
F.O. Obi
Department of Biochemistry
Faculty of Science
University of Benin
Benin City, Edo State, Nigeria
M.A. Adaikpoh
Department of Biochemistry
Faculty of Science
University of Benin
Benin City, Edo State, Nigeria
Journal of Medical Sciences
Year: 2004 | Volume: 4 | Issue: 2 | Page No.: 90-94







ABSTRACT
The effects of orally administered doses of Congo Red (CR) on rat testes weight, DNA and protein content as well as acid and alkaline phosphatase activities have been examined. Following 6 weekly exposure of group of rats to 5, 10, 20, 30 and 40 mg CR kg-1 body weight there was significant loss in testis weight of rats exposed to ≥20 mg CR kg-1 bd wt relative to the CR-free control rat testis (Control: 1.116±0.010 g; test (20 mg): 1.033±0.082 g). When compared to the control, the DNA obtained from the testis of rats exposed to ≥10 mg CR was statistically significantly lower (Control: 89.67±0.58 μg DNA g-1 tissue: test (10 mg) 84.50±1.00 μg DNA g-1). The protein content, the acid and alkaline phosphatase activities of the testes from CR-exposed rats did not differ significantly from that of CR-free control rats. Strong and significant correlation was demonstrated between the changes in testis weight and CR-dosage(r=-0.98; p<0.01), between its DNA content and CR-dosage (r =-0.77; p<0.10) and between testis weight change and changes in DNA level (r = +0.81; p<0.05)
PDF Abstract XML References Citation
How to cite this article
F.O. Obi and M.A. Adaikpoh, 2004. Gravimetric and Biochemical Effects of Orally Administered Congo Red Dye on Rat Testes. Journal of Medical Sciences, 4: 90-94.
DOI: 10.3923/jms.2004.90.94
URL: https://scialert.net/abstract/?doi=jms.2004.90.94
DOI: 10.3923/jms.2004.90.94
URL: https://scialert.net/abstract/?doi=jms.2004.90.94
INTRODUCTION
Azo dyes make up a large proportion of manufactured dyestuffs. They are primarily used as colourants in food, textile, leather, paper and plastic industries [1]. A number of them such as Congo red (Direct Red 28, colour index # 22120) and Chlorazol black E (Direct Black 38, colour index # 30235) are based on benzidine. Others are based on benzidine congeners, dimethylbenzidine and dimethoxybenzidine[2].
It has been reported that in mice and rats, prenatal exposure to the dye Congo red permanently reduces the number of germ cells in male and female offspring. Also that it causes infertility in female mice following fetal exposure[2,3]. Further studies subsequently revealed that only benzidine-based dyes alter testicular development and hypospermatogenesis during adulthood following gestational exposure[2]. These and several earlier studies on the reproductive toxicity of azo dyes were based on histological, gravimetrical and anatomical evaluations[4-6]. Hence there is paucity of information on the biochemical changes associated with the gonadal toxicity of azo dyes. It was therefore considered necessary to investigate the effect of CR not only on the weight of testes but also on any associated biochemical changes in it when the dye is administered orally and directly to male rats.
MATERIALS AND METHODS
Treatment of animals and dye administration: Six-weeks old male Wistar rats used for the experiments were obtained from Lab Animal Services, LUTH, Lagos, Nigeria. They were divided into 6 groups of 3-4 rats each and housed in clear wire cages (33x20x13 cm) on wood shaving. They were allowed free access to water and commercial rat pellets (Pfizer, Nigeria Plc) and left to acclimatize to our laboratory conditions for 14 days before the commencement of the experiment. Congo red (E. Merck AG, Darmstadt) in water (5, 10, 20, 30 and 40 mg kg-1 body weight of rats in groups 2, 3, 4, 5 and 6, respectively) was given in not more than 0.5 ml water by gavage once a week for 6 weeks. Control rats (group 1) were given 0.5 ml H2O kg-1 body weight. After 6 weeks of administration rats were necropsied under chloroform (BDH, Poole, England) anaesthesia. The necropsy included saving and weighing of the testes and subsequent storage at -20°C until required for biochemical examination
Biochemical analyses
Extraction and quantitation of DNA and protein: DNA and protein were extracted from 0.5 g of testis as described by Yates et al.[7]. 20% homogenate of testis was prepared using ice cold 6% Trichloroacetic acid (TCA) and centrifuged at 960xg for 2 min. The residue was washed twice with 1 ml of 6% TCA, once with 1 ml ethanol/diethyl ether mixture (3:1v/v) and twice with 1 ml of diethyl ether. The acid insoluble defatted residue was resuspended in 1 ml of 0.3M NaOH and incubated for 18 h at 37°C. 2 ml of ice cold 50% (w/v) TCA was added and the mixture chilled by placing on ice for 30 min. The precipitate (DNA and Protein) was collected by centrifugation, 960xg for 10 min. The precipitate was suspended in 1 ml TCA and then heated at 95°C for 15 min to dissolve. The solution was cooled and centrifuged at 960xg for 5 min. The supernatant was analysed for DNA content while the precipitate was recovered for Protein estimation. The DNA content was estimated spectrophotometrically by measuring the absorbance at 260 and 320 nm, respectively (Milton Ray Unican SP1800 UV spectrophotometer). The absolute amount of DNA in each extract was calculated by using the relationship-DNA (50 μg ml-1) = one absorbance unit at A’260; where A’260 = A260-A320. reagent.
The quantity of protein extracted and in the homogenates (see below) was determined by using biuret reagent. To 0.4 ml of test sample in a test tube, 0.6 ml of distilled water was added. In another test tube, the blank, 1 ml distilled water was added. Three ml of 1.5M NaOH and 3 ml biuret reagent were then added to both tubes. The contents were mixed by swirling, allowed to stand for 30 min and the absorbance read at 540 nm.
Assay of acid and alkaline phosphatase: The activity of acid phosphatase (ACP) in 20% testis homogenate in citrate buffer, pH 4.85 was determined by the method of Bodansky[8]. Two replicates per sample were prepared for the assay. These were put in two separate test tubes labeled CT. Another test tube labeled CB served as blank. 1 ml of buffered substrate [prepared by mixing 1 part of 0.09 mol L-1 citrate buffer (pH 4.9) with 1 part of 4.0 mg ml-1 stock substrate solution of p-nitrophenyl phosphate (pNPP )] was then added to all the test tubes. The contents of the tubes were mixed thoroughly. 0.09 ml of the homogenate was added to the CT tubes. The contents of the test tubes were mixed by swirling and allowed to stand for 30 min. at 37°C. 4 ml of 0.1M NaOH solution was added to all the tubes to stop further enzyme reaction. 0.09 ml distilled water was added to the CB tubes and the absorbance read at 410 nm. Standard calibration curve was prepared by diluting 1, 2, 4, 6, 8 and 10 ml of 0.5 mmol ml-1 p-nitrophenol (pNP), with water to a volume of 10 ml. The instrument blank consists of 10 ml of water. 1.1 ml of 0.2M NaOH was added to all tubes and the absorbance read at 410 nm. From the calibration curve the activity of ACP in each sample was calculated.
Alkaline phosphatase (ALP) was assayed using the method described by Plummer[8] after preparing a 20% homogenate of testis in sodium carbonate/bicarbonate buffer, pH 10.2. Two replicates per sample were prepared for the assay. To each of them, 2.5 ml of the buffer was added. 0.3 ml of buffered substrate (0.63 mmol ml-1) was added to give a final substrate concentration of 0.063 M in the reaction mixture. The mixture was brought to a final volume of 3 ml with the addition of 0.2 ml of the diluted tissue homogenate to initiate the reaction. The mixtures were incubated at 37°C for 10 min. The reaction was stopped by the addition of 1 ml of 0.1M NaOH solution and left standing on ice for 10 min. The content of each tube was centrifuged for 5 min at 3000 rpm. The absorbance was read against the blank at 430 nm. Calibration curve was prepared by diluting 0.02, 0.04,0.06, 0.08 and 0.10 ml of standard pNP solution (555 ug ml-1) with water to a volume of 0.10 ml. The instrument blank consists of 0.10 ml of water. 4.9 ml of the alkaline buffer was added to all tubes, mixed and absorbance was read at 430 nm. ALP activity was estimated based on the calibration curve.
Statistical analysis: The data are presented as means±SD. The mean values were compared using ANOVA and Least square difference test. Correlation coefficient was determined by using the Pearson product-Moment statistical analysis[10].
RESULTS AND DISCUSSION
We administered water or CR orally once a week to control and CR-exposed male rats respectively and observed the effects of these treatments on the testes of the animals at the end of 6 weeks.
Relative to the control the weight of testis from rats exposed to 20 mg CR and above were significantly lower than that of the control rats (Table 1). Compared to the control the DNA obtained from the testis of rats exposed to 10 mg CR and above was significantly lower. Changes in protein content of testis from CR-exposed rats were not statistically significantly different from the CR-free control value (Table 2). Relative to the control values, no significant change in activity of both ACP and ALP was observed in the CR-exposed rat testis (Table 3).
The weight of all test rat testes, except those of groups 2 and 3, was significantly reduced relative to that of the CR-free rats. This is an indication that at 20 mg CR and above, the dye per se or its metabolites[11] that get to the testis or are generated within them caused changes in testis weight.
| Table 1: | Weight changes in rat testes (n = 3-4) |
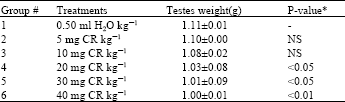 | |
| * P-value relative to group 1 | |
Decreased testis weight (Table 1) can occur as a result of impaired cell growth and/or proliferation. An indication of increased or decreased cell number can be obtained from tissue DNA level[12] . From DNA analysis (Table 2) it is evident that CR-exposure reduced the level of testicular DNA. It is therefore likely that the low testis weight is a consequence of impaired cell growth and/or proliferation. Besides nuclear DNA in the testes, sperm head DNA is another location and source of DNA in the organ. Consequently hypospermatogenesis, complete inhibition of the process or injury of sperm producing cells would cause a reduction in the level of DNA in the affected testis relative to normal testis. Results suggesting that benzidine-based azodyes alter testicular development and cause hypospermatogenesis in mice during adulthood have been described by previous workers[2]. In the on-going investigation, part of which makes up the present report, we obtained histopathological evidence (data not shown) that the testis of rats exposed to 10 mg CR and above were damaged to varying extents. Particularly basement membranes of the tubular cell (germ cells) were disrupted. This would have contributed to cell death, low testicular DNA level and hence low testis weight. These interpretations appear to be supported by the correlation analysis between testis weight and testis DNA content, which has r-value of +0.81 and a P-value of <0.05 (Table 4).
Table 2 shows that exposure of rats to CR did not alter testicular protein content significantly when compared to the control rats. Since cell proliferation involves increased DNA production, there ought to be a corresponding change in protein level. Present data suggest otherwise. However, it is pertinent to keep in mind that what we analysed is total cellular protein not nuclear or DNA associated chromatin proteins (predominantly histones) which alter with nuclear DNA simultaneously. Indeed inhibition of histone synthesis results in decreased rate of DNA synthesis; the converse is also true[13]. Hence presently we cannot discountenance CR impaired cell proliferation as the cause of loss in testis weight on the basis of total testicular protein analysis. (Table 2).
| Table 2: | DNA and Protein content of rat testes (n = 3-4) |
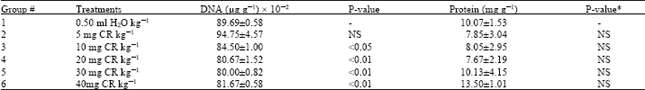 | |
| *P-value relative to group 1 | |
| Table 3: | ACP and ALP Activities in CR-exposed and CR-free rat testes |
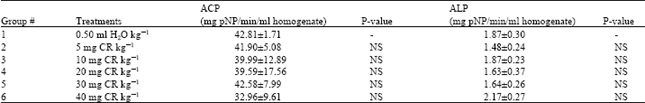 | |
| Table 4: | Correlation coefficients and P-values |
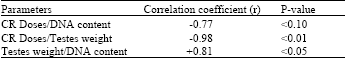 | |
Testicular ACP and ALP activities were not affected in CR-exposed rats. (Table 3). Evidently these two testicular enzymes are not sensitive to either the dye or its reduction products and/or their derivatives such as benzidine, 1,2-diaminonaphthalene-4-sulphonate, mono- and diacetyl benzidine[14,15]. In earlier studies the ability of this dye to inhibit alkaline phosphatase from rat bone, kidney and liver[16]as well as rabbit and chicken serum alkaline phosphatase and L-aspartate aminotransferase activities[17] in vitro was demonstrated. However, the intestinal isoform of alkaline phosphatase was not inhibited[16]. So the inability of the dye or its metabolites to inhibit ACP and ALP in the testis under in vivo condition may well be due to the nature of the testicular isoforms of both enzymes. Another possibility is that inhibition is effected only by the parent compound (the dye per se). But following in vivo metabolism of the dye in the GIT[11] and other tissue en route to the testes, its inhibitory ability is lost. In the study reported by Gray and Ostby[2] the benzidine based azodyes were administered to mice at a dose of 1 g/kg/day by gavage for 3 gestational days. This high dose according to the authors was predicated because of the large amount of the dye usually excreted in the faeces without metabolism and/or absorption. Although we found it to be so, it appears from the present study that even at considerably lower doses the toxic effects of benzidine based azodyes can still be evaluated by using biochemical indices. Furthermore present results indicated that sufficient toxic metabolites able to cause detectable physical and biochemical changes in the testes were generated from the low CR doses.
In conclusion, Congo red administration significantly lowers testis weight in rat probably due to (i) an impairment of testicular cell growth and/or proliferation and (ii) CR-induced testicular cell injury and death. This is attested to by significantly low DNA content in the testes of rats exposed to 10 mg CR and above.
REFERENCES
- Rinde, E. and W. Troll, 1975. Metabolic reduction of benzidine Azo-dye to benzidine in the rhesus monkey. J. Nat. Cancer Inst., 55: 181-182.
PubMed