
Marla A. Endriga
lnstitute of Biology, University of the Philippines, Diliman- Quezon City, Philippines
Elmer-Rico E. Mojica
Institute of Chemistry, University of the Philippines, Los Ba�os, Laguna, Philippines
Florinia E. Merca
Institute of Chemistry, University of the Philippines, Los Ba�os, Laguna, Philippines
Marivic S. Lacsamana
Institute of Chemistry, University of the Philippines, Los Ba�os, Laguna, Philippines
Custer C. Deocaris
Gene Function Research Center, I-I-I Higashi, AIST Central 4: Tsukuba 305-8562, Japan
Journal of Medical Sciences
Year: 2005 | Volume: 5 | Issue: 1 | Page No.: 31-34







ABSTRACT
Ten lectins from different sources were screened for cytotoxic activity against Acanthamoeba sp. (a keratitis-causing amoeba) and Tetrahymena pyriformis (a ciliate). Lectins from Schefflera odorata (“lima-lima” plant) [molecular weight = 271 kDa], Swietenia macrophylla (large leaf mahogany) [molecular weight = 295 kDa] and Lenzites sp. (a mushroom) [molecular weight = 184 kDa] were found to possess high cytotoxic activity against the test organisms. These results indicate that lectins may be further exploited as potential chemotherapeutics against certain parasitic diseases.
PDF Abstract XML References Citation
How to cite this article
Marla A. Endriga, Elmer-Rico E. Mojica, Florinia E. Merca, Marivic S. Lacsamana and Custer C. Deocaris, 2005. Evaluation of Some Lectins as Anti-protozoal Agents. Journal of Medical Sciences, 5: 31-34.
DOI: 10.3923/jms.2005.31.34
URL: https://scialert.net/abstract/?doi=jms.2005.31.34
DOI: 10.3923/jms.2005.31.34
URL: https://scialert.net/abstract/?doi=jms.2005.31.34
INTRODUCTION
Lectins are carbohydrate-binding proteins or glycoproteins of non-immune origin that can agglutinate cells or precipitate glycoconjugates and polysaccharides[1]. Lectins have also been defined as proteins possessing at least one non-catalytic domain that binds reversibly to specific mono-or oligosaccharides[2]. However antibodies and proteins with enzymatic activity related to carbohydrates are not considered lectins[3]. Because of their chemical properties, lectins have become useful tools in several fields of biological research such as immunology, cell biology, membrane structure, cancer research and genetic engineering.
Lectins are present in a wide range of organisms from bacteria to animals, being present in all classes and families although not in all kinds and species[4]. With their binding capacity, they have the capability to serve as recognition molecules within a cell, between cells or between organisms and are assumed to play fundamental biological roles in different organisms.
In this study, the cytotoxic activity of ten lectins against Acanthamoeba sp. and Tetrahymena pyriformis was evaluated. Acanthamoeba sp. is an amoeba that caused keratitis, a serious and potentially devastating corneal infection generally seen in soft contact lens wearers[5], while the ciliated Tetrahymena pyriformis is a widely used as a test organism in the assessment of the cytotoxicity of different compounds and therapeutic agents[6]. Results from this study can be used as preliminary data in the possible utilization of lectins against pathogenic microorganisms such as those that are found in our water supply.
MATERIALS AND METHODS
Isolation of lectins: The lectins used in the experiment were isolated from different organisms listed in Table 1. These were isolated during the period 2000-2003 using the method of Pisueña et al.[7].
Microorganisms: Acanthamoeba were grown at 28°C on Monoxenic Non-nutrient Agar (MNA) medium and trophozoites collected from 24 to 48 h old cultures. The agar surfaces were flooded with 5 mL of PBS and gently scraped with an inoculating loop. Trophozoites were harvested from suspension by centrifugation at 350xg for 10 min. The supernatant was aspirated out and the sediment was washed twice in PBS. The trophozoites in the resultant suspension were optically counted in a hemocytometer, adjusted to a final concentration of 104 amoeba per mL and used immediately for testing.
| Table 1: | Sources of lectin |
 | |
On the other hand, Tetrahymena pyriformis were grown axenically at 25°C in tissue culture flasks to a density of approximately 105 protozoa per mL in 1% proteose peptone medium enriched with 0.1% yeast extract. The protozoa were then centrifuged at 200xg for 1 min. The supernatant was diluted with sterile distilled water to a final density of 104 cell per mL.
Anti-protozoal assay: The purified lectins were then tested for anti-protozoal properties against Acanthamoeba sp. and Tetrahymena pyriformis using microplate-based in vitro bioassays. This was done by mixing 50 μL of cell suspension with the same volume but varying concentrations of the purified lectin in microtiter plate wells. Mortality of the cultured cells was examined under a microscope 1 and 24 h after addition of the lectin. Ara-C and Etoposide were used as positive controls while sterile distilled water served as negative control.
RESULTS
All lectins, with the exception of those from marine invertebrates, showed activity against the protozoans at 500 ppm. The growth of both protozoans upon exposure to the lectins was inhibited after 24 h of incubation. Lenzites sp. lectin was found to be active against both protozoans after one hour of incubation. These were also observed in the two seed lectins, where C. papaya lectin exhibited activity against Tetrahymena pyriformis while A. blancoi lectin showed activity against Acanthamoeba (Table 2).
For the Acanthamoeba, lectins sourced from the leaves of S. macropylla and S. odorata, notably both medicinal plants, showed the most potent activity with lectin from the former inhibiting the growth of cells at a concentration as low as 25 ppm (Table 3). Fungi lectins and seed lectins also inhibited the growth of Acanthamoeba cells at concentrations higher than the lectins from medicinal plants.
The minimum concentration of lectins needed to inhibit the growth of Tetrahymena pyriformis is lower than that for Acanthamoeba. Lectins from the medicinal plants S. odorata and S. macrophylla and from Lenzites sp. showed activity at concentrations as low as 10 ppm (Table 4).
| Table 2: | Preliminary assay of the cytotoxic activity of different lectins against Acanthamoeba sp. and Tetrahymena pyriformis |
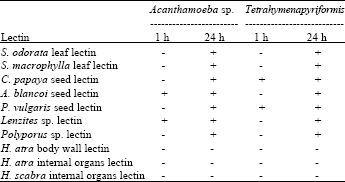 | |
| Table 3: | Results of antiprotozoal test against Acanthamoeba sp. |
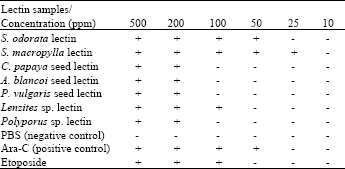 | |
| Table 4: | Results of antiprotozoal test against Tetrahymena pyriformis |
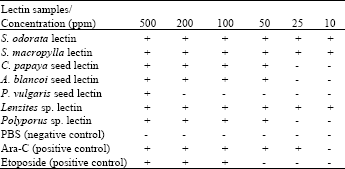 | |
| + inhibition in cell growth observed, - no inhibition in cell growth observed | |
The other lectins, with the exception of P. vulgaris seed lectin, showed cytotoxic activity against T. pyriformis at much lower concentration than in Acanthamoeba. P. vulgaris seed lectin appeared more active against Acanthamoeba than against T. pyriformis. In addition, some lectins showed stronger or the same level of activity in comparison to Ara-C and Etoposide, cancer therapeutic agents that were used as positive control (Table 4).
DISCUSSION
Few studies on antiprotozoal agents from plant-based materials have been done. Of those that have been reported, many used Entamoeba histolytica as test organism. Some agents have been tested against Acanthamoeba and extracts from plants such as Ipomoea sp., Kaempferia galanga and Cananga odorata were found to be active against three species of this protozoan.
Almost all antiprotozoal agents known are therapeutic agents synthesized in the laboratory. These include ivermectin, fungizone, pentamidine and Distamycin A[8-11], which were found active against Acanthamoeba. For Tetrahymena pyriformis, the cytotoxicity of several therapeutic agents like cyclosporin-A, cisplatin and doxorubicin[12] has also been assessed. The toxicity of non-ionic surfactants such as Triton X-100[13], some tricyclic antidepressants[14] and food additives[15] like the food color tartrazine, the preservatives sodium nitrate and sodium benzoate and the antioxidant BHT was also evaluated using Tetrahymena pyriformis. In this study, lectins were found to exhibit different levels of cytotoxic activity. The activity of the purified lectins could be due to their interaction with sugars present in the protozoans. The carbohydrate-lectin interactions are known to play important, broad roles in cell recognition, adherence, cell division, among others, in different organisms. It has been reported that Concanavalin A can bind both to the plasma membrane and to intracellular structures of Tetrahymena pyriformis[16]. Binding of lectins with some sugars in the protozoans could cause interference in chemical or biological processes that eventually lead to the death of these microorganisms.
Lectins were found to exhibit cytotoxic activity at varying concentrations against Acanthamoeba sp. and Terahymena pyriformis. Results indicate that lectins may be exploited as potential chemotherapeutic agents against certain parasitic disease and that it might be worthwhile looking into the potential activity of lectins against other microorganisms, particularly those that cause disease.
ACKNOWLEDGMENTS
We extend our heartfelt thanks to Rowena Quizon, Mary Princess Ballatan, Mary Dawn A. Celiz, Bethsaida M. Alpante and Melvelyn David for the initial isolation of some of the lectin samples used in the study and the Institute of Biology, UP Diliman and St. Luke’s Medical Center for the microorganisms.
REFERENCES
- Goldstein, I.J., R.C. Hughes, M. Monsigny, T. Osawa and N. Sharon, 1980. What should be called a lectin? Nature, 285: 66-66.
CrossRef