
Wilson Abrao Saad
Basic Institute of Bioscience, University of Taubate, Unitau, Taubate, SP, Brazil Centro Universitario de Araraquarsa, Araraquara SP, Uniara
Department of Physiology and Pathology, School of Dentistry,
Paulista State University, UNESP Araraquara, SP Brazil
Ismael Francisco Motta Sigueira Guarda
Department of Anesthesiology, Clinic Hospital State of Sao Paulo,
Sao Paulo, Brazil
Luis Antonio de Arruda Camargo
Department of Physiology and Pathology, School of Dentistry,
Paulista State University, UNESP Araraquara, SP Brazil
Talmir Augusto Faria Brizola dos Santos
Basic Institute of Bioscience, University of Taubate, Unitau, Taubate, SP, Brazil
William Abrao Saad
Department of Gastroenterology, School of Medicine,
University of S�o Paulo, Brazil
Journal of Medical Sciences
Year: 2007 | Volume: 7 | Issue: 5 | Page No.: 845-849







ABSTRACT
We determined the effects of AT1 and AT2 (selective no peptides antagonists angiotensin receptors), arginine vasopressin V1 receptor antagonist as well as L-arginine, a nitric oxide donor and NW-nitro-L-arginine methyl ester (L-NAME), a nitric oxide synthase inhibitor, injected into supraoptic nucleus (SON) on water and sodium intake induced by the injection of angiotensin II (ANGII). Male Holtzman rats weighing 200-250 g with canulae implanted into the SON were used. The drugs were injected in 0.5 μL over 30-60 sec. The water intake after injection of saline SAL+SAL 0.15 M NaCl was 0.40±0.1 mL 2 h-1; SAL+ANGII increase water intake. Losartan decreased the water intake induced by ANGII. PD123319 injected prior to produce no change in water intake induced by ANGII. AVPA prior to ANGII reduced the water intake with a less intensity than losartan. L-arginine prior to ANGII decreases the water intake at a same intensity than losartan. L-NAME prior to ANGII potentiated the dipsogenic effect of ANGII. Losartan injected simultaneously with L-arginine prior to ANGII blocked the dipsogenic effect of ANGII. These results confirm the importance of SON in the control of water intake and strongly suggest that AT1, V1 receptors interact with nitrergic pathways within the SON influencing the dipsogenic effect of ANGII.
PDF Abstract XML References Citation
How to cite this article
Wilson Abrao Saad, Ismael Francisco Motta Sigueira Guarda, Luis Antonio de Arruda Camargo, Talmir Augusto Faria Brizola dos Santos and William Abrao Saad, 2007. Effects of Nitric Oxide and Arginine Vasopressin on Water Intake Induced by Central Angiotensin II. Part 1. Journal of Medical Sciences, 7: 845-849.
DOI: 10.3923/jms.2007.845.849
URL: https://scialert.net/abstract/?doi=jms.2007.845.849
DOI: 10.3923/jms.2007.845.849
URL: https://scialert.net/abstract/?doi=jms.2007.845.849
INTRODUCTION
Central injection of angiotensin II (ANGII) elicits prompt and pronounced responses such as increased blood pressure, thirst, sodium appetite and the release of vasopressin (Saad et al., 2004a, b). Influence of vasopressin receptor and nitric oxide on the water, sodium intake and arterial blood pressure induced by angiotensin injected into the third ventricle of the rat brain has been demonstrated (Saad et al., 2006a).
NO syntethase inhibitors have been used widely to determine the role of endogenous No. Nw-nitro-L-arginine methyl ester (L-NAME) reduces renal blood flow, urine flow rate and urinary sodium excretion (Naees et al., 1992). Several studies have showed that NO may function as a neurotransmitter or a neuromodulator. Recognition of the role of nitric oxide in cell-to-cell communication has changed the concept of traditional neurotransmission. N-methyl-D-aspartate receptors mediate the dipsogenic response and c-Fos expression induced by intracerebroventricular (icv) infusion of angiotensin II (ANGII) (Zhu and Herbert, 1997). It has been demonstrated that NO may facilitate the release of excitatory transmitters, possibly through a presynaptic cyclic GMP-dependent mechanism (Wu et al., 1997). The influence of NO on angiotensin effects has been demonstrated (Saad et al., 2002).
The systemic application of ibersartan and losartan abolished the ANGII central physiologic responses (water intake, sodium intake and increase arterial pressure (Camargo and Saad, 1999). Nitric oxide and AT1-AT2 receptors influence the pressor effect of angiotensin II (Saad et al., 2006b). The role of renin-angiotensin system in the control of arterial blood pressure and salt appetite in rats has been demonstrated (Thunhorst and Johnson, 1994). Endothelial and neuronal nitric oxide synthase inhibitors influence angiotensin II pressor effect in central nervous system (Saad et al., 2006c).
Since the SON is involved in the control of hydromineral balance we investigated whether the dipsogenic response induced by the injection of ANGII into the SON could be mediated by angiotensin, vasopressin receptors and nitric oxide.
MATERIALS AND METHODS
Holtzman rats weighing 200-250 g with canulae implanted unilaterally into SON were used. The animals were housed in individual metabolic cages. Food (Purina Rat Chow) and tap water is available, ad libitum, for the duration of the experiments. The room temperature was maintained at 22±2°C. The light cycle was held at 12:12 with lights on 06:00 h. All experiments were conducted during the light period, between 09:00 am and 03:00 pm. All experiments were conducted during the light period, between 09:00 am and 03:00 pm. If it started 9 am it ended 11:00 pm.
The animal were anesthetized with ketamine (80 mg kg-1 of body weight) plus xylazine (7 mg kg-1 of body weight) intraperitoneally (ip) and implanted with 10 and 12 mm long and 0.7 mm OD stainless steel cannulae into the SON, according to the coordinates of the Paxinos and Watson (1986) rat brain atlas. The coordinates were 1.4-1.8 mm caudal to bregma, 2.3 mm lateral to middle line and 9.2 mm bellow the duramater. The cannulae were fixed to the skull with the aid of jeweler screws and dental acrylic resin and protected with a stylet. Rats recovered from surgery for a minimum of 5 days beginning of testing.
Injections into SON were made using 10 μL Hamilton syringes connected by polyethylene tubing (PE-10) to 30-gauge injection cannulas. At the time of testing, the obturator was removed and the injection cannula introduced into the chronically implanted guide cannula. The injection cannula was 0.5 mm longer than the guide cannula. The injection volume was 0.5 μL delivered over 20 to 30 sec. After injection, the styles were replaced and the rats were placed back into the cage.
ANGII purchased from Sigma (Chemical Co., St. Louis, MO) and dissolved in saline (0.15 M NaCl) at 10 nmol/0.5 μL.
PD123319 and losartan purchased from DuPont, Merck, Wilmington, DE USA and dissolved in saline (0.15 M NaCl), at 80 nmol/0.5 μL. d(CH2)5-Tyr(Me)-AVP (AVPA) purchased from Bachem, Inc., Torrance, Ca, USA and dissolved in saline (0.15 M NaCl), at 80 nmol/0.5 μL.
L-arginine purchased from Sigma (Chemical Co., St. Louis, MO) dissolved in saline (0.15 M NaCl) at 20 μg/0.5 μL.
NG-nitro-L-arginine methyl ester (L-NAME) purchased from Sigma (Chemical Co., St. Louis, MO), dissolved in saline (0.15 M NaCl) at 40 μg/0.5 μL. ANGII (10 pmol/0.5 μL) or vehicle was injected into the SON, water was offered. Each antagonist was also injected into the SON at the dose of 80 nmol/0.5 μL, 15 min before water was offered. The antagonists were injected into the SON 10 min before ANGII was injected into the SON.
L-arginine (20 μg/0.5 μL) nitric oxide donor and L-NAME (40 μg/0.5 μL) nitric oxide inhibitor were injected into SON 10 min prior to ANGII injection into the SON. Water or sodium intake was recorded each 30 min over a 2 h period using individual metabolic cages.
At the end of the experiments, the rats were anesthetized with ether and given at 0.5 μL injection of fast green dye via the intracranial cannula, followed by perfusion with saline and buffered formalin.
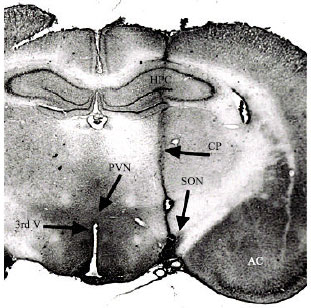 | |
| Fig. 1: | Photomicrography of a hematoxylin-stained transverse section showing the site of injection into the SON (Arrow). CP cannulae pathways; AC anterior commissure; HPC hippocampus; PVN paraventricular nucleus; 3rdV third ventricle |
The brains were removed, fixed in 10% formalin, frozen to -25°C and cut into 20-30 μm coronal sections and cut into 20-30 μm coronal sections and stained with hemathoxilin-eosin. Only animals in which the injection was placed in the lateral medial and caudal portion of the SON were use in this study Fig. 1.
Results are reported as means±standard error of the mean (SEM) for the indicated experiments. Statistical analysis was subjected ANOVA followed by the Newman-Keuls post-hoc test. Differences were considered significant at p<0.05.
RESULTS AND DISCUSSION
Water intake after injection of 0.15 M NaCl into SON was 0.4±0.1 mL 2 h-1. Injection of ANGII into SON increased water intake (10.6±1.2 mL 2 h-1) (p<0.05). Losartan injected into the SON before ANGII injection decreased water intake (2.5±0.4 mL 2 h-1) (p<0.05). AVPA injected into the SON decreased water intake induced by ANGII injection (4.5±0.5 mL 2 h-1) (p<0.05). L-arginine injected into SON previous to ANGII decreased water intake (2.1±0.2 mL 2 h-1) (p<0.05). L-NAME potentiated the effect of ANGII on these responses (15.1±0.7 mL 2 h-1) (p<0.05). PD 123319 injected into SON previous to ANGII injection produced no changes in water intake (11.3±1.3 mL 2 h-1).
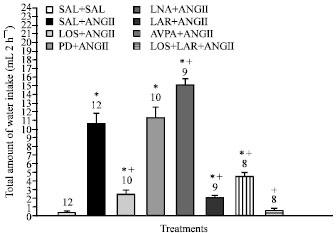 | |
| Fig. 2: | Effect of pretreatment with losartan (LOS), PD123319 (PD), d(CH2)5-Tyr(Me)-AVP (Antagonist of -AVP) (AVPA), L-arginine (LAR, L-NAME (LNA) and Losartan+L-arginine or vehicle (saline) into the SON on water intake evoked by injection of ANGII into the SON. The number of animals is represented at the top of each column. Data are reported as mean±SEM. *p<0.05 compared to the saline group; +p<0.05 compared to the ANGII group (Newman-Keuls post-hoc test) |
Losartan plus L-arginine injected into SON previous to ANGII abolished the dipsogenic effect of ANGII (0.6±0.2 mL 2 h-1) (p<0.05) Fig. 2.
The present results show that whereas injection of the selective angiotensin II AT1 antagonist losartan into the SON reduced water intake the injection of PD123319, a selective antagonist for AT2 receptors, had no effect on water intake. These findings are supported by the results of McKinley et al. (2003) and Saad et al. (2004a), showing that angiotensin AT1, AT2 and AT4 receptors are also plentiful in the brain. AT1 receptors are found in several brain regions such as the hypothalamic paraventricular and supraoptic nucleus, the lamina terminalis, lateral parabrachial nucleus, ventro lateral medulla and nucleus of the solitary tract, which are know to have roles in the regulation of the cardiovascular system and/or body fluid and electrolyte balance.
In addition, they also show that the previous injection of d (CH2)5-Tyr (Me)-AVP, an arginine vasopressin V1 receptor antagonist, injected into the SON decreased the ANGII-induced water intake. These results indicated that the water intake induced by ANGII also involves the V1 arginine vasopressin receptors.
Quantitative autoradiography with selective ANGII-receptor antagonists revealed that the SON of the rat contains AT1 receptors (Tsutsumi and Saavedra, 1991).
An endogenous origin for ANGII is suggested by various reports of angiotensin-like immunoreactivity in the magnocellular neurons of the SON (Renaud and Bourque, 1991). It is thus possible that ANGII is released locally from axon collaterals or somato-dendritic sites in a manner similar to that proposed for oxytocin or vasopressin (Richard et al., 1991). ANGII-induced water intake can be explained at least in part by working antagonistically with the ANP content and presumed release from SON, medial basal hypothalamus, anterior ventricle third ventricle (AV3V) and median eminence. The increased release of ANP from axons of neurons terminating on the effector neurons of the drinking response by stimulation of ANP receptors would inhibit the stimulatory response evoked by the action of ANGII on its receptors on these same effector neurons (De Angelis et al., 1966). The present results also show that L-NAME increased the water intake induced by thirst stimuli. It has been demonstrated that icv injection of NG-monomethyl-L-arginine (NMMA), a blocker of NOS, attenuates water intake (Kadekaro et al., 1994). Kadekaro (2004) have been demonstrated that nitric oxide has a modulation role on the hypothalamo-neurohypophyseal system.
Water intake increased after sc administration of hypertonic saline and the L-NAME previously injected into the lateral ventricle (LV) progressively reduced this ingestion (Saad et al., 1999). Electrolytic lesion of lateral preoptic area (LPO) and subfornical organ (SFO) in the same animal abolished the daily water intake induced by hypertonic saline (Saad and Camargo 1980). Previous injection of L-NAME into the SFO produced a progressive reduction in water intake induced by ANGII. These results agree with those of Liu et al. (1998). Drinking behavior induced by sc injection of hypertonic saline or by ANGII injection into the LV is attenuated by central inhibition of nitric oxide synthase with L-NAME and that the highest dose of L-NAME abolished the water intake induced by cellular dehydration and by ANGII injected into the LV (Saad et al., 1999). These results clearly demonstrate that ANGII implicatd NO to produce the dipsogenic effect and that the SON is an important area for this behavior. It has been demonstrated that NO attenuated the ANGII-induced water intake.
Also other suggestion is that, whereas the AT1 receptors of the SON mediate water ingestion induced by angiotensinergic activation, the arginine vasopressin V1 AVPergic neurons activate water intake. The AT2 receptor antagonist receptor produced no effect on water intake.
SON is an important structure of the central nervous system that regulates the dipsogenic, natriorexigenic, diurese, natriurese and pressor response induced by activation of the renin-angiotensin system. Thus the neural pathways that inhibit vasopressin release in response to an increase in blood pressure and an increase in blood volume may overlap at the perinuclear zone of SON (Cunningham et al., 2002). NO contributes to the fine regulation of water intake and vasopressin synthesis and release (Vacher et al., 2003). This affirmative is confirmed by the experiments utilizing both combination of losartan and L-arginine which abolished the ANGII effects.
The major findings of this study was that angiotensin of the SON interact with arginine vasopressin and nitric oxide to control water metabolism by controlling water ingestion.
In conclusion, these findings confirm that SON is an important central structure in controlling water metabolism such as others structures. Also angiotensinergic neural pathways and angiotensin peptides of the SON are important in neural function and may have important homeostatic roles, particularly related to water intake interacting with nitrergic and vasopressinergic pathways. These findings are strongly supported by that of Yu et al. (2006) showing that histochemical investigations indicated that the oxytocin (OXY), the arginine vasopressin (AVP) and the nitric oxide synthase (NOS) have been synthesized in the SON neurons.
ACKNOWLEDGMENTS
Research supported by CNPq, FAPESP, FUNDUNESP PRONEX and FUNADESP UNIARA.
REFERENCES
- Camargo, L.A.A. and W.A. Saad, 1999. Renal effects of angiotensin II receptor subtype 1 and 2-selective ligands injected into the paraventricular nucleus of conscious rats. Regul. Peptides, 84: 91-96.
Direct Link - Cunningham, J.T., S.B. Bruno, R.R. Grindstaff, K.H. Higgs, D. Mazzela and M.J. Sullivan, 2002. Cardiovascular regulation of supraoptic vasopressin neurons. Prog. Brain Res., 139: 257-273.
Direct Link - Kadekaro, M., 2004. Nitric oxide modulation of the hypothalamo-neurohypophyseal system. Braz. J. Med. Biol. Res., 37: 441-450.
CrossRefDirect Link - Liu, H., M.L. Terrell, V. Bui and J.Y.S. Long and M. Kadekaro, 1998. Nitric oxide control of drinking vasopressin and oxitocin release and blood pressure in dehydrated rats. Physiol. Beh., 63: 763-7569.
Direct Link - McKinley, M.J., A.L. Albiston, A.M. Allen, C.N. May and R.M. McAllen et al., 2003. The brain rennin-angiotensin system: Location and physiological roles. Int. J. Biochem. Cell. Biol., 35: 901-918.
Direct Link - Saad, W.A., I.F.M.S. Guarda, L.A.A. Camargo, T.A.F.B. Santos, W.A. William and S. Simoes, 2002. Role of nitric oxide and beta receptors of the central nervous system on the salivary flow induced by pilocarpine injection into the lateral ventricle. Pharmacol. Biochem. Behav., 72: 229-235.
Direct Link - Saad, W.A., L.I. Gutierrez, I.F.M.S. Guarda, L.A.A. Camargo and T.A.F.B. Santos et al., 2004. Interaction between supraoptic nucleus and septal area in the control of water, sodium intake and arterial blood pressure induced by injection of angiotensin II. Pharmacol. Biochem. Behav., 77: 667-674.
CrossRefDirect Link - Saad, W.A., L.I. Gutierrez, I.F.M.S. Guarda, L.A.A. Camargo and T.A.F.B. Santos et al., 2004. Nitric oxide of the supraoptic nucleus influences the salivary secretion, sodium renal excretion, urinary volume and arterial blood pressure. Life Sci., 74: 1593-1603.
CrossRefDirect Link - Saad, W.A., I.F.M.S. Guarda, L.A.A. Camargo, R.S. Guarda, W.A. Saad and T.A.F.B. Santos, 2006. Influence of vasopressin receptor and nitric oxide on the water, sodium intake and arterial blood pressure induced by angiotensin injected into the third ventricle of the rat brain. J. Biol. Sci., 182: 182-186.
- Saad, W.A., I.F.M.S. Guarda, L.A. de Arruda Camargo, W.A. Saad, R.S. Guarda, T.A.F.B. Santos and S. Simoes, 2006. Endothelial and neuronal nitric oxide synthase inhibitors influences angiotensin II pressor effect in central nervous system. Int. J. Pharmacol., 2: 341-347.
CrossRefDirect Link - Thunhorst, R.T. and A.K. Johnson, 1994. Renin-angiotensin, arterial pressure and salt appetite in rats. Am. J. Physiol., 266: R458-R465.
Direct Link - Tsutsumi, K. and J.M. Saavedra, 1991. Characterization and development of angiotensin II receptors subtypes (AT1 and AT2) in rat brain. Am. J. Physiol., 261: R209-R216.
Direct Link - Vacher, C.M., H. Hardin-Pouzet, H.W. Steinbusch, A. Calas and J. De Vente, 2003. The effect of nitric oxide on magnocellular neurons could involve multiple indirect cyclic GMP-dependent pathways. Eur. J. Neurosci., 17: 455-466.
Direct Link - Wu, S.Y., S.L. Dun, V. Forstemann and N.J. Dun, 1997. Nitric oxide and excitatory post-synaptic currents in immature rat sympathetic preganglionic neurons in vitro. Neuroscience, 79: 237-245.
Direct Link - Yu, K.L., Y. Tamada, F. Suwa, Y.R. Fang and C.S. Tang, 2006. Age-related changes in oxytocin- arginine vasopressin- and nitric oxide synthase-expressing neurons in the supraoptic nucleus of the rat. Life Sci., 780: 1143-1148.
Direct Link - Renaud, L.P. and C. Bourque, 1991. Neurophysiology and neuropharmacology of hypothalamic magnocellular neurons secreting vasopressin and oxytocin. Prog. Neurobiol., 36: 131-169.
Direct Link