
Suhail Rasool
Department of Laboratory Medicine, All India Institute of Medical Sciences, New Delhi India-110029 and Amity Institute of Biotechnology, Noida India
B.L. Jailkhani
Department of Laboratory Medicine, All India Institute of Medical Sciences, New Delhi India-110029 and Amity Institute of Biotechnology, Noida India
M. Irshad
Department of Laboratory Medicine, All India Institute of Medical Sciences, New Delhi India-110029 and Amity Institute of Biotechnology, Noida India
Madhuri Behari
Department of Neurology Neurosciences Center, All India Institute of Medical Sciences, New Delhi India-110029 and Amity Institute of Biotechnology, Noida India
Shahnaz Suhail
Department of Neurology Neurosciences Center, All India Institute of Medical Sciences, New Delhi India-110029 and Amity Institute of Biotechnology, Noida India
H. Shabirul
Department of Laboratory Medicine, All India Institute of Medical Sciences, New Delhi India-110029 and Amity Institute of Biotechnology, Noida India
Journal of Medical Sciences
Year: 2007 | Volume: 7 | Issue: 2 | Page No.: 195-202







ABSTRACT
High affinity β-bungarotoxin binding protein was purified from skeletal muscle of human cadaver by affinity chromatography using β-bungarotoxin as a ligand. The SDS-PAGE of purified protein revealed two major protein bands with molecular weight of 86 and 68 kD. The purified protein has a kD of 2.3±0.15 nmoles and binding sites of 34±2.3 f mole/mg tissue. Immunoreactivity profile of purified presynaptic receptor (β-bungarotoxin binding protein) from muscle with myasthenic sera was negatively affected by treatment with sodium-metaperiodate, glucosidase and trypsin, where as no effect was seen on treatment with lipase. This provides evidence that purified presynaptic receptor (β-bungarotoxin binding protein) is a glycoprotein.
PDF Abstract XML References
How to cite this article
Suhail Rasool, B.L. Jailkhani, M. Irshad, Madhuri Behari, Shahnaz Suhail and H. Shabirul, 2007. Purification of Beta Bungarotoxin (β-Bgtx) Binding Protein from Human Cadaver Skeletal Muscle. Journal of Medical Sciences, 7: 195-202.
DOI: 10.3923/jms.2007.195.202
URL: https://scialert.net/abstract/?doi=jms.2007.195.202
DOI: 10.3923/jms.2007.195.202
URL: https://scialert.net/abstract/?doi=jms.2007.195.202
INTRODUCTION
Myasthenia Gravis (MG) is an autoimmune disease that results from antibody mediated damage of acetylcholine receptor (AChR) at the neuromuscular junction (Patrick and Lindstrom, 1973). Much evidence has been presented which supports this hypothesis, including the induction of Experimental Autoimmune MG (EAMG) by immunization with AChR (Patrick and Lindstrom, 1973). Anti-AChR antibodies acting on the AChR may thereby causes the abnormal muscular fatigue and other signs of MG (Lindstrom et al., 1976; Lefvert and Bergstom, 1978 ). The autoimmune character of MG and pathogenic role of AChR antibodies have been established by several workers e.g; the demonstration of anti-AChR antibodies in about 90% of MG patients (Lindstrom et al., 1976; Brenner et al., 1978; Dwyer and Bradly, 1979; Vincent et al., 2003). It has been demonstrated that patients with MG have also antibodies against second protein named presynaptic membrane receptor (PsmR), which is identified by utilizing β-Bgtx, a ligand which binds to PsmR (Link, 1992). PsmR represents another antigen besides AChR relevant for development of MG. β-Bgtx is a neurotoxin isolated from venom of Bungarus multicinictus and acts exclusively on the presynaptic side of the neuromuscular junction (Chang and Lee, 1962) which is known as presynaptic membrane receptor (PsmR). This presynaptic membrane receptor (PsmR) has been isolated from electroplax tissue of Torpedo californica (Qiao, 1994) and fetal bovine diaphragm muscle (Lu, 1991) by utilizing β-Bgtx. The present study aims to purify the presynaptic membrane receptor from skeletal muscle of human cadaver
MATERIALS AND METHODS
Beta bungarotoxin (β-Bgtx),Tween-20, Bovine Serum Albumin (BSA) benzethonium chloride, benzamidine hydrochloride, phenylmethyl sulphonyl floride, bacitracine, trypsin, sodiummetaperiodate, lipase, CNBr activated Sepharose 4B were all purchased from Sigma Aldrich, USA. All other reagents were used of Analytical Grade (AR).
Solublization of receptor: The receptor from skeletal muscle of human cadaver was solubilized according to the method as described by Jailkhani et al. (1986). Briefly the tissue were minced and homogenized at 4°C in 4 volumes of 0.01 M phosphate buffer pH 7.4 containing 0.1 M NaCl, 0.02% NaN3, 0.001 M EDTA, 0.1 M benzethonium chloride, 0.002 M benzamidine hydrochloride, 0.0001 M phenylmethyl sulphonyl fluoride (PMSF) and 0.5 mg mL-1 bacitracin (homogenizing buffer). The homogenate was then centrifuged at 20,000 g for 60 min at 4°C. The pellet obtained at 20,000 g centrifugation of tissue homogenate was extracted for 3 h at 4°C in 2 volumes of homogenizing buffer containing 2% (v/v) triton X-100. The supernatant (triton extract) obtained on centrifugation at 20,000 g for 60 min was filtered through glass wool, aliquoted and will be stored at -20°C or below as a source of solubilized receptor.
Protein estimation: The protein concentration of different preparation of antigens was determined by the method of Lowery using BSA as a standard (Lowry et al., 1951). However in case of triton extracts 2% (v/v) of triton X-100 was used in the standard protein solution and samples centrifuged (in order to remove the precipitate formed) before reading the absorbance (Gotti et al., 1982).
Coupling of β-Bgtx CNBr (Cynogen bromide ) activated sepharose-4B: CNBr (Cynogen bromide) activated Sepharose-4B was used for affinity chromatography. CNBr activated sepharose 4B was washed and swelled for 30 min in ice-cold HCl, followed by washing with distilled water. The gel was then washed with coupling buffer-0.1 M NaHCO3 containing 0.5 M NaCl, with a pH of 8.3-8.5. The washed gel was then mixed with the protein solution (β-Bgtx dissolved in coupling buffer) and mixed overnight at 4°C with gentle stirring. The supernatant was removed and the unreacted ligand washed away. The unreacted groups on the gel were blocked by using 0.2 M glycine pH 8.0 for 16 h at 4°C. Blocking was followed by 5 alternate cycles of washing with high and low buffer solutions: coupling buffer, pH 8.3-8.5 and 0.1 M acetate buffer, pH 4. The gel was then equilibrated with the buffer (homogenizing buffer with 0.5 M NaCl and 0.1% v/v triton X-100).
Affinity purification of β-Bgtx binding protein on β-Bgtx-sepharose 4B gel: The affinity gel was packed in a column and equilibrated with homogenizing buffer. The triton extract of skeletal muscle was loaded and re-passaged onto the affinity gel till nearly all the β-Bgtx binding protein was bound to gel. Washing was done extensively with homogenizing buffer containing 0.5 M NaCl and triton X-100 whose concentration was reduced from 1-0.1% v/v. The elution was done with 1 M ammonium hydroxide. The eluate was dialyzed against the homogenizing buffer to remove the ammonium hydroxide. The concentrated protein was then assessed for immunological characteristics (ELISA with myasthenic pool sera), toxin binding (radio-iodinated α or β- Bgtx), nature and molecular weight (SDS-PAGE).
SDS-PAGE: For knowing the subunit composition, purified receptor preparation was mixed with an equal volume of 2X sample buffer (containing 0.5 M Tris HCl pH 6.8, 20% SDS, 20% glycerol, 0.1 M EDTA, 25% β-mercaptoethanol and 0.1% bromophenol blue ) and heated at 100°C for 10 min. The supernatant was loaded on the gel. SDS-PAGE was conducted in 10% polyacrylamide resolving gel and 5% stacking gel containing 1% SDS (Laemmli, 1970).
Electrophoresis was performed at a constant current of 30 MA until bromophenol blue reaches the bottom of the gel.
Immunoblotting: The protein from SDS-PAGE gel was transferred onto nitrocellulose sheet after equilibration in tris glycine buffer. Transfer was carried out in a wet type transfer system at 15 volts for 16-18 h at 4°C. The nitrocellulose membrane after transfer of proteins was blocked by 5% milk powder in tris saline (containing 50 mM Tris pH 7.5, 150 mM NaCl with 0.05% Tween-20 TTBS ). The nitrocellulose paper was immersed in pooled myasthenic sera for 1 h at room temperature. The membrane was washed thrice with TTBS and incubated for 1 h at room temperature in anti-human IgG-HRP conjugate diluted in tris buffer saline with 5% milk powder. These membrane was washed with TBS and the incubated with substrate solution (5 mg Diaminobenzidine in Tris buffer saline with 5 μL H2O2) and reaction was stopped by washing the blot once with TBS.
Treatment with sodium metaperiodate and enzymes: The purified protein from muscle was coated directly at pH 9.7 on ELISA plates and treated with sodium metaperiodate and the enzymes: trypsin, lipase and glucosidase, for 3 h at 37°C. After washing the ELISA was carried out and the effect on immunoreactivity with the myasthenic pool sera tested.
RESULTS
β-Bgtx was coupled to CNBr-activated Sepharose 4B at a concentration of 3 mg g-1 of the dry gel. The actual amount of β-bungarotoxin coupled to the gel was determined by competition ELISA. The precoupling toxin solution and post coupling supernatant were assessed for their ability to compete with known concentration of β-bungarotoxin for binding presynaptic membrane receptor in the triton extract. In the two preparations, 43.7 and 42% of the toxin was found to be coupled to the gel.
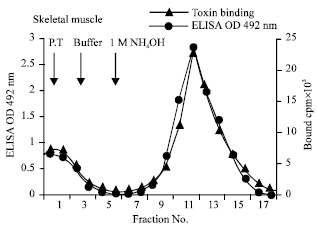 | |
| Fig. 1: | Affinity purification on β-Bgtx sepharose 4B column. 40 mL of triton extract from skeletal muscle and parietal lobe of cadaver brain were passed (3 times) through β-Bgtx Sepharose 4B column. After extensive washing (100 mL), the receptor was eluted with 1M NH4OH. 1M fractions were collected, dialyzed to remove NH4OHand then assessed by indirect ELISA and radioiodinated β-Bgtx binding |
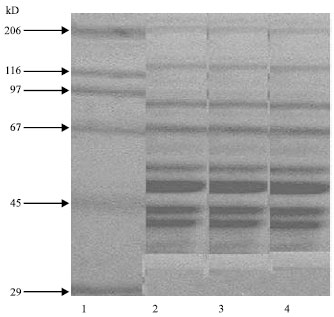 | |
| Fig. 2: | SDS-PAGE of triton extract from different human skeletal muscle. Triton extract from different skeletal muscle, were run on SDS-PAGE using a stacking gel of 5% and a resolving gel of 10%. Electrophoresis was carried out at 100 volts for 3 h. The gel was stained with Commassie brilliant blue and destained |
Freshly prepared triton extract from muscle. The extracts were repassed 3-4 times to achieve the maximum adsorption of β-bungarotoxin binding proteins. The elution of the presynaptic receptor was done with 1 M NH OH by stepwise elution, collecting 1 ml 4 fraction.
| Table 1: | Affinity purification of presynaptic receptor from muscle |
 | |
| Specific activity: 0.91±0.06, Recovery: 43±2.6, Fold purification: 2633±120 | |
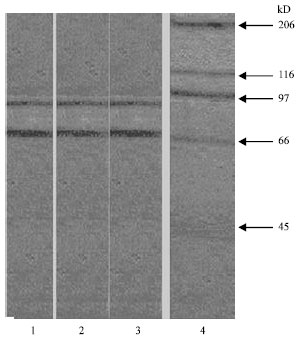 | |
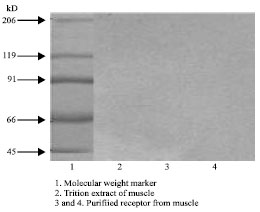
| Fig. 3: | 1, 2, 3: Affinity purified receptor 4: Molecular weight marker. SDS-PAGE of purified presynaptic receptor ($-Bgtx binding protein) from human skeletal muscle. Affinity purified $-bungarotoxin binding protein from skeletal muscle (4 µg protein) were run on SDS-PAGE of purified presynaptic receptor (Beta Bgtx binding protein) from human skeletal muscle. Affinity purified b-bungarotoxin binding protein from skeletal muscle (4 μg protein) were run on SDS-P AGE using a stacking gel of 5% and a resolving gel of 10%. Electrophoresis was carried out at 100 volts for 3 h. The gel was containing the purified protein was silver stained |
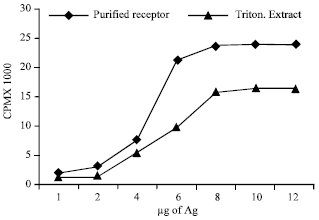 | |
| Fig. 4: | Radio-iodinated β-Bgtx binding profile of Purified receptor of muscle. Indicated volumes of triton extract and affinity purified β-Bgtx binding receptor from skeletal muscle were incubated with l.5x105cpm of radio-iodinated β-Bgtx using the ammonium sulfate precipitation method |
| Table 2: | Affinity and number of Beta-Bgtx binding sites in whole triton extract and purified presynaptic receptor of cadaver muscle. |
 | |
The eluted fractions were dialyzed to remove NH4OH. The immunoreactivity was determined by ELISA using MG sera and by ligand binding assay using 125I-β-Bgtx as shown in Fig. 1. The details of affinity purification of presynaptic receptor from skeletal muscle shown in Table 1. Four preparation of skeletal muscle were affinity purified. The mean sp activity, recovery and fold purification are shown in Table 1.
Figure 2 shows the SDS PAGE profile of triton extract, which consists of numerous proteins. Figure 3 shows the SDS PAGE profile of purified presynaptic membrane receptor (β-Bgtx binding protein) of skeletal muscle. In the purified preparations, there were two prominent bands corresponding to 87 and 68 kd.
Ligand binding characteristics: 125I-_β Bgtx binding activity was compared in the purified presynaptic receptor of muscle with its corresponding triton extracts. It was observed that purified presynaptic receptor has more toxin binding activity as compared to its corresponding triton extract as shown in Fig. 4.
The specificity of 125I-_β Bgtx binding in the purified presynaptic receptor (β-Bgtx binding protein) preparations and the corresponding triton extract of skeletal muscle was demonstrated by competition with cold β-Bgtx, scatchard plots were constructed (Fig. 5). The binding characteristics Bmax and kd calculated from the scatchard plots are shown in Table 2.
Biochemical nature: Purified presynaptic receptor (β-Bgtx binding protein) was subject to enzymatic treatment with trypsin, glucosidase, lipase, sodium meta periodate and the effect on the immunoreactivity with pooled MG sera was assessed by ELISA (Fig. 6). Treatment with sodium metaperiodate, glucosidase and trypsin were shown a significant decrease in the immunoreactivity of purified presynaptic receptor of muscle, where as lipase did not produce any effect.
 | |
| Fig. 5: | Effect of cold b-bungarotoxin on binding of radio-iodinated b-bungarotoxin to triton extract and purified presynaptic receptor of skeletal muscle. Fifty microliter of triton extract and purified presynaptic receptor from skeletal muscle were incubated with 1.5x105 cpm of radio-iodinated b-bungarotoxin and competed with different concentration of non radio-active b-bungarotoxin. The scatchard plots were made by plotting [B]/[F] against [B]. [B]: Concentration of toxin bound at each input concentration of cold toxin. [F] : Input concentration of toxin |
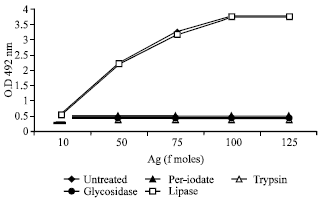 | |
| Fig. 6: | Effect of various enzymes on immunoreactivity of purified beta bungarotoxin binding receptor. Purified receptor from skeletal muscle was coated onto ELISA plate wells by direct coating at pH 9.6 for 2 h at 37°C. 200 μL of sodium mateperiodate (50 mg mL-1), trypsin (1 mg mL-1), lipase (0.5 mg mL-1) and glucosidase (0.25 mg mL-1) were then added to the wells and incubated for 3 h at 37°C. Untreated. After washing the receptor was sequentially reacted with MG sera pool and HRPO conjugated secondary antibody as usual and the effect on immunoreactivity assessed |
These result suggested that purified presynaptic receptor is a glycoprotein as immunoreactivity resides in the carbohydrate as well as the peptide epitopes.
Immunological characteristics: Immunoreactivity of the purified receptor was greater than that of the receptor in the triton extract when coated through β-Bgtx (Fig. 7), where as no biding was seen in the purified preparation when coated through α-Bgtx (Fig. 8).
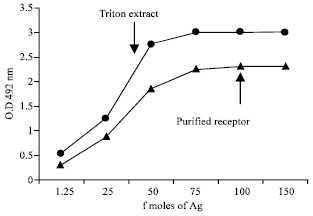 | |
| Fig. 7: | Titration of purified presynaptic receptor of parietal lobe and muscle, along with their respective triton extract on b-Bgtx precoated plates. Triton extract and affinity purified protein from skeletal muscle containing 100 fmoles of b-Bgtx binding sites was coated on to ELISA plate wells indirectly through 1 ug mL-1 of toxin b-Bgtx at pH 9.6. Immunoreactivity was then seen with MG sera pool |
Thereby indicated the specificity of the pure receptor preparation with regards to autoantibodies in pool MG sera.
The affinity purified presynaptic receptor (β-Bgtx binding protein) of muscle, triton extract of muscle and brain (parietal lobe) showed immunological cross reactivity with each other where as liver triton extract did not show any competition (Fig. 9).
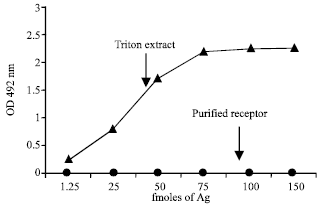 | |
| Fig. 8: | Titration of purified presynaptic receptor of muscle along with its respective triton extract on β-Bgtx precoated plates. Triton extract and affinity purified protein of muscle and brain was coated on to ELISA plate wells indirectly through 1 μg mL-1 of toxin a-Bgtx. Immunoreactivity was then seen with MG sera pool |
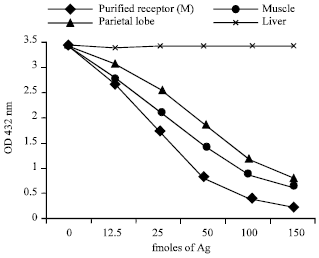 | |
| Fig. 9: | Competition ELISA of purified receptor of muscle with other antigens. 100 fmoles of affinity purified b-bungarotoxin receptor of muscle was coated onto ELISA plate through 1 ug mL-1 b-bungarotoxin. Indicated concentrations of the competing antigens were pre-incubated with the MG sera pool (1:200 dilution at overnight at 4°C) and then reacted with the coated antigen followed by the conjugate and substrate |
Immunoblot: Five to seven microgram of the purified protein along with triton extract of muscle was electrophoresed on SDS-PAGE. Later gel was transferred on nitrocellulose paper. The blocked membrane was then reacted with pooled MG sera. In the purified protein of muscle, two bands were observed of 87 and 68 kd (Fig. 10). No reactivity was seen with control sera pool (Fig. 11). This indicated that both the two subunits of purified receptors from muscle are immunoreactive.
 | |
| Fig. 10: | Immunoblot of triton extract and purified presynaptic receptor from muscle of human cadaver with myasthenic sera. Affinity purified presynaptic receptor (5-7 μg protein) from skeletal muscle, were run at 100 volts, for 3 h on SDS-P AGE. The protein was then transferred to nitrocellulose membrane at 20 volts for 12 h in a wet transfer apparatus. The membrane was probed with MG sera pool at a dilution of 1: 500 to indicate the MG specific immunoreactive bands. Molecular weight markers are indicated on the left-hand side |
 | |
| Fig. 11: | Immunoblot of purified presynaptic receptor of muscle and brain of human cadaver with control sera. Affinity purified presynaptic receptor (5-7 μg protein) from skeletal muscle, were run at 100 volts, for 3 h on SDS-P AGE. The protein was then transferred to nitrocellulose membrane at 20 volts for 12 h in a wet transfer apparatus. The membrane was probed with control sera pool at a dilution of 1: 500. Molecular weight markers are indicated on the left-hand side |
DISCUSSION
MG is considered as an autoimmune disease caused by antiacetylcholine (anti-AChR) antibodies, resulting in functional loss of AChR at the neuromuscular junction (Vincent and Newsom-Davis, 1979; Lindstrom et al., 1976). The basic abnormality in myasthenia gravis is a decrease in the number of acetylcholine receptors at the neuromuscular junctions which causes fatigabilty of the muscle (Drachman et al., 1978). This was first identified by use of radiolabelled snake toxin α-Bgtx, which binds specifically, quantitatively and irreversibly to acetylcholine receptor of skeletal muscles (Chang and Lee, 1962). α-Bgtx, has been long used for specific identification, quantification and purification of the receptor (Pestronk, 1985; Conti-Tronconi and Raftery, 1982). Patients with MG have also antibodies against a second protein which is called presynaptic membrane receptor (PsmR), which has been been isolated from bovine diaphragm muscle utilizing β-Bgtx (Xiao, 1991; Link, 1992 ). Antibodies of PsmR and AChR from MG patients sera showed about 45-55% cross reactivity and there-is high correlation between serum levels of both antibodies (Xiao, 1991). The present study was to purify presynaptic receptor (β-Bgtx binding protein) from muscle of human cadaver. The use of β-Bgtx CNBr activated sepharose 4B affinity matrix was used for purification of presynaptic receptor. The elution of the purified presynaptic receptor (β-Bgtx binding protein) was done with 1 M NH4OH. The immunoreactivity and toxin binding profile of the fractions eluted from the affinity column for skeletal muscle (Fig. 1). Earlier β-Bgtx binding protein was purified from bovine diaphragm, torpedo by affinity chromatography using CNBr activated sepharose 4B. The bound protein was eluted with 0.5 M KCl, sequentially loaded on wheat germ lectin column and eluted by N-acetylglucoseamine (Xiao, 1991; Qiao, 1994). The SDS- PAGE profile of purified presynaptic receptor (β-Bgtx binding protein) of skeletal muscle as shown in figure showed two prominent bands corresponding to 87 and 68 kd. Immunoreactivity profile of purified presynaptic receptor (β-Bgtx Binding protein) from muscle with myasthenic sera was negatively affected by treatment with sodium-metaperiodate, glucosidase and trypsin, where as no effect was seen on treatment with lipase. This provides evidence that purified presynaptic receptor (β-Bgtx binding protein) is a glycoprotein as reported earlier (Xiao, 1991). The specificity of 125I-_β Bgtx binding in the purified presynaptic receptor preparations and the corresponding triton extracts of skeletal muscle was demonstrated by competition with cold β-Bgtx and scatchard plots were constructed. The binding characteristics Bmax and kd calculated from the scatchard plots (Table 2 ). Immunoblotting of the purified protein of muscle with pooled myasthenic sera gave a prominent bands of 87 and 68 kd (Fig. 10) and no reactivity was seen with control sera pool (Fig. 11).
REFERENCES
- Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Patrick, J. and J. Lindstrom, 1973. Autoimmune response to acetylcholine receptor. Science, 180: 871-872.
CrossRefDirect Link