
Mohammad A. Awal
Department of Anatomy and
Histology,
Bangladesh Agricultural
University,
Mymensingh-2202, Bangladesh
Masamichi Kurohmaru
Department of Veterinary Anatomy, Graduate School of Agricultural and Life Sciences, The University of Tokyo, Bunkyo-ku, Tokyo 1 1 3-8657, Japan
Takuo Mizukami
Department of Veterinary Anatomy, Graduate School of Agricultural and Life Sciences, The University of Tokyo, Bunkyo-ku, Tokyo 1 1 3-8657, Japan
Bibin B. Andriana
Department of Veterinary Anatomy, Graduate School of Agricultural and Life Sciences, The University of Tokyo, Bunkyo-ku, Tokyo 1 1 3-8657, Japan
Yoshiakira Kanai
Department of Veterinary Anatomy, Graduate School of Agricultural and Life Sciences, The University of Tokyo, Bunkyo-ku, Tokyo 1 1 3-8657, Japan
Yoshihiro Hayashi
Department of Veterinary Anatomy, Graduate School of Agricultural and Life Sciences, The University of Tokyo, Bunkyo-ku, Tokyo 1 1 3-8657, Japan
Journal of Medical Sciences
Year: 2002 | Volume: 2 | Issue: 1 | Page No.: 19-23







ABSTRACT
The investigation was conducted to evaluate the effect of bisphenol-A on the sertoli cell culture from prepubertal Wistar rats. Sertoli cell culture was obtained by sequential collagenase and trypsin-EDTA treatment. The cell culture was incubated and maintained for 21 days. The culture was trypsinized and seeded at an appropriate density. Three days later, bisphenol-A was added to the cell culture at the concentration of 10, 100 and 1000pg ml-1, respectively. Control group was treated with saline. The cultured cells were examined by light and transmission electron microscopy. A phase contrast microscope was also used to observe the morphological changes of the cells. At 2 days after treatment, the increase of degenerative sertoli cell was 7.2, 14.7, and 20.8% at the concentration of 10, 100, and 1000pg ml-1, respectively. Similarly, at 4 days, those increased as 11.4, 19.5, 28.3%, and at 6 days as 12.6, 21.3 and 34.9%, respectively. The maximum degenerative change of sertoli cell was observed at the concentration of 1000pg ml-1 and at 6 days after treatment. They were significantly different from those in the control group (p<0.05). Distinct histopathological changes in the sertoli cell culture were noted after exposure to bisphenol-A. This findings suggested that bisphenol-A showed a direct effect on sertoli cells in culture.
PDF Abstract XML References Citation
How to cite this article
Mohammad A. Awal, Masamichi Kurohmaru, Takuo Mizukami, Bibin B. Andriana, Yoshiakira Kanai and Yoshihiro Hayashi, 2002. Effect of Bisphenol-A on the Sertoli Cell Culture from Prepubertal Male Wistar Rats. Journal of Medical Sciences, 2: 19-23.
DOI: 10.3923/jms.2002.19.23
URL: https://scialert.net/abstract/?doi=jms.2002.19.23
DOI: 10.3923/jms.2002.19.23
URL: https://scialert.net/abstract/?doi=jms.2002.19.23
INTRODUCTION
A large number of cell lines obtained from mammalian tissues have been used to evaluate the effect of biochemical and environmental hormones (endocrine disruptors) on organism (Perez et al., 1998). Endocrine disruptors are a diverse group of chemicals that alter the functions of endocrine system. Several endocrine disruptors are known to have an estrogenic activity in nearly all classes of vertebrates (Kloas et al., 1999; Krishnan et al., 1993). Plastic and plastic products, widely used for daily necessaries, contain estrogenic endocrine-disruptors. Kembra et al. (1999), reported that these products can interfere with the mammalian reproduction. At present, it has been accepted that the reproductive organs are the major targets for their actions (Monsees et al., 2000; Silvana et al., 2000).
The findings on polychlorinated biphenyl and pentachlorophenol have revealed that these drugs cause the decrease of testicular weight and sperm production, and alteration of testicular morphology (Beard et al., 1999, Carlsen et al., 1992). Bisphenol-A has generally been used as polycarbonate plastic products such as baby bottles and packaging of beverage and food (Feldman and Krishnan, 1995). Takao et al. (1999), reported that bisphenol-A leaches out from such products, and its rate increases with repeated use. It has been suggested that environmental contaminants with bisphenol-A affect the reproduction and other endocrine systems in human and wild life (Toppari et al., 1996). To date, only a few studies on the effect of bisphenol-A have been carried out (Silvana et al., 2000; Akita et al., 2000). Most of them have been conducted in vivo (Kembra et al., 1999; Beard et al., 1999; Colborn et al., 1993).
Therefore, this research work was designed to evaluate the effect of bisphenol-A on the sertoli cell culture from prepubertal Wistar rats.
MATERIALS AND METHODS
Digestion procedure: Ten days old NRC Wistar male rats (body weight 20-21g) were purchased from the Japan Biological Research Center. The sertoli cell suspension suitable for culture was obtained according to the method of Mather et al. (1982) Under pentobarbital anesthesia; decapsulated testes were enzymatically digested with sequential collagen digestion. Digestive solution was composed of Dulbecco’s Minimum Essential Medium (DMEM) + equal volume of 0.1% collagenase, 100Fl of 0.1% hyaluronidase and 0.01% DNase (Sigma Chemical Co., Ltd., USA).
The dissociated seminiferous tubules were pipetted gently and allowed for shaking water bath at 37°C for 30min (120 oscillations min-1). They were centrifuged and washed with DMEM. This treatment was repeated twice. To obtain single cells, trypsin-EDTA solution (Gibco chemical Co., Ltd.,USA) was added to the cell suspension. They were centrifuged and the sediment was washed with bovine serum albumin (BSA)-trypsin inhibitor solution (Sigma chemical Co., Ltd., USA). The cells obtained were washed with DMEM, and treated with antibiotics (200 unit penicillin 100 IU ml-1, streptomycin 100, gentamicin 40, fungizone 0.5Fg ml-1, respectively) supplemented with 10% fetal bovine serum (Cansera International Inc., Canada). The isolated sertoli cells were cultured in a plastic dish for initial culture using the procedure of Part and wood (1996). The cells were seeded at a density of 106 cells cm-2 (determined using a hemocytometer) into 100 cm2 culture dish (Fulcon®), and kept at 32.5°C in a humidified atmosphere containing 95% air and 5% CO2 for 3 weeks. Non–adherent cells were removed by changing the medium at 24hr. and again at 96hr. Until this time, the medium contained antibiotics. Thereafter, the medium was changed every 3 days until the cells were harvested for final culture. The culture was maintained for 3 weeks to avoid germ cells contamination.
For harvesting, the medium was removed, and the petridish was rinsed with phosphate buffer saline (PBS) followed by addition of trypsin-EDTA solution. Then, the culture was incubated at 37°C for about 10 min. The dish was vigorously pipetted and agitated mechanically. The cell suspension was then treated with BSA-trypsin inhibitor solution and filtered with 70Fm nylon filter (Falcon®). The cells were washed several times with DMEM, pelleted by centrifugation, and then resuspended in antibiotic-free medium (Chris and Peter, 1997). The cell density was determined (5 x 105 cells cm-2) before plating the cells on the 24 Multi well plate (Sumilon®), coating coverslips (Sumitomo Beck Right Chemical Co., Ltd., Tokyo, Japan), and/or the Millipore filter with reconstituted basement membrane extracts. Millipore ® (Hadley et al., 1988).
Preparation of Millipore filter chamber: The permeable filters used for Millipore cell culture inserts were 0.45Fm in pore size and 12mm in diameter (Millipore® Cyclopore polyethylene terephthalate; Becton Dickinson, Franklin Lakes, New Jersey, USA). The cells grew on the upper surface of the permeable filter. The culture medium was added to both the insert (upper surface) and the well (lower surface). Prior to seeding, each membrane was wetted for 2-3hr with antibiotic-free medium. The cells were placed in the center of the insert membrane and shook by gentle swirling. At 24hr after seeding and then every 48hr., the medium was changed.
Preparation for seeding on coated cover slips and Multi well plate: Collagen coated cover slips (Serodex 13.5 ΨTYPE l, Japan) were placed on the bottom of 24 Multi well plates. The cells suspension was added to the center of cover slips.
Three days later after final seeding of cells, the sertoli cell culture was treated with bisphenol-A (Wako Pure Chemical Industries Ltd., Japan) at the concentration of 10, 100 and 1000pg ml-1, respectively. The control group was treated with saline only. At 2, 4 and 6 days after treatment, the sertoli cells plated on coating cover slips and Millipore chambers were processed for microscopic study. The cells plated on the Multi well plates were routinely observed with a phase contrast microscope (Olympus, Japan). The results were statistically analyzed with Student’s t test.
Light microscopy:
Coated cover slips: The medium was removed and the cover slips were gently rinsed 2-3 times with PBS followed by fixation with 4% paraformaldehyde for 30 min. The specimens were allowed to dry overnight at room temperature and then stained with hematoxylin for 3hr. followed by counterstaining with eosin for 10 min. They were mounted on glass slides, dried, and observed by light microscopy.
Millipore inserts: The medium was removed, the Millicell was gently washed with PBS, fixed with 3% glutaraldehyde in PBS (pH, 7.3) for 15 min and followed by 0.5% Triton x 100 in Milli-Q water for 5 min. The Millicell was stained with hematoxylin, placed on Durapore membrane (Durapore®; 12mm diamater), mounted on glass slides, and observed by light microscopy.
Transmission electron microscopy: For transmission electron microscopy, the specimens were fixed with 2.5% glutaraldehyde-0.05M cacodylate buffer (pH, 7.4) at 4°C for 1hr. and then
washed 3 or 4 times with the same buffer. They were postfixed with 1% Osmium tetraoxide (OsO)4 for 1hr. dehydrated through a graded series of ethanol (C2H5OH), and embedded in Araldite. Thin sections of 1Fm thickness were cut and stained with 1% toluidine blue for light microscopy. Ultrathin sections were cut and stained with uranyl acetate and lead citrate and examined with a JEM-1200 EX transmission electron microscope at 80 kV.
RESULTS
Light microscopy: Within 48hr of incubation, sertoli cells formed an epithelial-like monolayer on the plastic support and impregnated filter membrane. Contaminated germ cells were totally removed after 21 days of culture (Fig. 1a, c and e). At 2 days after treatment, the increase of degenerative sertoli cells were 7.2, 14.7, and 20.8% at the concentration of 10, 100, and 1000pg ml-1, respectively. Similarly, at 4 days, those increased as 11.4, 19.5, 28.3% and at 6 days as 12.6, 21.3 and 34.9%, respectively (Fig. 3). The maximum degenerative change of sertoli cells were observed at the concentration of 1000pg ml-1 and at 6 days after treatment (Fig. 1b, d and f). Thus, the dose and time-dependent increases in the number of degenerating sertoli cells were observed in all the treated groups. They were significantly different from those in the control group, (p<0.05).
By phase contrast microscopy, the similar degenerative changes in the sertoli cells were also recognized (Fig. 1e-f).
Transmission electron microscopy: The sertoli cells on the Millipore filter membrane were examined by transmission electron microscopy. Most of the cells in the control group revealed normal in structure. While, in treated groups, degenerating sertoli cells which possessed several large vacuoles, electron-dense materials and so on were frequently recognized (Fig. 2a, b).
DISCUSSION
Although a number of toxicological investigation have been carried out using with the sertoli cell culture (Alexander et al., 1996; Robins et al., 1997) most of them focussed on the secretory functions of sertoli cell. On the other hand, a number of in vivo and in vitro studies have been made to clarify the mechanism of the action of bisphenol-A and other endocrine-disruptors on testicular tissues. Beard et al. (1999), reported that pentachlorophenol altered testicular growth and sperm maturation in ram by inhibiting the secretion of thyroid hormone. Silvana et al. (2000) revealed that the polychlorinated biphenyl compound brought about reproductive toxicity in rats by inhibiting testicular androgenesis.
Until now, however, there is only one report regarding the effects of bisphenol-A on the sertoli cell culture from the immature rats (Monsees et al., 2000). They reported that the secretion of lactate and inhibit from the cells increased significantly by exposure of the sertoli cell culture to bisphenol-A.
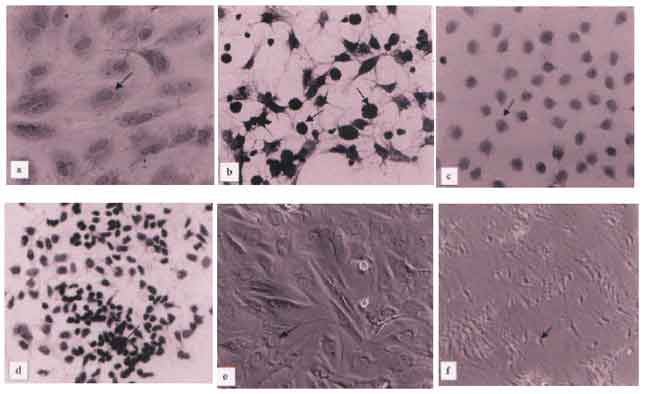 | |
| Fig. 1: | Light micrographs of cultured sertoli cells in control (a,c,e) and treated (b,d,f) groups. a) sertoli cells grow on coated cover slips. Arrow indicates the nucleus of sertoli cell. b) Degenerated sertoli cells (arrows) are observed on cover slips. c) Sertoli cells grow on Millipore filter chamber. Arrow indicates the nucleus of sertoli cell. d) Degenerated sertoli cells (arrows) are observed on Millipore filter chamber. e) Phase contrast microscopic figure of cultured sertoli cells on Multi well plate. f) Arrow indicates the degenerated sertoli cell on Multi well plate (x20 each) |
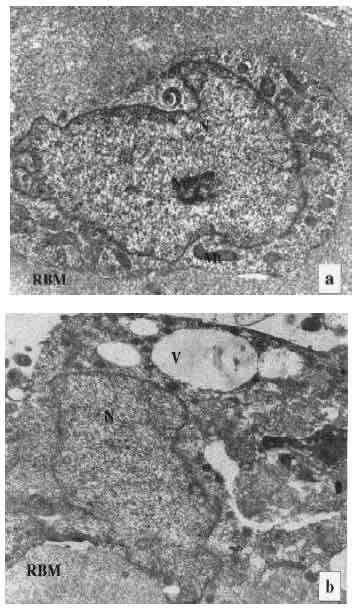 | |
| Fig. 2: | Transmission electron micrographs of sertoli cells in culture. a) The sertoli cell shows normal in structure (control). N-nucleus, Nu-nucleolus, Mt-mitochondria. RBM-reconstituted basement membrane. b) The sertoli cell has some vacuoles and electron dense materials in culture treated with bisphenol-A for 6 days. N-nucleus. V-vacuoles. Asterisk (*)-electron dense material (x6000) |
Different from the experiment on secretory aspects of sertoli cells, this investigation dealed with the degeneration rate of sertoli cells in culture by their exposure to bisphenol-A. Our electron microscopic observation confirmed that the sertoli cells in culture showed a degenerative change by the treatment of bisphenol-A. In this study, the dose and time-dependent increase in the number of degenerating sertoli cells were found out in all bisphenol-A treated groups. The number of degenerated sertoli cells in the treated groups were significantly different from those in the control group (p<0.05). The maximum degenerative change was recognized at the concentration of 1000pg ml-1 and at 6 days after treatment. From these observation, it has become obvious that bisphenol-A may showed a direct action on sertoli cells in culture.
Various toxicants as well as endocrine disruptors including bisphenol-A are known to affect sertoli cells (Monsees et al., 2000). The exposure to these reproductive toxicants at prepubertal life reduced the sertoli cell replication (Sharpe et al., 1995). In most species, including man, sertoli cells proliferate at fetal, neonatal and prepubertal stages. This proliferation is mainly controlled by FSH, and ceases at the onset of puberty (Pelliniemi et al., 1993). The exposure to exogenous estrogens at these stages of life can lead to suppression of FSH secretion by the pituitary gland and cause a reduced rate of sertoli cells count. The significantly decreased number of sertoli cells in this study is in well agreement with the report of Sharpe et al. (1995).
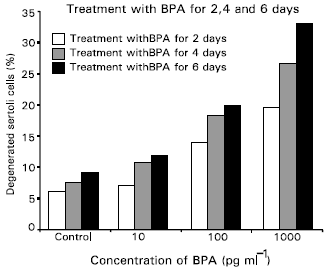 | |
| Fig. 3: | The number of degenerated sertoli cells in culture in control and bisphenol-A treated groups. Dose-and time-dependent increases in the number of degenerated sertoli cells in culture treated with bisphenol-A are obviously recognized. Treated groups are significantly different control groups (p<0.05). Maximum degenerative changes of sertoli cells are observed at the concentration of 1000pgml-1. |
Janecki et al. (1992) investigated the effects of cadmium chloride (CdCl2) on sertoli cell functions by using two-compartment cultures from immature rats. They reported that the toxic effect of this compound depended on the concentration as well as on the duration of exposure. Our findings are well consistent with this report.
In order to elucidate the mechanism of bisphenol-A induced testicular dysfunction, a number of investigation have been carried out in vivo and in vitro. Several hypotheses have been advocated on the possible mechanism of action of the reproductive toxicants. However, it is still remains to be unsolved (Krishnan et al., 1993; Kembra et al., 1999).
In conclusion, it is suggested here that treatment with relatively high-dose of bisphenol-A showed a direct effect on sertoli cells in culture from prepubertal rats. A further investigation should be needed to clarify the mechanism of the action of this compound to sertoli cells.
ACKNOWLEDGMENTS
The authors thank Mr. I. Tsugiyama (Department of Veterinary Anatomy, The University of Tokyo) for his technical assistance during conducting this research. This work was supported in part by a grant-in-aid from Japan Society for Promotion of Science (JSPS), and in part from the Ministry of Education, Science, Sports and Culture, Japan.
REFERENCES
- Alexander, B.H., H. Checkoway, C. van Netten, C.H. Muller and T.G. Ewers et al., 1996. Semen quality of men employed at a lead smelter. Occup. Environ. Med., 53: 411-416.
Direct Link - Yokoyama, A., M. Akita, S. Shimizu, Y. Nozaki, H. Shikuma and Y. Kuroda, 2000. Effects of bisphenol A on cultured rat embryos. J. Toxicol. Sci., 25: 243-243.
Direct Link - Mather, J.P., I.Z. Zhuang, V. Perez-Infante and D.M. Phillips, 1982. Culture of testicular cells in hormone-supplemented serum-free medium. Ann. N. Y. Acad. Sci., 383: 44-68.
PubMedDirect Link - Part, P. and C.M. Wood, 1996. Na/H exchange in cultured epithelial cells from fish gills. J. Comp. Physiol. B, 166: 37-45.
CrossRefDirect Link - Perez, P., R. Pulgar, F. Olea-Serrano, M. Villalobos and A. Rivas et al., 1998. The estrogenicity of bisphenol A-related diphenylalkanes with various substituents at the central carbon and the hydroxy groups. Environ. Health Perspect., 106: 167-174.
CrossRefPubMedDirect Link - Robins, T.G., M.S. Bornman, R.I. Ehrlich, A.C. Cantrell, E. Pienaar, J. Vallabh and S. Miller, 1997. Semen quality and fertility of men employed in a South African lead acid battery plant. Am. J. Ind. Med., 32: 369-376.
PubMedDirect Link - Sharpe, R.M., J.S. Fisher, M.M. Millar, S. Jobling and J.P. Sumpter, 1995. Gestational and lactational exposure of rats to xenoestrogens results in reduced testicular size and sperm production. Environ. Health Perspect., 103: 1136-1143.
CrossRefPubMedDirect Link - Andric, S.A., T.S. Kostic, S.S. Stojilkovic and R.Z. Kovacevic, 2000. Inhibition of rat testicular androgenesis by a polychlorinated biphenyl mixture aroclor 1248. Biol. Reprod., 62: 1882-1888.
PubMedDirect Link - Toppari, J., J.C. Larsen, P. Christiansen, A. Giwercman and P. Grandjean et al., 1996. Male reproductive health and environmental xenoestrogens. Environ. Health Perspect., 104: 741-803.
Direct Link - Takao, Y., H.C. Lee, Y. Ishibashi, S. Kohra, N. Tominaga and K. Arizono, 1999. Fast screening method for bisphenol-A in environmental water and food by solid-phase microextraction (SPME). J. Health Sci., 45: 39-39.
CrossRefDirect Link - Monsees, T.K., M. Franz, S. Gebhardt, U. Winterstein, W.B. Schill and J. Hayatpour, 2000. Sertoli cells as a target for reproductive hazards. Andrologia, 32: 239-246.
CrossRefDirect Link - Krishnan, A.V., P. Stathis, S.F. Permuth, L. Tokes and D. Feldman, 1993. Bisphenol-A: An estrogenic substance is released from polycarbonate flasks during autoclaving. Endocrinology, 132: 2279-2286.
PubMedDirect Link - Kloas, W., I. Lutz and R. Einspanier, 1999. Amphibians as a model to study endocrine disruptors: II. Estrogenic activity of environmental chemicals in vitro and in vivo. Sci. Total Environ., 225: 59-68.
CrossRefDirect Link - Howdeshell, K.L., A.K. Hotchkiss, K.A. Thayer, J.G. Vandenbergh and F.S. vom Saal, 1999. Environmental toxins: Exposure to bisphenol A advances puberty. Nature, 401: 763-764.
CrossRefDirect Link - Beard, A.P., P.M. Bartlewski, R.K. Chandolia, A. Honaramooz and N.C. Rawlings, 1999. Reproductive and endocrine function in rams exposed to the organochlorine pesticides lindane and pentachlorophenol from conception. J. Reprod. Fert., 115: 303-314.
PubMedDirect Link - Colborn, T., F.S. vom Saal and A.M. Soto, 1993. Developmental effects of endocrine-disrupting chemicals in wildlife and humans. Environ. Health Perspect., 101: 378-384.
Direct Link - Carlsen, E., A. Giwercman, N. Keiding and N.E. Skakkebaek, 1992. Evidence for decreasing quality of semen during past 50 years. Br. Med. J., 305: 609-613.
PubMedDirect Link - Wood, C.M. and P. Part, 1997. Cultured branchial epithelia from freshwater fish gills. J. Exp. Biol., 200: 1047-1059.
PubMedDirect Link - Feldman, D. and A. Krishnan, 1995. Estrogens in unexpected places: Possible implications for researchers and consumers. Environ. Health Perspect., 103: 129-133.
CrossRefDirect Link - Hadley, M.A., S.W. Byers, C.A. Suarez-Quian, D. Djakiew and M. Dym, 1988. In vitro models of differentiated sertoli cell structure and function. In Vitro Cell. Dev. Biol., 24: 550-557.
CrossRefPubMedDirect Link - Janecki, A., A. Jakubowiak and A. Steinberger, 1992. Effect of cadmium chloride on transepithelial electrical resistance of sertoli cell monolayers in two-compartment cultures-A new model for toxicological investigations of the blood-testis barrier in vitro. Toxicol. Applied Pharmacol., 112: 51-57.
CrossRefDirect Link