
Mohamed A. Al-Omar
Department of Pharmaceutical
Chemistry
College of Pharmacy
King Saud University P.O. Box 2457, Riyadh 11451
Saudi Arabia
Journal of Medical Sciences
Year: 2005 | Volume: 5 | Issue: 3 | Page No.: 163-168







ABSTRACT
2-hydroxy-1,4-naphthoquinone (HNQ; Lawsone, CAS 83-72-7) is the main dye ingredient found in the natural plant of Henna (Lawsonia inermis). The percentages of superoxide anion (O2.) and hydrogen peroxide (H2O2,) formation during the oxidation of 100 μM phenanthridine by guinea pig aldehyde oxidase have been measured and found to be 6-10% and 85-90%, respectively. The effect of HNQ on the initial rates of phenanthridine oxidation, hydrogen peroxide formation and superoxide anion with aldehyde oxidase was investigated and compared with those of xanthine oxidase. It was found that HNQ is a specific inhibitor of aldehyde oxidase in vitro. In this study, HNQ inhibits the production of superoxide anion and substrate oxidation more potently than hydrogen peroxide. Thus, the site of interaction is thought to be flavin semiquinone, FADH.. The IC50 value of HNQ with phenanthridine oxidation by aldehyde oxidase is 9.3±1.1 μM, which in excess of 15 fold of maximal plasma concentrations (Cmax) of HNQ, indicating a high degree of safety margin. The interaction of HNQ with aldehyde oxidase has been also investigated using different electron acceptors including, oxygen, potassium ferricyanide and cytochrome c and found to be equipotent.
PDF Abstract XML References Citation
How to cite this article
Mohamed A. Al-Omar, 2005. Effect of 2-hydroxy-1, 4-naphthoquinone, a Natural Dye of Henna, on Aldehyde Oxidase Activity in Guinea Pig Liver. Journal of Medical Sciences, 5: 163-168.
DOI: 10.3923/jms.2005.163.168
URL: https://scialert.net/abstract/?doi=jms.2005.163.168
DOI: 10.3923/jms.2005.163.168
URL: https://scialert.net/abstract/?doi=jms.2005.163.168
INTRODUCTION
2-hydroxy-1, 4-naphthoquinone (HNQ; Lawsone, Fig. 1) is the principal natural dye ingredient in the leaves of Henna (Lawsonia inermis). In Eastern countries, Henna has been used for long time as a hair dye and tattoo. In fact, HNQ is also widely used and has become popular in Western countries in pure and natural forms due to its natural origin.
Each 20 g of Henna corresponding to approximately 300 mg of pure HNQ. There are very few published studies on HNQ toxicity and safety. The genetic and general toxicity of Henna and HNQ were recently reviewed by Kirkland and Marzin[1] and by Nohynek et al.[2]. They concluded that HNQ has a complex toxicity and genetic toxicity profile. On the basis of these data, HNQ was considered to be toxic in vivo.
In view of the wide use of Henna in Saudi Arabia and all over the globe, it is necessary to conduct an investigation for its interaction with body metabolic pathways. Interestingly, HNQ is structurally related to menadione (Fig. 1), a potent and selective inhibitor for aldehyde oxidase (EC 1.2.3.1) [3,4]. Menadione is used in vitro to characterize aldehyde oxidase activity in both oxidation and reduction reactions and with aldehyde oxidase from different species[5].
There is paucity of literature on HNQ, therefore the present study investigate its interaction with aldehyde oxidase, which catalyzes the oxidation of aldehydes and N-heterocyclic compounds to the corresponding carboxylic acids and lactams, respectively and belong to the xanthine oxidase family, a family of molybdenum (Mo)-containing hydroxylase[3-5]. Aldehyde oxidase generates superoxide and hydrogen peroxide but in contrast to xanthine oxidase, aldehyde oxidase seems to be a permanent oxidase, with no activity towards NAD+. Studies on aldehyde oxidase and xanthine oxidase have shown that modulation of enzyme activities, cofactor availability, substrate concentration and oxygen tension all affect rates of intracellular ROS production[3-5].
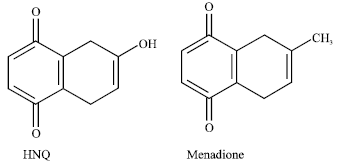 | |
| Fig. 1: | Chemical structures of lawsone (2-hydroxy-1,4-naphthoquinone; HNQ; CAS 83-72-7) and Menadione (2-methyl-1,4-naphthoquinone; vitamin K3; CAS 58-27-5) |
The aim of the present research was to investigate the interaction of HNQ with partially purified guinea pig liver molybdenum hydroxylase in term of superoxide anion production, hydrogen peroxide formation and the overall substrate oxidation.
MATERIALS AND METHODS
Instrumentation: Enzyme activity was determined spectrophotometrically using a Cary 50 UV/VIS spectrophotometer (Varian Australia Pty Ltd., Mulgrave/Victoria, Australia), which was linked to a cell temperature control unit. With the exception of enzyme, which was kept in ice until mixing with other components, all solutions were pre-warmed to 37°C. The spectrophotometer was computer-controlled by Carry WinUV® spectroscopy software package with additional kinetics software (2002).
Reagents and chemicals: All reagents and solvents are of analytical grade. All chemicals and reagents were purchased from Sigma-Aldrich Chemical Company Ltd. (Poole, Dorset, UK). Cytochrome c (from horse heart), horseradish peroxidase (HRP, type VI) and superoxide dismutase (SOD, from bovine liver suspension in 3.8 M (NH4)2SO4, pH 7.0) were also purchased from Sigma Chemical Company Ltd. (Poole, Dorset, UK).
Animals care: Dunkin-Hartley guinea pigs (600-700 g) were inbred and housed in pairs in plastic cages under a cycle of 07:00-21:00 h light and 21:00-07:00 h dark daily. They were given free access to a diet of FD1 guinea pig pellets supplied by Special Diet Services, hay, twice weekly and water ad libitum. They were maintained at a temperature of 18-19°C and humidity at 50%. The animals were killed by cervical dislocation between 10:00 am and 11:00 am daily.
Sorenson’s phosphate buffer (67 mM), pH 7.0: Sorenson’s phosphate buffer (67 mM) was made from two separate solutions containing either 9.511 g L-1 Na2HPO4 or 9.118 g L-1 KH2PO4 in distilled water. The buffer was made by adding 611 mL of Na2HPO4 solution to 389 mL of KH2PO4 solution and the pH was adjusted to pH 7.0 with 0.3 M phosphoric acid, using a pH-meter. EDTA (0.0372 g) was added to one liter of above mixture to give a final concentration of 100 μM EDTA. The buffer was stable for at least one month at 4°C.
Preparation of partially purified molybdenum hydroxylases from guinea pig liver: Aldehyde oxidase was partially purified from liver homogenate of mature Dunkin-Hartley guinea pigs following a published methodology[6]. Partially purified enzyme was stored in liquid N2 until needed.
Determination of initial oxidation rates and production of superoxide anion: Aldehyde oxidase activity in partially purified molybdenum hydroxylase fractions was monitored, at 37°C, using 100 μM 2-pyrimidinone, 50 μM phenanthridine, 100 μM phthalazine (enzyme fraction was diluted, 1:10) and 100 μM indole-3-aldehyde (1:40 dilution) as substrates in 67 mM Sorenson’s phosphate buffer, pH 7.0, containing 100 μM EDTA. Enzyme activity of guinea pig liver molybdenum hydroxylase fractions was also measured using 50 μM xanthine as a substrate. The initial activity of guinea pig liver molybdenum hydroxylase fractions was also measured in the presence of 1, 10, 40 and 100 μM HNQ. The initial velocity for substrate oxidation was determined by measuring the change in absorbance/minute and calculating enzyme activities in μmol/min/mg protein in the presence and absence of HNQ. The following equation was used to calculate the enzyme activity:
 |
Reduction of cytochrome c by partially purified molybdenum hydroxylase fractions at 550 nm was followed using phthalazine, indole-3-aldehyde, 2-pyrimidinone, phenanthridine and xanthine as substrates. Superoxide anion reacts with ferricytochrome c reducing Fe(III) to Fe(II).
Fluorimetric determination of hydrogen peroxide formation during molybdenum hydroxylase-catalyzed oxidation: Hydrogen peroxide formation was monitored fluorimetrically, using an MPF-3 Fluorescence Spectrophotometer with the excitation wavelength set at 395 nm and the emission wavelength set at 470 nm. The slit width was fixed at 6 mm for both excitation and emission, using a 150-watt Xenon Lamp as the light source. Hydrogen peroxide generated during molybdenum hydroxylase-catalysed oxidation was reacted with horseradish peroxidase to form a complex (compound I). The complex causes the oxidation of the fluorophore, scopoletin, resulting in a decrease in fluorescence, which is proportional to original hydrogen peroxide content as described previously[7].
Protein determination for partially purified molybdenum hydroxylase fractions: A Pierce Bicinchoninic acid (BCA®) protein reagent assay kit was used. This technique depends on the production of Cu (I) from the reaction of protein with Cu (II) in an alkaline medium as described by Smith et al.[8].
RESULTS AND DISCUSSION
Effect of HNQ and other inhibitors on substrate oxidation: Aldehyde oxidase and xanthine oxidase are both present in guinea pig partially purified molybdenum hydroxylase fractions; accordingly, specific enzyme inhibitors were used to confirm the specificity of the spectrophotometric assay. Chlorpromazine, menadione were used as specific aldehyde oxidase inhibitors[5,9]. Under the conditions used in this study, these inhibitors usually inhibit substrate oxidation by 98-99%[9]. In the present study, 100 μM of chlorpromazine and menadione decreased initial oxidation rates of 100 μM phthalazine, indole-3-aldehyde, 2-pyrimidinone and 50 μM phenanthridine by 97-99% (p<0.001). In contrast, 100 μM allopurinol, a xanthine oxidase inhibitor, caused minimal reduction of substrate oxidation (2-4%)[10]. As it has been shown that allopurinol is slowly converted by aldehyde oxidase to oxipurinol, it though that allopurinol is a competitive substrate of aldehyde oxidase[10]. HNQ was tested as an inhibitor for aldehyde oxidase and found to be a good inhibitor. The effect of HNQ on substrate oxidation has been compared to those of traditional aldehyde oxidase inhibitors (Table 1). HNQ was incubated with the enzyme preparation using oxygen as electron acceptor and the incubation mixture was monitored by repetitive scanning between 200-700 nm for up to 10 min. There were no change observed in the spectrum of HNQ. It was therefore concluded that HNQ is not a substrate for guinea pig liver aldehyde oxidase. Interestingly, HNQ caused negligible inhibition (4%) of xanthine oxidation. This is strongly indicates that HNQ is a specific aldehyde oxidase inhibitor (94-96%) with very little effect on xanthine oxidase (4%). In fact, HNQ is in close resemblance with menadione (Fig. 1).
Electrons egress from the enzyme can be followed using electron acceptors that interact with the enzyme at different redox centers. Potassium ferricyanide (K3Fe(CN)6) accepts electrons from iron-sulfur center whereas cytochrome c is reduced by superoxide anion at FAD site. Inhibition of potassium ferricyanide and cytochrome c reduction during substrate oxidation by aldehyde oxidase was found to be compared to that of oxygen (Table 2). Which may indicate that HNQ inhibits the enzyme at other sites than molybdenum center. However, the inhibition of reactive oxygen species formation may shed more light on the specific-site of inhibition.
| Table 1: | Effects of HNQ, potent aldehyde oxidase and xanthine oxidase inhibitors on the oxidation of phthalazine, indole-3-aldehyde, 2-pyrimidinone and phenanthridine catalyzed by partially purified guinea pig liver molybdenum hydroxylase fractions |
 | |
| * Results are expressed as mean percentage inhibition±SD (male guinea pig, n = 3), ** Control rates (μmol/min/mg protein) | |
| Table 2: | Effects of HNQ and menadione, a potent aldehyde oxidase inhibitor, on the oxidation of phenanthridine catalyzed by partially purified guinea pig liver molybdenum hydroxylase fractions using different electron acceptors |
 | |
| Table 3: | Effects of HNQ, chlorpromazine and menadione on the reactive oxygen species (ROS) formation during phenanthridine oxidation catalyzed by partially purified guinea pig liver molybdenum hydroxylase fractions using different electron acceptors |
 | |
| * The results are expressed as mean percentage inhibition±SD (n = 4) **The control rates for superoxide anion and hydrogen peroxide formation are 0.054 and 0.185 μmol/min/mg | |
Effect of HNQ on reactive oxygen species formation: During aldehyde oxidase catalysed-reactions, reactive oxygen species are produced in substantially high amounts as reviewed recently[11]. The effect of HNQ on production of reactive oxygen species, hydrogen peroxide and superoxide anion, has been compared to those of chlorpromazine and menadione during the oxidation of 50 μM phenanthridine (Table 3).
The effect of HNQ on superoxide anion production (96%) was more pronounced than that of hydrogen peroxide (79%). Similarly, menadione inhibits the superoxide anion production more than hydrogen peroxide. In contrary, chlorpromazine has been found to be equipotent on both species. Interaction of menadione is thought to occur at FAD site[3,12], which is consistent with the ability of menadione to act as an electron acceptor of xanthine oxidase. As a result, the oxidation rates of xanthine oxidase are enhanced in the presence of menadione[12,13].
In this study, HNQ was found to be a selective inhibitor of guinea pig liver aldehyde oxidase with little reactivity towards xanthine oxidase. As HNQ inhibits superoxide anion production more effectively than hydrogen peroxide production, the site of interaction is thought to be flavin semiquinone, FADH.. The proposed site of HNQ interaction during phenanthridine oxidation by aldehyde oxidase is shown in Scheme 1.
By analogy with the reaction between xanthine and xanthine oxidase/dehydrogenase, it can be postulated that substrate oxidation occurs at the molybdenum center reducing Mo(VI) to Mo(IV)[14]. This is thought to be the rate-limiting step in substrate oxidation[15]. It is likely that chlorpromazine inhibits the enzyme at this stage as similar effects are observed on substrate oxidation, hydrogen peroxide production and superoxide anion formation. However, in view of non-competitive nature of chlorpromazine inhibition, it appears that chlorpromazine does not bind to fully oxidized aldehyde oxidase but to a reduced form molybdopterin cofactor containing either Mo(IV) or Mo(V) Scheme 1-A[16].
Rapid intra-molecular electron transfer will generate reduced Fe/S and FADH. (flavin semiquinone, Scheme 1-B). Although electrons are transferred singly from Mo(IV), under steady state conditions the flavin is maintained as FADH2 (flavin hydroquinone, Scheme 1-D), which favors reduction of molecular oxygen to hydrogen peroxide. During enzyme turnover, aldehyde oxidase is thought to cycle between four-electron and two-electron reduced forms thus both Fe/S groups will be reduced. Rapid equilibria will be established between FAD/FADH. and FADH./FADH2. The major flow of electrons will be through the four-electron reduced form containing FADH2 (Scheme 1-D) whereas electron flux from FADH. to form superoxide anion has been shown in this study and other studies to account for only 6-10% of total substrate turnover[17].
In summary, HNQ appears to bind to FADH. as the effect on superoxide anion formation is more pronounced than that on hydrogen peroxide formation (Scheme 1-B).
It is noteworthy that topical administration of Henna produces CMAX values of less than 100 ng mL-1 (= 0.6 μM) of HNQ[2] while, this study indicates that IC50 value of HNQ with phenanthridine oxidation by aldehyde oxidase is 9.3±1.1 μM (Fig. 2, for comparison the IC50 value for menadione is 9.6±1.5 μM). Given that toxicokinetic-base safety margins in excess of 15-fold are considered to indicate a high degree of safety and taking into account that humans are exposed only intermittently and on a far small proportion of their body surface (700 cm2 or 4% of the body surface area), these results indicate that hair dyeing with Henna or HNQ poses no or negligible risk to human health.
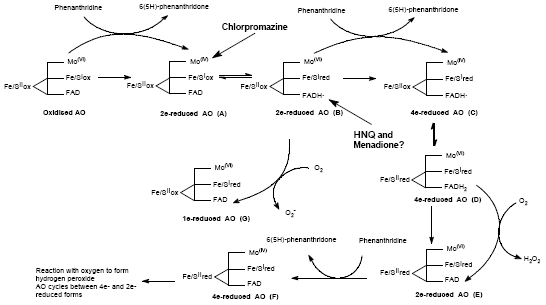 | |
| Scheme 1: | Reduction of aldehyde oxidase by phenanthridine and subsequent electron transfer to form superoxide anion and hydrogen peroxide in the presence of different inhibitors (AO: aldehyde oxidase; HNQ: 2-hydroxy-1,4-naphthoquinone; Fe/S: iron-sulfur center) |
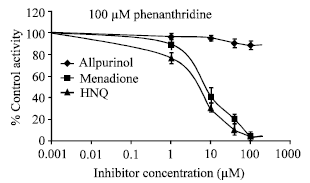 | |
| Fig. 2: | IC50 determinations for the inhibition of phenanthridine oxidation by guinea pig liver aldehyde oxidase in the presence of (1, 10, 40 and 100 μM) allopurinol, menadione and HNQ (n= 3±SD), the control values as indicated in Table 1 |
This study also investigated the site of interaction between HNQ and aldehyde oxidase and thought to be conducted at FADH.. The effect of HNQ on reduction reaction of aldehyde oxidase may shed more light on the role of HNQ on reactive oxygen species production. This was not investigated in the present study but will be focused on in further studies.
ACKNOWLEDGMENT
The author would like to thank Department of Pharmaceutical Chemistry, College of Pharmacy, King Saud University for the financial support and facilities.
REFERENCES
- Al-Omar, M.A., C. Beedham, F. Belal, J.A. Smith and A.A. El-Emam, 2005. Fluorimetric measurement of hydrogen peroxide produced during aldehyde oxidase catalysed oxidation using scopoletin. J. Med. Sci., 5: 10-20.
CrossRefDirect Link - Smith, P.K., R.I. Krohn, G.T. Hermanson, A.K. Mallia and F.H. Gartner et al., 1985. Measurement of protein using bicinchoninic acid. Anal. Biochem., 150: 76-85.
CrossRefPubMedDirect Link - Al-Omar, M.A., C. Beedham and I.A. Al-Sarra, 2004. Pathological roles of reactive oxygen species and their defence mechanisms. Saudi Pharm. J., 12: 1-18.
Direct Link