
Sheikh Arshad Saeed
Center for Molecular Medicine and Drug Research, International Center for Chemical Sciences, University of Karachi, Pakistan
Muhammad Ahmed Mesaik
Center for Molecular Medicine and Drug Research, International Center for Chemical Sciences, University of Karachi, Pakistan
Journal of Medical Sciences
Year: 2005 | Volume: 5 | Issue: 4 | Page No.: 350-357







ABSTRACT
Stem cell technology has emerged in the face of extraordinary advances for prevention, diagnosis and treatment of human diseases such as heart disease, diabetes, cancer and diseases of the nervous system, such as Parkinson’s disease and Alzheimer’s Disease (AD). It offers great promise for tissue regeneration, cell replacement and gene therapy, but clinical applications remain limited. Despite considerable ethical and practical barriers to their acceptance in the society, their potential in medicine is tantalising. There is a balance between the search for ways to regenerate bodies ravaged by age and disease on the one hand, with the need to ground these ideas in scientific fact and medical caution on the other. Stem cells are certainly exciting both as models of developmental biology and potential in human disease. Brain can be turned into blood, blood can be turned into brain, muscle, myocardium or liver and in some hands, bone marrow and neural stem cells can be turned into almost any type of cell. Such remarkable properties render it now feasible to contemplate isolating living healthy stem cells from the body, expanding them under cell culture conditions, combining them with biocompatible carrier molecules, directing their proliferation with growth factors and then transplanting them or their progeny into patients for clinical gain. In this way stem cells may, in future, be used to alleviate degenerative disorders, replace diseased or failing tissues with engineered substitutes and correct genetic disease. These living tissue substitutes may indeed overcome the drawbacks of classic transplantation and the limitations of standard treatments for degenerative disease, but their origins and potential are increasingly challenged by developments in science and medicine, politics and ethics.
PDF Abstract XML References Citation
How to cite this article
Sheikh Arshad Saeed and Muhammad Ahmed Mesaik, 2005. New Vistas in the Therapeutic Uses of Stem Cells. Journal of Medical Sciences, 5: 350-357.
DOI: 10.3923/jms.2005.350.357
URL: https://scialert.net/abstract/?doi=jms.2005.350.357
DOI: 10.3923/jms.2005.350.357
URL: https://scialert.net/abstract/?doi=jms.2005.350.357
INTRODUCTION
Stem cells are undifferentiated cells from the embryo, fetus, or adult that has the ability to self renew and exhibit multi-lineage differentiation. The first criterion requires each stem cell when it divides to generate at least one new identical stem cell, which is how these cells maintain their population unchanged over long periods of time[1]. They usually remain undifferentiated, but could be induced to differentiate into different cell types (Fig. 1).
Although stem cells are the primary units of development, they make up only a small proportion of total cells in the body. Stem cells are characterised by distinct sets of cell specific antigens and enzymes and expression of developmentally regulated genes. None of these markers is, however, unique for a particular stem cell type and thus the exact identity of stem cells in many organs remains elusive[2,3] (Fig. 2).
Stem cells have varying degrees of potential and are characterised as being totipotent, pluripotent or multipotent. Totipotency refers to the ability to form the embryo and trophoblast of the placenta. Pluripotent cells are derived from the early human embryo or from fetal gonadal tissue and have the potential to develop into any cell-type derived from the three embryonic germ layers: endoderm, mesoderm and ectoderm. Multipotent cells are sometimes said to be tissue restricted, meaning they exist within specific tissues and give rise to differentiated cells of that tissue only[4,5] (Fig. 3).
At least three differentiated stem cells have been identified: Embryonal Carcinoma (EC) cells, Embryonic Stem (ES) cells derived from the inner cell mass of blastocysts and Embryonic Germ (EG) cells obtained from post-implantation embryo[6]. First characterised in the mouse, ES and EG cells from humans can now be readily grown in the laboratory. ES cells are generally obtained from clinics and can generate most, if not all the 200 tissues or they are unable to give rise to the placenta, a property unique to the zygote[4,7-9]. The proof that ES cells differentiate along almost all cell lineages comes from introducing them into host blastocysts was done in the mouse, but not the human for obvious reasons.
Stem cells form the Central Nervous System (CNS) was first derived from fetal rat or mouse brain. They seem more limited in their differentiation potential compared with ES cells, but still give rise to the three major CNS cell types: neurons, astrocytes and oligodendrocytes. Neural Stem Cells (NSC) have also been isolated from the human adult CNS, although it is not clear whether they are multipotent, or have more restricted fates[10]. However, stem cells have been derived from almost all tissues in the body. Conventional wisdom held that these adult stem cells were developmentally committed, restricted to differentiate into cells from the brain form NSC and cells in the bone marrow give rise to blood stem cells. Although transdifferentiation is the presumed explanation, the environment dictates development of tissue-derived stem cells into a specific lineage, it is also possible that stem cells go through a step of de-differentiation to a more primitive state first and then re-differentiate[11].
Bone marrow has received the most attention as a source of stem cells for therapy because it contains mesenchymal and haemopoietic stem cells, both of which have the remarkable ability of migrating from their niches to sites of tassel growth and repair. Indeed, the best characterised stem cells are those responsible for haemopoisis. Four decades of work have defined the activities of bone marrow haemopoietic (Blood forming) Stem Cells (HSC), which are both self-renewing and multipotent in their ability permanently and clonally (from the single cell level) to produce all cell types of blood and the immune system in engrafted hosts. HSC are accessible as a source of stem cells and already proven safe in vivo; adult HSC transplants are commonly used to treat a variety of haematological diseases. Peripheral blood HSC can also be mobilized from human bone marrow after chemotherapy or administration of haemopoietic growth factors. MSC have been well characterised both by their properties in vitro and in vivo after transplantation into various animal models[11].
Stem cells may also be obtained from fetal blood and or tissues and from umbilical cord blood. Both HSC and mesenchymal (mesodermal origin) stem cells (MSC) have been isolated from and characterised in first trimester fetal blood, liver and bone marrow and resemble adult-derived cells in immunophenotype and differentiation capabilities[12]. There is some evidence that fetal stem cells are more plastic than adult stem cells and hence have greater therapeutic potential[13]. Umbilical cord blood is rich in HSC.
Uses of stem cells in research: These are some of the larger problems in basic and clinical biology where the use of stem cells might be the key to understanding.
Human developmental biology: The knowledge gained from human ES cells will impact many fields, which include cancer biology since it is now understood that many cancers arise by perturbations of normal developmental processes[14]. The availability of human ES cells will also greatly accelerate the understanding of the causes of birth defects and thus lead directly to their possible prevention[6].
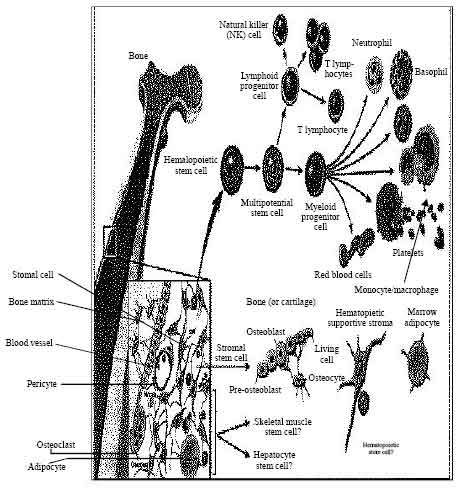 | |
| Fig. 1: | Hematopoietic and stromal stem cell differentiation |
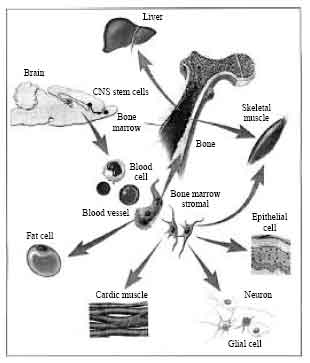 | |
| Fig. 2: | Preliminary evidence of plasticity among Nonhuman Adult Stem Cell This figure is taken from stem cells: scientific progress and future research direction. Report prepared by The NIH, USA, 2001 |
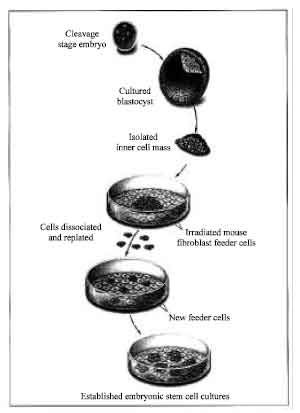 | |
| Fig. 3: | Techniques for generating embryonic stem cell cultures |
Models for human disease: Investigation of a number human disease is severely constrained by a lack of in vitro models. ES cells might provide cell and tissue types that will greatly accelerate investigation into HIV, HCV (which grow only in human or chimpanzee cells) and other viral diseases. Current animal models of neurodegenerative disease such as Alzheimer’s disease give only a partial representation of the disease’s process.
Transplantation: Pluripotent stem cells could be used to create an unlimited supply of cells, tissues, or even organs that could be used to restore function without the requirement for toxic immunosuppression and without regard tissue matching compatibility[15,16].Such cells, when used in transplantation therapies, would be suitable for universal donation. Bone marrow transplantation could become safe, cost effective and available for treating a wide range of clinical disorders, including aplastic anemia skeletal myocytes and hematopoietic diseases and certain inherited blood disorders[17,18].
However, these are some road blocks to therapeutic application of stem cells particularly in the field of transplantation which include the formation of the teratomas and teratocar cinemas and destruction of joints[19]. As well as cells to be transplanted has to display appropriate molecular identity on this functioned properly[20].
Gene therapy: In gene therapy, the genetic materials that provides a missing or necessary protein, or causes a clinically relevant biochemical process, is introduced into an organ for a therapeutic effect. For gene-based therapies, it is critical that the desired gene be introduced into organ stem cells in order to achieve long-term expression and therapeutic effect. However, still yet no bonafide success on that is due to proper delivery problems. The loss of expression or insufficient expression is another important limiting factor in successful application of gene therapy and could be overcome by transferring genes into stem cells (which presumably will then differentiate and target correctly)[21].
Clinical uses
Nervous system: The adult brain has limited capacity for repair. Cell death in brain disorders like stroke makes brain injury an ideal model for stem cell therapy. In mice, transplanting bone marrow-derived cells after stroke results in differentiated and co-localised cerebral endothelial cells and cells that express neuronal markers. While such studies do not provide evidence of functional neurons, the finding that stem cells can be used as a source of endothelial cells and neurons after brain injury shows promise for enhancing recovery from stroke[22].
Experience already exists with human fetal neural tissue transfer in neurodegenerative disorders. Many patients with Parkinson’s disease have received fetal tissue grafts with symptomatic improvement, but with unacceptable side effects. ES cells and NSC have both shown promise results in rat models of Parkinson’s, but immune rejection and excessive cell proliferation still remains a problem[23,24]. Instead, it is hoped that stem cells could be harvested from bone marrow and used to generate neurons for transplantation, or even administered systemically to find their way to the damaged brain. Early work suggests bone marrow-derived MSC can differentiate into glia and neurons but these cells have not yet been shown to have the electrophysiological characteristics of neurons[10] .
Spinal cord injury results not only in damage to axons, loss of neurons and glia, but also leads to secondary processes like ischaemia, anoxia and scar formation. Preserving even a small amount of tissue significantly improve functional recovery. A number of studies have demonstrated the successful use of multipotent NSC for demyelinating disease (e.g., multiple sclerosis)[25].
NSC can be engineered to produce therapeutic molecules and engraft in the brain: restricting their effect to the CNS without disturbing other neurobiological processes. In rat models of glioblastoma, engrafted cells that express interleukin-4 conferred a survival advantage on affected animals. This may lead to a new approach for gene therapy to brain tumours. Similarly, bone marrow-derived MSC may serve as vehicles for delivery of biological agents in tumours. MSC altered to express interferon-â have been shown to engraft at tumour sites in murine models after subcutaneous injectional and even to inhibit the growth of malignant cells[26].
The heart: The finding that stem cells from peripheral blood and bone marrow contribute to the formation of new blood vessels in adults in vivo, as well as the observation in heart transplant patients that stem cells from the recipient migrate into the donor organ, has encouraged new thinking on the mechanism of cardiac repair after injury[27]. Since normal repair injury, mobilisation of stem cells from elsewhere in the body and/or implantation of additional stem cells populations could confirm that implanted bone marrow stem cells improve function and perfusion in patients with previous myocardial infarction, as well as generating new tissue. Specialised subtypes of cardiomyocytes can be differentiated in vitro from ES cells and while currently only used as models for assessment of cardiac tissue and drug testing, are in future proposed as donor cell types for cardiac disease[28].
Skin: Tissues that can be engineered using stem cells now comprise a diverse range form epithelial surfaces to skeletal tissues. Tissue-engineered transplants are tissue-specific cells grown on a biocompatible carrier scaffold, with the aim of transplanting autologous cells to minimise graft versus host responses. Regeneration of skin is a particular challenge, as restoration of tissues that are characterised by high and continuous self-renewal requires extensively self-renewing stem cells. Successful transduction of epidermal stem cells has paved the way for the first trail of gene therapy for junctional epidermolysis bullosa (a rare skin disorder resulting in blistering to touch)[29,30].
The skeleton: With an increasing number of people suffering degenerative bone diseases (e.g., osteoarthritis), the demand for skeletal therapies is increasing. As with skin, the focus with bone has shifted from autologous grafts to multipotent stem cell strategies. In the simplest such procedure for bone reconstitution, MSC isolated from bone marrow would be expanded ex vivo, loaded onto an appropriate carrier and transplanted into or directed to the target tissue. Bioengineered scaffolds are effective in supporting bone regeneration, either alone or in conjunction with osteogenic growth factors[31].
Articular cartilage is adapted to withstand enormous mechanical stress, yet once destroyed it has only limited capacity for repair. Stem cells implanted into cartilaginous defects result in limited healing and tissue formation. Since some cartilage defects in vivo appear to heal partially after invasion of MSC from the bone marrow, therapeutic approaches now include using MSC to develop new cartilage and support cartilage repair. Delivery systems range from solid grafts of cells embedded in collagen sponges, cell-fibrin glue suspensions and cartilage cells transfected with adenoviral vectors carrying growth factors[32].
Liver: The prevalence of chronic liver disease and the shortage of donor livers have led to a search for suitable cell replacements for transplantation. The liver has indigenous populations of both functional and potential stem cells. In response to loss of liver tissue, hepatocytes behave like functional stem cells and restore the liver mass. In circumstances of prolonged liver damage or inhibition of hepatocyte repair, oval cells can act as facilitative stem cells. When the liver’s ability to regenerate has been overcome, hepatocytes can also be derived from bone marrow[33,34]. The ability of bone marrow HSC to cure a metabolic liver disease has already been shown in mice with type 1 tyrosinaemia (elevated blood levels of tyrosine, resulting in liver damage). In transplant patients, the degree of HSC engraftment into human liver varies according to the severity of liver damage, suggesting that local organ damage is necessary for significant stem cell recruitment into liver[35]. It is possible that an individual’s bone marrow stem cells are designed to mobilise during hepatic failure to increase the regenerative capacity of the liver[36].
Blood and bone marrow: There are three sources of HSC for allogeneic stem cell transplantation: bone marrow harvests, growth factor-mobilised Peripheral Blood (PB) collections and Umbilical Cord Blood (UCB) cells. HSC transplants are widely used for treatment of malignant haematological disorders, such as leukaemia, lymphoma, myeloma and breast carcinoma[37]. They have been more recently used for autoimmune disease, are also employed in the treatment of non-malignant disease including aplastic anaemia, β-thalassaemia and inborn errors of metabolism[38]. Critical properties of HSC for transplantation are not only their ability to engraft but also the rapidity and durability of their engraftment. The source of HSC influences the risk of developing Graft Versus Host Disease (GVHD). For example, the most rapid engraftment is observed with Peripheral Blood (PB) transplants, but they suffer from the adverse effects of a high lymphocyte dose and show an increase in chronic GVHE. In terms of overall survival after transplant, PB and bone marrow are comparable, although PB transplants may be advantageous in patients with advanced disease. HSC in UCB provide an alternative source of cells for transplantation, which has led to much interest in cord blood storage for future use[39]. Due to the limited number of HSC in a given cord blood unit, cord blood transplants are associated with slower haemopoietic engraftment, especially in larger recipients. So while dedicated donations of cord blood in at risk families continue and altruistic cord blood banks add to the number of samples available through bone marrow registries, routine directed collection of cord blood is not currently advocated given the difficulties of correct collection and storage and the low chance of a sample being used[40].
Infusions of bone marrow-derived MSC are under investigation to facilitate HSC engraftment[41]. MSC support haemopoisis in vitro and express cytokines that encourage bone marrow homing and long-term engraftment, suggesting that they may a have a role in improving success of HSC transplantation. Other advantages are that MSC do not express major histocompatibility antigens but possess immunosuppressive properties and are thus less likely to be rejected in vivo[42]. Durable engraftment of MSC has been demonstrated in immunocompetent animal models. Co-infusion of MSC with autologous blood stem cells in women with breast cancer receiving chemotherapy has a beneficial effect on haemopoietic engraftment[43].
Disease in the child: Osteogenesis imperfecta is a genetic disorder of mesenchymal cells characterised by defective type I collagen, the major structural protein in bone, for which there is no cure. For this reason and as osteogenic differentiation of MSC is well characterised, it was one of the first diseases for which stem cell-based therapy was proposed. Already, a few children with osteogenesis imperfecta have been transplanted with allogeneic bone marrow. Transplants engrafted in the genetically defective bone led to improved bone mineral content and growth velocity over three years of follow-up. Subsequent work has shown that additional therapy using MSC without marrow ablative treatment can boost responses in the host[44].
In the developing brain, stable clones of NSC contribute to normal brain development when injected into the germinal zones of newborn mice. Diffuse white matter loss in preterm brain injury in neonates is thought to be mediated through damage to oligodendrocyte precursors, leading to interest in their possible replacement by stem cells[45].
Duchenne muscular dystrophy is a progressive X-linked myopathy where the absence of dystrophin in muscle fibres eventually exhausts the muscle’s innate ability to regenerate, resulting in fibrosis. Infusion of muscle stem cells, bone marrow MSC and HSC, as well as transduced stem cells to deliver gene products, have all been proposed as treatment modalities. Skeletal muscle repair after infusion of bone marrow-derived stem cells has been variable, with engraftment ranging from undetectable to 10% of muscle fibres. More promisingly, MSC from human synovial membranes, which appear to be similar to adult bone marrow MSC, have been shown to engraft and differentiate into muscle. In murine muscular dystrophy models, their infusion induced synthesis of human dystrophin and decreased the number of abnormal muscle fibres, suggesting efforts should continue to develop MSC for postnatal or even prenatal therapy[46].
CONCLUSIONS
Stem cell research is a rapidly evolving field, as can be seen by the explosion of publications in the scientific literature and the focus of media attention. Just as gene therapy did 10 years ago, stem cell therapy promises widespread application in biomedical research and regenerative medicine, but many questions must be answered before it becomes a realistic prospect in clinical practice. Which stem cells will provide maximal tissue repair? What quantity of cells should be administered? How will stem cell therapy be delivered in order to ensure therapeutic benefit without harm? Which diseases should be treated with stem cells? Can chemokines, immune therapy, or pharmacological therapy enhance grafting and function of stem cell implants? Will stem cell therapy be applied early enough in the course of disease to change its natural history? How can we be sure that it is subject to overgrowth or carcinogenesis? What is the durability and longevity of stem cell implants and will these cells have the same requirements during growth and susceptibility to disease as the cells they replace? How safe is this therapy ultimately?
If the current peace of research continues, breakthroughs in stem cell research should soon offer the prospect of effective treatment for a wide range of human diseases and alter the way we think about the origin and regeneration of tissues and the development of disease. Their potential raises a number of scientific and ethical concerns and there is some way to go before stem cells are considered safe for their seemingly limitless range of clinical applications.
ACKNOWLEDGMENT
We thank Mr. Ghulam Rasool for expert editorial assistance.
REFERENCES
- Al-Hajj, M. and M.F. Clarke, 2004. Self-renewal and solid tumor stem cells. Oncogene, 23: 7274-7282.
CrossRefDirect Link - Alison, M.R., R. Poulsom, S. Forbes and N.A. Wright, 2002. An introduction to stem cells. J. Pathol., 197: 419-423.
CrossRefPubMedDirect Link - Baum, C.M., I.L. Weissman, A.S. Tsukamoto, A.M. Buckle and B. Peault, 1992. Isolation of a candidate human hematopoietic stem-cell population. Proc. Natl. Acad. Sci. USA., 89: 2804-2808.
Direct Link - Jiang, Y., B.N. Jahagirdar, R.L. Reinhardt, R.E. Schwartz and C.D. Keene et al., 2002. Pluripotency of mesenchymal stem cells derived from adult marrow. Nature, 418: 41-49.
CrossRefDirect Link - Campagnoli, C., I.A.G. Roberts, S. Kumar, P.R. Bennett, I. Bellantuono and N.M. Fisk, 2001. Identification of mesenchymal stem/progenitor cells in human first-trimester fetal blood, liver and bone marrow. Blood, 98: 2396-2402.
CrossRefDirect Link - Reya, T., S.J. Morrison, M.F. Clarke and I.L. Weissman, 2001. Stem cells, cancer and cancer stem cells. Nature, 414: 105-111.
CrossRefPubMedDirect Link - Ruiz-Arguelles, G.J., D. Gomez-Rangel, G.J. Ruiz-Delagado, A. Ruiz-Arguelles, B. Perez-Romano and L. Rivadeneyra, 2003. Results of an autologous noncryopreserved, unmanipulated peripheral blood hematopoietic stem cell transplant program: A single-institution, 10-year experience. Acta Haematol., 110: 179-183.