
Raesa A. Mohamed
Department of Histology, Faculty of Medicine, Cairo University, Egypt
Hoda M. El Aasar
Department of Anatomy, Faculty of Medicine, Cairo University, Egypt
Lobna A. Mohamed
Department of Histology, Faculty of Medicine, Cairo University, Egypt
Amal M. Abbas
Department of Histology, Faculty of Medicine, Cairo University, Egypt
Journal of Medical Sciences
Year: 2007 | Volume: 7 | Issue: 2 | Page No.: 161-169







ABSTRACT
This study was performed on human muscle autopsies, from triceps of the left arm, obtained from 23 non-athletic built males, with no evidence of neuromuscular involvement. Their age covered the range from one month to 81 years. They were divided into four groups according to their age namely, the infant, the child, the adult and the elderly groups. The autopsy specimens were processed for light and transmission electron microscopical study. As for light microscopy, they were subjected to hematoxylin and eosin and Masson trichrome staining methods. An image analysis study was included to estimate the mean cross sectional area of the muscle fiber, the mean area percent of the fibrocollagenous septa enclosed between the muscle bundles and the mean satellite cell counts. The study revealed the presence of considerable variations in the histological structure of the skeletal muscle with the advancement of age. The cross sectional area of the skeletal muscle fiber reached its peak value in the adult group followed by marked regression in the elderly group. An expected continuous spectrum of growth and maturation was observed in the fibrocollagenous septa between the muscle bundles. These septa became densely packed and irregular in the elderly age group. The maximum number of satellite cells was observed in the child group followed by a decline in the adult group to become scarce and pyknotic in the elderly one. Ultrastructurally, in senescent individuals, the sarcolemma appeared irregular, with irregular arrangement of the dark and light bands and the mitochondrial aggregates appeared ballooned and degenerated. These observations could suggest that muscle atrophy is a major contributor to the decrease in muscle strength manifesting with advancement of age.
PDF Abstract XML References Citation
How to cite this article
Raesa A. Mohamed, Hoda M. El Aasar, Lobna A. Mohamed and Amal M. Abbas, 2007. Morphological Features of Normal Human Skeletal Muscle in Different Age Groups: A Histological and Ultrastructural Study. Journal of Medical Sciences, 7: 161-169.
DOI: 10.3923/jms.2007.161.169
URL: https://scialert.net/abstract/?doi=jms.2007.161.169
DOI: 10.3923/jms.2007.161.169
URL: https://scialert.net/abstract/?doi=jms.2007.161.169
INTRODUCTION
In the skeletal muscle fibers, aging results in a general slowing of the mechanical characteristics of the muscle (Kirkeby and Garbarsch, 2002) and predictable age-induced alterations in the striated muscles were documented (Akataki et al., 2002). To assess the age-related loss of muscle mass and to determine the mechanisms behind this aging atrophy, the muscle structure and fiber type composition have been estimated using invasive and non-invasive techniques (Lexell, 1995; Fayet et al., 2001; Klein et al., 2003). Significant reduction in skeletal muscle isokinetic strength, loss of muscle mass and disturbed histochemical fiber characteristics were observed in senescent individuals (Frontera et al., 2000; Klein et al., 2003; Karakelides and Sreekumaran, 2005). The measurement of the aerobic and anaerobic enzymatic activities of skeletal muscles was impaired in elderly individuals compared to younger age groups (Pansarasa et al., 2002). At the cellular level, muscle looses both cross sectional area and fiber number (Manta et al., 1996), with type II muscle fibers being the most affected by aging (Manta et al., 1995; Kirkendall and Garrett, 1998; Fayet et al., 2001). On the other hand, Klein et al. (2003) ascribed the reduction of the maximal area of the skeletal muscle, by aging, to the reduction of the muscle fiber size without significant difference in the estimated number of muscle fibers between the young and old men.
The number, the mitotic activity and the differentiation potential of satellite cells, which are responsible for postnatal muscle fiber formation and the muscle fiber diameter are tightly regulated events in normal muscle (Maier and Bornmann, 1999). The effect of aging on the satellilte cell population has been studied by Renault et al. (2003) and Kadi et al. (2004) who revealed that a reduction in the satellite cell population with decreasing the muscle regenerative capacity occurred as a result of increasing age in healthy men and women. However, the capillary supply per unit fiber area was not affected by age but was enhanced by training (Desaki et al., 1996). The understanding of the basis of dysfunction of skeletal muscles in old age has important clinical and social implications. The available literature seems to be deficient in coverage of the histological aspects associated with this age-related dysfunction. Hence, the present study aimed to examine the histological changes induced by the advancement of age in human male triceps muscle trying to overcome this knowledge gap.
MATERIALS AND METHODS
This study was performed, during the period between January 2004 and February 2005, on human skeletal muscle autopsies taken from refrigerated fresh corpses, within 2-6 h of death, kept (4°C) in the postmortem room of the Faculty of Medicine, Cairo University, Egypt. The autopsies were taken from 23 average non-athletic built males, after excluding any case with history of neuromuscular involvement. Their age ranged from one month to 81 years old with a mean of 47.6±23.2. They were classified into four groups:
Infant group: Which included 4 infants, with an age ranging from one month to 11 months (mean age 0.7 years ±0.6).
Child group: Which included 3 children, with an age ranging from 2 years to 15 years (mean age 7.4 years ±3.6).
Adult group: Which included 7 cases, with an age ranging from 24 to 50 years (mean age 37.7 years ±5.8).
Elderly group: Which included 9 cases, with an age ranging from 65 to 81 years (mean age 74.6 years ±4.6).
In each case, the skeletal muscle specimen was taken from the middle of the medial part of long head of triceps muscle of the left arm. For the light microscopical study, specimens were fixed in 10% neutral buffered formalin and embedded in paraffin wax. From each block, five micrometer-thick cross sections were cut and subjected to Hx and E and Masson trichrome staining (Bancroft and Gamble, 2002). For the transmission electron microscopical study, the specimens were fixed in buffered glutaraldehyde solution for 6 h and then transferred to phosphate buffered saline overnight. All samples were then post-fixed in buffered osmium tetroxide and dehydrated in alcohol then embedded in araldite. Semithin sections were obtained and stained with toluidine blue for light microscopical examination. The ultrathin sections were prepared and double stained with uranyl acetate and lead citrate to be examined using a Joel-100S transmission electron microscope (Bancroft and Gamble, 2002).
Morphometric study: The mean Cross Sectional Area (CSA) of the individual muscle fiber in the H and E stained sections and the mean area percent of fibrocollagenous septa comprising the perimysium in the Masson trichrome stained sections were analyzed morphometrically for each case. The mean satellite cell count per field was also morphometrically calculated from the semithin toluidine blue-stained sections. The data were analyzed using Leica Quin 500 image analyzer. Sections or areas with artifacts or indistinct cell borders were excluded from the counts. In each specimen, ten fields were examined using an eye piece of X 10 and an objective of X 40 i.e., at a magnification of 400 and the mean values were calculated.
Statistical analysis: Results were expressed in the form of mean±standard deviation. The one way analysis test of variants (Duncan test) was used in this study. Differences were considered significant at p≤0.05.
RESULTS
Histological results: Microscopical examination of Hx and E stained sections (Table 1 and Fig. 1) revealed that in the infant group, the muscle fibers appeared small in size. Most of the fibers had a rounded to oval contour. In the child group, an increase in the size of the muscle fibers was observed. The muscle fibers exhibited a polyhedral pattern with extreme peripheral location of the nuclei. In the adult group, there was a remarkable increase in the size of the muscle fibers. The muscle fibers showed a roughly polyhedral contour with flattened sides where they were compressed by adjacent fibers. Nuclei appeared as small oval profiles at the periphery of each fiber. In the elderly group, some variability in the muscle fiber size was noticed, but as a whole, a marked reduction in the muscle fiber size was noticed compared to the adult group. A statistical level of significance was reached comparing the CSA of each group to the others (p<0.05). It is noteworthy to mention that the satellite cells were not clearly discernible in Hx and E sections.
Microscopical examination of the masson trichrome stained sections: Table 1 and Fig. 2 showed that minimal amount of the fibrocollagenous septa comprising the perimysium was observed in the infant group. The collagen bundles were very thin and irregularly extending between the bundles. A slight increase in the amount of the septa was noticed in the child group and the septa were more regularly arranged between the muscle bundles. In the adult group, the mean area percent of the septa reached and they appeared broad and regularly arranged. In the elderly group, a marked increase in the amount of the fibrocollagenous septa was reported. The septa became densely packed and irregularly arranged with widening of the spaces between the muscle bundles.
| Table 1: | Measurements of CSA of muscle fibers, area percent of fibrocollagenous septa and mean satellite cell count/HPF in muscle specimens of all age groups |
 | |
| CSA = Cross Sectional Area FCS = Fibrocollagenous Septa HPF = High Power Field | |
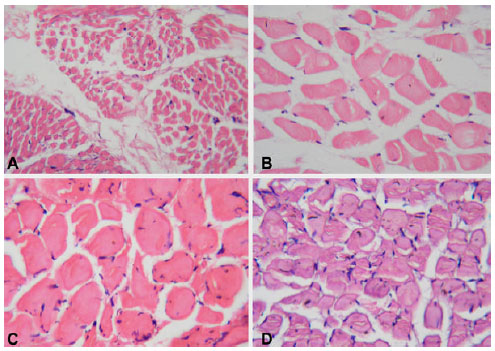 | |
| Fig. 1: | Photomicrographs of cross sections in human skeletal muscle fibers stained by Hx and E. (X 200) showing: (A): In an infant, 7 months-old, small-sized muscle fibers. Note the oval to round contour of the fibers, (B): In a child, 12 years old, an increase in the size of the muscle fibers. The fibers exhibit a polyhedral pattern with extreme peripheral location of the nuclei, (C): In an adult, 35 years-old, a remarkable increase in the size of the muscle fibers, which appear polyhedral with flattened side. Their nuclei appear as oval profiles at the periphery of each fiber, (D): In a 71 years-old male, marked variability in the muscle fiber size with overall reduction, compared to the adult group. Note the irregular outer borders of the muscle fibers |
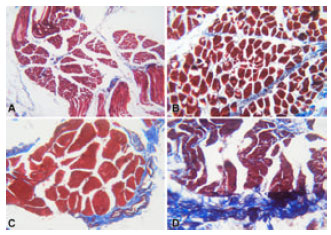 | |
| Fig. 2: | Photomicrographs of cross sections in the human skeletal muscle fibers stained with Masson trichrome (X 100) showing: (A): In a seven months-old infant, thin collagenous fibers running irregularly between the muscle bundles, (B): In a 12 years-old child, slightly more packed collagen bundles than in infants, (C): In a 35 yearsold adult, broad and uniformly arranged fibrocollagenous septa, (D): In a 76 years-old male, densely packed and irregulary arranged fibrocollagenous fibers with widening of the spaces between the muscle bundles |
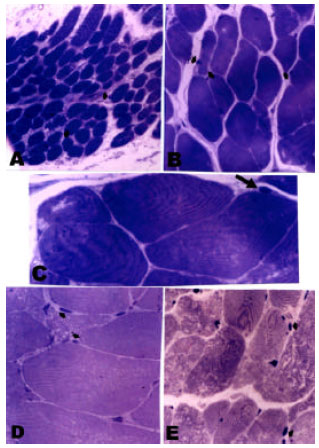 | |
| Fig. 3: | Photomicrographs of toluidine blue-stained semithin sections in human skeletal muscles showing: (A): In an 11 months-old infant, ill-defined myofibrils with some scattered satellite cells (arrows) (X400), (B): In a child 12 years-old, more clear myofibrils and more satellite cells (arrows), compared with the infant group (X 400), (C): A higher magnification in a child muscle revealing the satellite cell (arrow) as small spindle shaped cell located immediately beneath the external lamina of the muscle fiber (X 1000), (D): In an adult 41 years-old, clearly identified cut ends of the myofibrils, which appear as numerous dark fine dots. Fewer satellite cells (arrows), compared to the child group, are noticed (X 400), (E): In an 81 years-old male, irregularly distributed myofibrils within the muscle fibers with scarce pyknotic satellite cells (arrows) (X 400) |
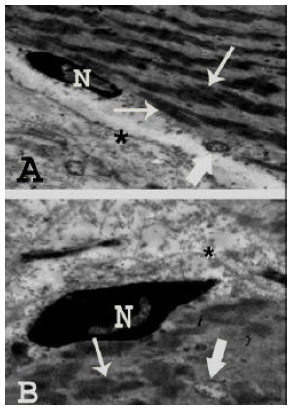 | |
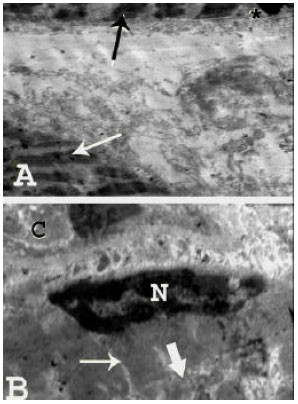
| Fig. 4: | Transmission electron micrographs of human skeletal muscle fibers in a 7-month old infant revealing: (A): A slightly irregular sarcolemma (asterisk). Dark and light bands are not well-defined (arrows) with normally appearing nucleus (N) and mitochondria (thick arrow) (TEM mag. X 2000; print mag. X 10587), (B): A nucleus (N) and uneven arrangement of the myofibrils with no clear striations (thin arrow). The cytoplasm in between the myofibrils contains few scattered apparently normal mitochondria (thick arrow) with well-developed sarcoplasmic reticulum. Note the slightly redundant sarcolemma (asterisk). (TEM mag. X 3000; print, mag. X 15880) |
Statistical level of significance was reported when comparing the mean area percent of the septa of each group to the other groups (p<0.05), except when comparing infant to the child group (p>0.05).
Semithin toluidine blue-stained sections examination: Figure 3 revealed that the transverse sections of skeletal muscles appeared to be packed with numerous dark fine dots representing the cut ends of the myofibrils. These myofibrils usually occupied most of the interior of the fiber. In some sections, obliquely cut muscle fibers revealed the regular transverse striations. These myofibrils were not clearly evident in sections of muscles obtained from infants. However, they became more obvious in the child, adult and elderly groups.
 | |
| Fig. 5: | Transmission electron micrographs of human skeletal muscle fibers in a 12 years-old child revealing: (A): More obvious striations of alternating dark and light bands (arrows). The sarcolemma (asterisk) appears clear and regular (TEM mag. X 2000; print. mag. X 10587), (B): A prominent nucleus (N) and thick myofibrils (thin arrow). Mitochondria (thick arrow) are evident in the cytoplasm enclosed between the myofibrils with more prominent sarcoplasmic reticulum. The intermuscular connective tissue reveals a section in a blood capillary (C) (TEM mag. X 3000; print mag. X 15880) |
Whereas these dots in both the child and adult groups appeared to be regular, fine and uniformly distributed in all the muscle fibers in the examined sections, in the elderly group, irregular dots were observed in most of the fibers. As for the satellite cells, they appeared as small spindle shaped cells which were located immediately beneath the external lamina of the muscle fibers. They were identified in all age groups included in this study (Table 1). In the infant group the mean satellite cell count was 4.5±0.65. They reached their maximum intensity in the child group (mean count 13.7±1.32) followed by progressive decline in the adult group (mean count 8.7±2.43). However, in the elderly age group, satellite cells were barely seen and appeared pyknotic (mean count 1.68±0.78). Statistical level of significance was reached when comparing the mean satellite cell count in each group to the other groups (p<0.05).
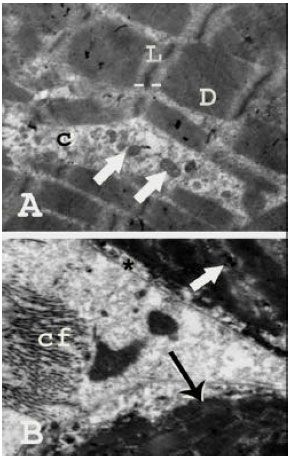 | |
| Fig. 6: | Transmission electron micrograph of human skeletal muscle fibers in an adult aging 35 years revealing: (A): Stacks of large myofibrils composed of alternating and overlapping zones of dark-stained (D) and light stained (L) filaments. A little amount of cytoplasm (C) in between the myofibrils containing a chain arrangement of mitochondria (arrows) surrounded by well-developed sarcoplasmic reticulum is seen (TEM mag. X 4000; print mag. X 21173), (B): Portions of two muscle fibers with clear and regular sarcolemma (asterisk). The thick myofibrils have prominent striations (thin arrow) and are surrounded by little amount of cytoplasm containing mitochondria (thick arrow). The intermuscular connective tissue shows accumulation of collagen fibers (Cf) (TEM mag. X 3000; print mag. X 15880) |
Transmission electron microscopic results: In the infant group (Fig. 4), the sarcolemma was slightly irregular and the myofibrils appeared small in size with no clear bands in most of the fibrils. Normally appearing mitochondria were seen with well-developed sarcoplasmic reticulum. In the child group (Fig. 5), regular sarcolemma, more numerous mitochondria and more prominent sarcoplasmic reticulum were noted. The myofibrils appeared larger than in infant group with more or less clear dark and light bands.
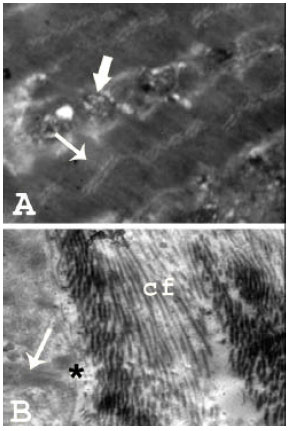 | |
| Fig. 7: | Transmission electron micrographs of human skeletal muscle fibers in an elderly aging 81 years (TEM mag. X 4000; print mag. X 21173) showing: (A): Distorted small myofibrils with lacking of the precise alignment of the dark and light bands (thin arrow). Plenty of cytoplasm with chains of degenerated aggregates of swollen mitochondria (thick arrow) is noticed, (B): Some degenerated myofibrils (arrow) surrounded by slightly irregular sarcolemma (asterisk). The intermuscular connective tissue reveals abundant collagen fiber deposition (Cf) |
As for the adult group (Fig. 6), stacks of large myofibrils composed of clear alternating zones of dark and light filaments were obvious. Numerous normal mitochondria were seen packed between the myofibrils. A well-characterized sarcoplasmic reticulum was also evident. Some collagen deposits could be seen between the muscle fibers. In the elderly age group (Fig. 7), the
sarcolemma was slightly irregular. The myofibrils were obviously small in size, some were degenerated and lacking the precise alignment of the dark and light bands. Some mitochondria appeared swollen and others appeared degenerated. More extensive deposition of collagen fibers was manifested.
DISCUSSION
Aging is a process that is inevitable consequence of senescence. This natural event is due to intrinsic and extrinsic factors. The intrinsic aging, in which genetics accounts for the age of onset and susceptibility, occurs in its pure form in non-exposed body parts. Extrinsic aging is directly related to environmental factors and occurs in habitually exposed body areas. Extrinsic aging is always superimposed on a background of intrinsic aging (Griffiths, 1992). In the skeletal muscle fibers, aging results in a general slowing of the mechanical characteristics of the muscle (Kirkeby and Garbarsch, 2002). Reviewing of the literature revealed that many reports had studied the gerontologic changes in human skeletal muscle, yet data were deficient in targeting the younger age groups including children and infants. Moreover, these studies were dominantly histochemical in design namely targeting the type I and II fibers alterations without focusing on the corresponding histological characteristics. The present study emphasized the histological and ultrastructural changes associated with the advancement of age from infancy to the elderly. This was done in a trial to couple these changes with the physiological muscle specification influenced by aging. In this study, the computerized histomorphometric analysis revealed a progressive increase in the cross sectional area of the human skeletal muscle fiber with age to reach its peak in the adult group. Thereafter, a remarkable, statistically significant, regressive decline was observed in the elderly group. These results were parallel with those of Fayet et al. (2001) in adult and old individuals, but their study did not employ younger age groups. These results could suggest that the senescent atrophy observed in skeletal muscle could result partially from the reduction in the cross sectional area of the single fiber. This suggestion is compatible with the findings presented by Klein et al. (2003) who claimed that the estimated number of muscle fibers was not significantly different between the young and old men and attributed the smaller maximal area of the skeletal muscle (biceps brachii) of old men to smaller muscle fiber size. Moreover, Manta et al. (1995) concluded that the progressive age-related reduction of type2 fiber diameter was a constant finding in all muscle studied-quadriceps femoris, biceps brachii and deltoid-regardless of sex, while Mattiello-Sverzut et al. (2003) revealed this age-related variation, in the size of different types of muscle fibers, only in the studied autopsies and biopsies of male subjects. On the other hand, Lexell (1995) and Kirkendall and Garrett (1998) suggested that aging atrophy seems to be due to a reduction in both number and size of muscle fibers, mainly type 2 and is to some extent caused by a slowly progressive neurogenic process. However, Klein et al. (2003) considered the relative contribution of a reduction in fiber number to age-related muscle atrophy, reported in other studies, to be a muscle-dependent. In addition, Nikolic et al. (2001) concluded that age-related muscle atrophy is not a general phenomenon and does not affect all muscles equally.
In the present study, the fibrocollagenous septa showed a statistically significant rise in its area percent in the elderly individuals compared to other age groups. This result indicated that the gradual decrease in size of the skeletal muscle fibers with advancing age was accompanied by a replacement of connective tissue. This finding was compatible with that reported by Lexell (1995) in his study trying to assess the age-related loss of muscle mass.
In the current study, the maximum number of satellite cells was observed in the child group. Being a resident of the muscle precursor cell, the prevalence of satellite cells in the child group could indicate a high growth and renewal rates of the skeletal muscle fibers in this young age group. This study, also, implies that a marked reduction in the satellite cell population occurred in the elderly group. Satellite cells appeared to be very scarce and pyknotic in this age group. The same results were recapitulated by Maier and Bornmann (1999) and Renault et al. (2003). The latter concluded that the decrease in the regenerative capacity with age might be partially explained by the reduced availability of satellite cells and their degenerative atrophic pattern that was observed in elderly individuals. Therefore, normal aging of skeletal muscle in vivo is reflected by the number of satellite cells available for regeneration, but not by the mean number of myonuclei per fiber or by telomere lengths (Renault et al., 2003). In addition, Maier and Bornmann (1999) proved a significantly higher relative frequency of satellite cells with increasing fiber diameter. Kadi et al. (2004), in their study of satellite cells employing a monoclonal antibody, enforced the previous results of the depletion of such cells with aging in both males and females. Lorenzon et al. (2004) added that ageing affects the excitation-contraction coupling mechanism in human myotubes derived from human satellite cells, thereby contributing to the loss in muscle strength in the aged.
In the present study, Ultrastructurally, the myofibrils attained their typical structure in the adult age group with the clear alternating dark and light filaments. However, in the elderly individuals, lack of precise alignment of the bands was noticed. This finding is compatible with the conclusion deduced from the study of Welle et al. (1996) who revealed that the rate of synthesis of myofibrillar proteins was slower in muscle of healthy subjects over 60 year old than it was in young adults and a single injection of growth hormone could increase muscle mass and strength in those elderly people without restoring a youthful rate of myofibrillar protein synthesis. The most outstanding finding in the electron microscopic results documented in this study was the impaired quality of the mitochondrial content observed in the senescent individuals. Some mitochondria appeared ballooned and others appeared degenerated. Besides, mitochondria tended to form aggregates in this age group. This result was compatible with that reported by Fayet et al. (2001) who added that the increased mitochondrial aggregates with age was observed earlier in females than in males. Poggi et al. (1997) attributed this age-related mitochondrial degenerative changes to the fact that the energy requirements decline with age and that the decrease in the mitochondrial quality in senescent individuals might correlate with reduced metabolic demand. Moreover, Karakelides and Sreekumaran (2005) stated that there is an impairment of mitochondrial protein synthesis as a result of generalized increase of the oxidative activity with ageing and this contributes to a decrease in muscle mass and strength, starting as early as the fourth decade of life. This is evidenced by the effectiveness of unilateral resistance exercise training in enhancing the skeletal muscle cellular antioxidant capacity in older adults with an increase in the muscle mass (Parise et al., 2005).
In conclusion, aging appeared to produce remarkable alterations in the histological structure of the skeletal muscles. These changes could correlate with the loss of force and the subsequent inevitable loss of mechanical characteristics reported by Kirkendall and Garrett (1998) in aged skeletal muscles. The existence of these age changes should be taken into account in the interpretation of muscle biopsies of aged individuals. Although neither the aging process nor the subsequent loss of the mechanical characteristics is inevitable, yet Kirkendall and Garrett (1998) and Melichna et al. (1990) in their studies confirmed that these losses could be minimized or even reversed with training. Endurance training could improve the aerobic capacity of the skeletal muscle and resistance training can improve its central nervous system recruitment and increase muscle mass (Kirkendall and Garrett, 1998). In addition, capillary supply per unit fiber area is not affected by age but is enhanced by training (Desaki et al., 1996; Proctor et al., 1995). Therefore, physical activity throughout life should be strongly encouraged to prevent sarcopenia, early onset of disability and much of the age-related impact on skeletal muscles. Skeletal muscle still deserves more extensive exploratory research in large population samples, employing more than one muscle, to elucidate the specific mechanisms responsible for muscle weakness in elderly. As the normal aging of skeletal muscle in vivo is reflected by the number of satellite cells for regeneration (Renault et al., 2003), therefore a definitive mapping for the satellite cells, as well as definition of the possible factors playing roles in their regulation is also recommended. Extensive tissue culture studies might provide the tool for achieving these goals. This might yield biological modulators that could resist or delay aging of the skeletal muscles. More important, they might provide a pathway for reverting the currently incurable muscle dystrophies.
REFERENCES
- Akataki, K., K. Mita, M. Watakabe and K. Ito, 2002. Age related change in motor unit activation strategy in force production: A mechanomyographic investigation. Muscle Nerve, 25: 505-512.
Direct Link - Desaki, J., S. Oki and Y. Matsuda, 1996. Morphological changes of capillaries associated with muscle-fiber atrophy in the extensor digitorum longus muscle of aged rats. J. Electron Microsc., 45: 364-371.
Direct Link - Fayet, G., A. Rouche, J.Y. Hogret, F.M. Tom and M. Fardeau, 2001. Age related morphological changes of the deltoid muscle from 50 to 79 years of age. Acta Neuropathol., 10: 358-366.
Direct Link - Kadi, F., N. Charifi, C. Denis and J. Lexell, 2004. Satellite cells and myonuclei in young and elderly women and men. Muscle Nerve, 29: 120-127.
Direct Link - Karakelides, H. and N.K. Sreekumaran, 2005. Sarcopenia of aging and its metabolic impact. Curr. Top. Dev. Biol., 68: 123-148.
Direct Link - Kirkendall, D.T. and W.T. Garrett, 1998. The effects of aging and training on skeletal muscle. Am. J. Sports Med., 26: 598-602.
Direct Link - Kirkeby, S. and C. Garbarsch, 2002. Aging in different human muscles: An image analysis of histomorphometric characteristics of fiber types in human masseter and vastus lateralis muscles from young adults to the very old. Histol. Histopathol., 15: 61-71.
PubMedDirect Link - Klein, C., G. Marsh, R. Petrella and C. Rice, 2003. Muscle fiber number in the biceps brachii muscle of young and old men. Muscle Nerve, 28: 62-68.
Direct Link - Lexell, J., 1995. Human aging: Muscle mass and fiber type composition. J. Gerontol. A. Biol. Sci. Med. Sci., 50: 11-16.
PubMedDirect Link - Lorenzon, P., E. Bandi, F. Guarrini, T. Pietrangelo and R. Schafer et al., 2004. Ageing affects the differentiation potential of human myoblasts. Exp. Gerontol. 39: 1545-1554.
CrossRef - Maier, F. and A. Bornemann, 1999. Comparison of the muscle fiber diameter and satellite cell frequency in human muscle biopsies. Muscle Nerve, 22: 578-583.
Direct Link - Mattiello-Sverzut, A., L. Chimelli, M. Moura, S. Teixeira and J. De Oliveira, 2003. The effects of aging on biceps brachii muscle fibers: A morphological study from biopsies and autopsies. Arq. Neuropsiquiatr., 61: 555-560.
Direct Link - Melichna, J., C. Zauner, L. Havlichova, J. Novak, D. Hill and R. Colman, 1990. Morphologic differences in skeletal muscle with age in normally active human males and their well-trained counterparts. Hum. Biol., 62: 205-220.
PubMedDirect Link - Nikolic, M., D. Malnar-Dragojevic, D. Bobinae, S. Bajek, R. Jerkovic and T.S. Vranic, 2001. Age-related skeletal muscle atrophy in humans: An immunohistochemical and morphometric study. Coll. Anthropol., 25: 545-553.
Direct Link - Pansarasa, O., G. Felzani, J. Veccchiet and F. Marzatico, 2002. Antioxidant pathways in human aged skeletal muscle: Relationship with the distribution of type II fibers. Exp. Gerontol., 37: 1069-1075.
CrossRef - Parise, G., S.M. Phillips, J.J. Kaczor and M.A. Tarnopolsky, 2005. Antioxidant enzyme activity is up-regulated after unilateral resistance exercise training in older adults. Free Radic. Biol. Med., 39: 289-295.
CrossRef - Welle, S., C. Thornton, M. Statt and B. McHenry, 1996. Growth hormone increases muscle mass and strength but does not rejuvenate myofibrillar protein synthesis in healthy subjects over 60 years old. J. Clin. Endocrinol. Metab., 81: 3239-3243.
Direct Link