals after reacting with some transitional metals (Fe2+/Fe3+ ), according to Fenton's Reaction as follows:
| (1) |
Thus, the hydrogen peroxides of biological sample (whole blood) generate free radicals (alcoxy and peroxyl radicals) after exposure to a transitional metal (F++/Fe+++). When a correctly buffered chromogen substance (N, N-diethyl-phenylendiamine) lead to the reduction of hydrogen peroxides which in turns colored as radical cation. Color intensity was read using spectrophotometer with peak absorbance of 505 nm. In the d-ROMs test results were expressed in CARR UNITS (CARR U). One CARR U relates to 0.08 mg H2O2/100 mL (FRAS II, Italy, 2002).
RESULTS
Baseline mean (±SEM) free radicals were 206.13±14.39 and increased to 324±12.61 CARR U in the lead treated group (Table 1). The changes in FR represent 57% increase in the lead treated group in comparison with the control group (Fig. 1). The result of F-test for variance homogeneity was not significant, indicating that two groups are withdrawn from the same population and hence it is appropriate to analyze among groups mean difference using independent t-test with equal variances (Table 2). The results of unpaired t-test with equal variances showed that mean free radicals production increased significantly (p<0.05) in the lead treated group (Table 3). It is very important to realize that our measurements were based on the rate of free radicals production based on the amount of reactive H2O2.
| Table 1: | Free radicals (CARR U) data in the two groups |
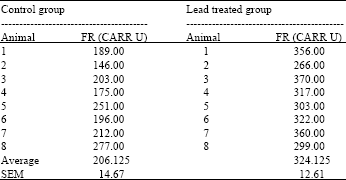 | |
| Table 2: | F test for variance equality in the two groupssis |
 | |
| Table 3: | Unpaired t-test assuming equal variance |
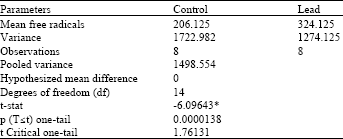 | |
| *p<0.05 (rejection value at p = 0.0000138 or less) | |
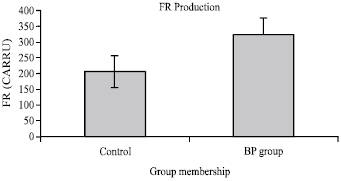 | |
| Fig. 1: | Histogram displays the mean FR in the two groups |
DISCUSSION
The major findings of the present study showed that toxicity induced by lead resulted in an increased rate of free radicals production. These results clearly demonstrated that exogenous lead toxicity result in an over production in Reactive Oxygen Species (ROS) and subsequently an accelerated free radicals production rate which overwhelmed the cell clearance rate. There is a general in agreement that mitochondria are the major oxygen radicals producing sites. Mitochondria release hydrogen peroxide to the cytosol, leading to imbalance between its generation and elimination by cellular antioxidants. It had been shown that cellular oxidative damage was related to cross links between lipid accumulation (Beuchat and Chong, 1998), protein (Barlett and Stadman, 1997; Iqbal et al., 1999; Braun, 2000) and DNA (Beckman and Ames, 1997).
Free radical determination of the present study was based on the measurements of H2O2 accumulation which reflected the plasma membrane failure to maintain osmotic balance and the subsequent cellular death. Unfortunately, most of Free radicals research had focused on the measurements of antioxidant changes rather than hydrogen peroxides as conducted in the present study which is an earlier event involved in cellular toxicity. Early comparative studies suggest that antioxidants were affected by reactive radicals which in turn influence the activities of antioxidants enzymes (Cutler, 1986; Ogburn et al., 1988; Yan et al., 1994). These authors concluded that, when tissue antioxidant were directly studied as a function of Mean Life Span (MLSP) had proved that FR production is lower in long-lived than in short-lived. Research studies on longevity studies confirmed the same conclusions (Honda et al., 1993; Ku and Sohal, 1993; Sohal et al., 1993; Holmes and Austand, 1995; Barja, 1999; Haffor et al., 2002).
From molecular standpoint, oxidative damage to nucleic acids includes adducts of base and sugar groups (Dizdarglu, 1992a, b; Epe, 1996), single-and double-links to other molecules (Halliwell and Dizarogln, 1992; Halliwell and Gutteridge, 2003). In addition exogenous hyperoxia places an oxidative stress during oxidation in the respiratory chain located in the mitochondrial cristae as evident by increasing FR production in the present study.
In conclusion exogenous heavy metal cellular toxicity resulted in an increased rate of free radicals production. Further research is needed to identify the molecular basis of heavy metals induced oxidative stress among species with different age groups.
ACKNOWLEDGMENTS
This project was supported by a Grant Number ZOO/2006/11, Research Center, College of Science, King Saud University, Riyadh, Saudi Arabia.
REFERENCES
- Barja, G., 1999. Mitochondrial free radical generation sites of production in state 4 and 3, organ specificity and relationship with aging rate. J. Bioenergy Biomembr., 31: 347-366.
CrossRefDirect Link - Berlett, B.S. and E.R. Stadtman, 1997. Protein oxidation in aging, disease and oxidative stress. J. Biol. Chem., 272: 20313-20316.
Direct Link - Beuchat, C.A. and C.R. Chong, 1998. Hyperglycemia and its consequences for hemoglobin glycation. Comp. Biochem. Physiol., 120: 409-416.
PubMedDirect Link - Epe, B., 1996. DNA damage profiles induced by oxidatizing agents. Rev. Physiol. Biochem. Pharmcol., 127: 223-249.
PubMedDirect Link - Haffor, S.A. and M.I. Al-Ayed, 2003. The effect of lead bioaccumulation on haem biosynthetic enzymes in fish. J. Environ. Biol., 24: 271-280.
PubMedDirect Link - Halliwell, B. and M. Dizdaroglu, 1992. Commentary the measurement of oxidative damage to DNA by HPLC and GC/MS techniques. Free Radic. Res. Commun., 16: 75-87.
CrossRefDirect Link - Sohal, R.S. and A. Dubey, 1994. Mitochondrial oxidative damage, hydrogen peroxide release and aging. Free Radic. Biol. Med., 16: 621-626.
PubMed